
Genome-Wide Identification of Vascular Plant One-Zinc-Finger Gene Family in Six Cucurbitaceae Species and the Role of CmoVOZ2 in Salt and Drought Stress Tolerance

Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of VOZ Proteins in Cucurbitaceae and Evolutionary Analysis
2.2. Chromosomal Localization Analysis and Physicochemical Properties
2.3. Conserved Motifs, Gene Structure, and Cis-Acting Elements Analysis
2.4. Plant Materials and Abiotic Stress Treatment
2.5. RNA Extraction and Quantitative Real-Time PCR
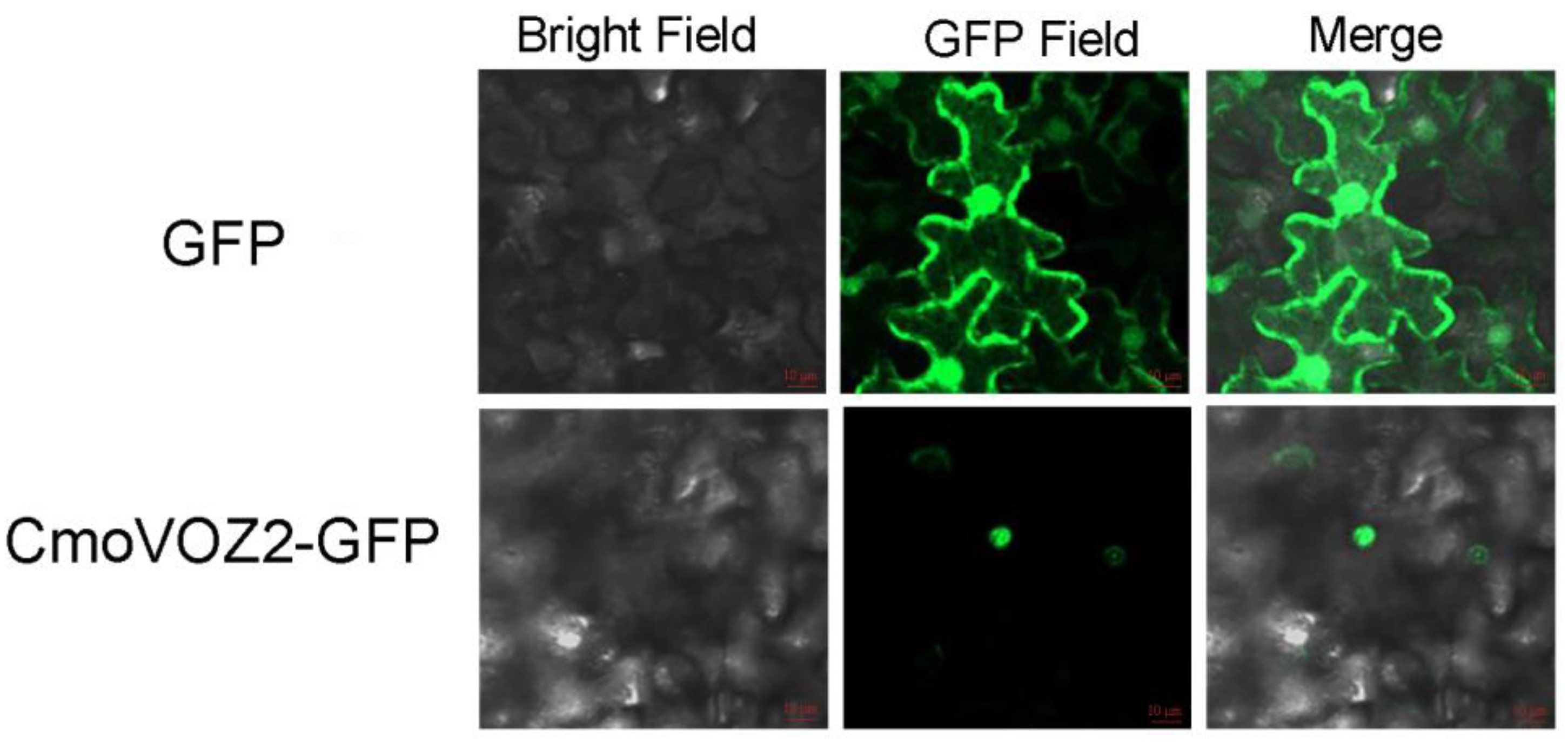
2.6. Transcriptional Activation Analysis and Subcellular Localization
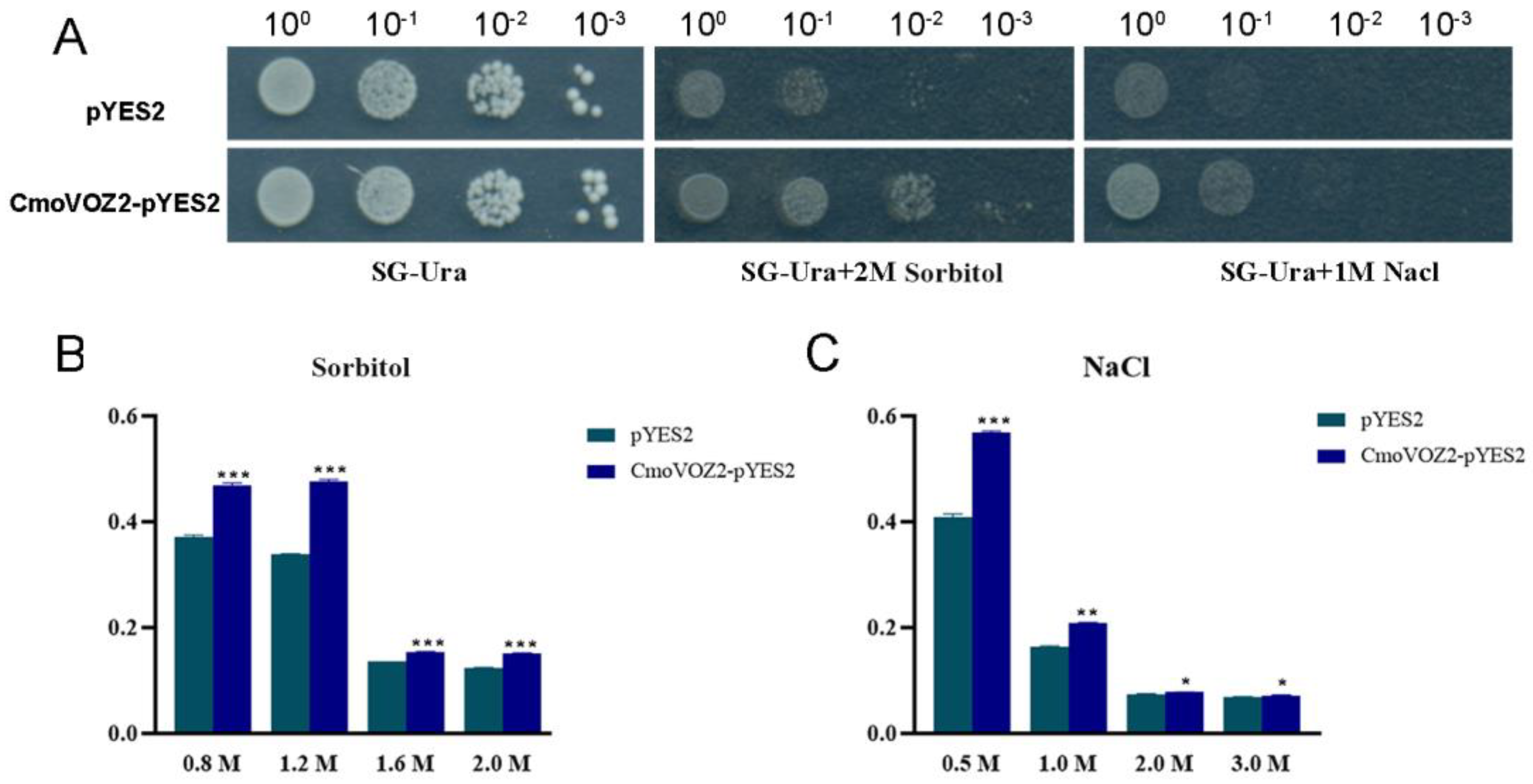
2.7. Drought and Salt Stress Tolerance Analysis of ComVOZ2 in Yeast
3. Results
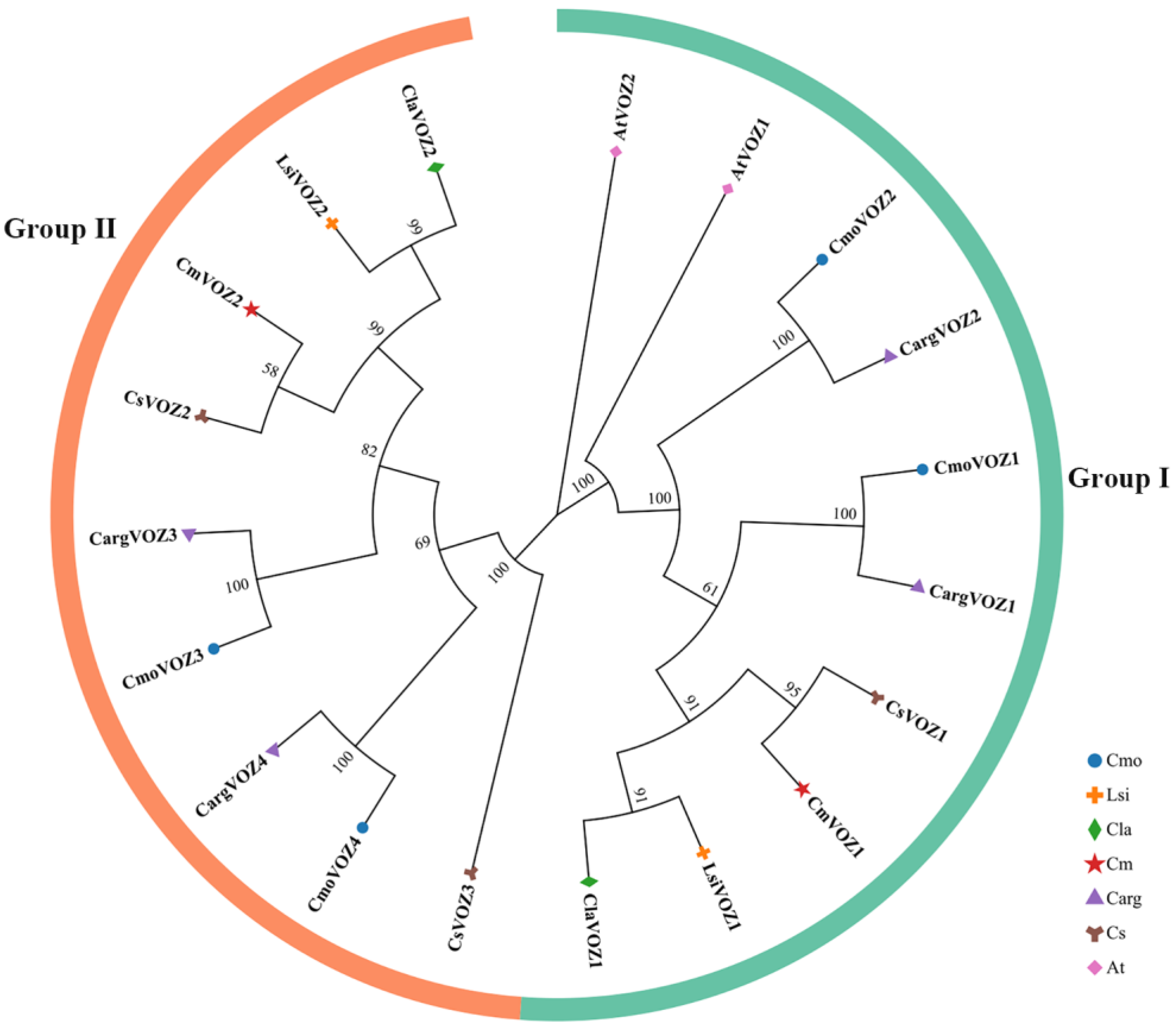
3.1. Identification and Evolutionary Analysis of VOZ Gene Family Members in Cucurbitaceae
3.2. Chromosomal Localization of VOZ Gene Members in Cucurbitaceae
3.3. Analysis of Structures and Conserved Motifs
3.4. Cis-Acting Elements Analysis
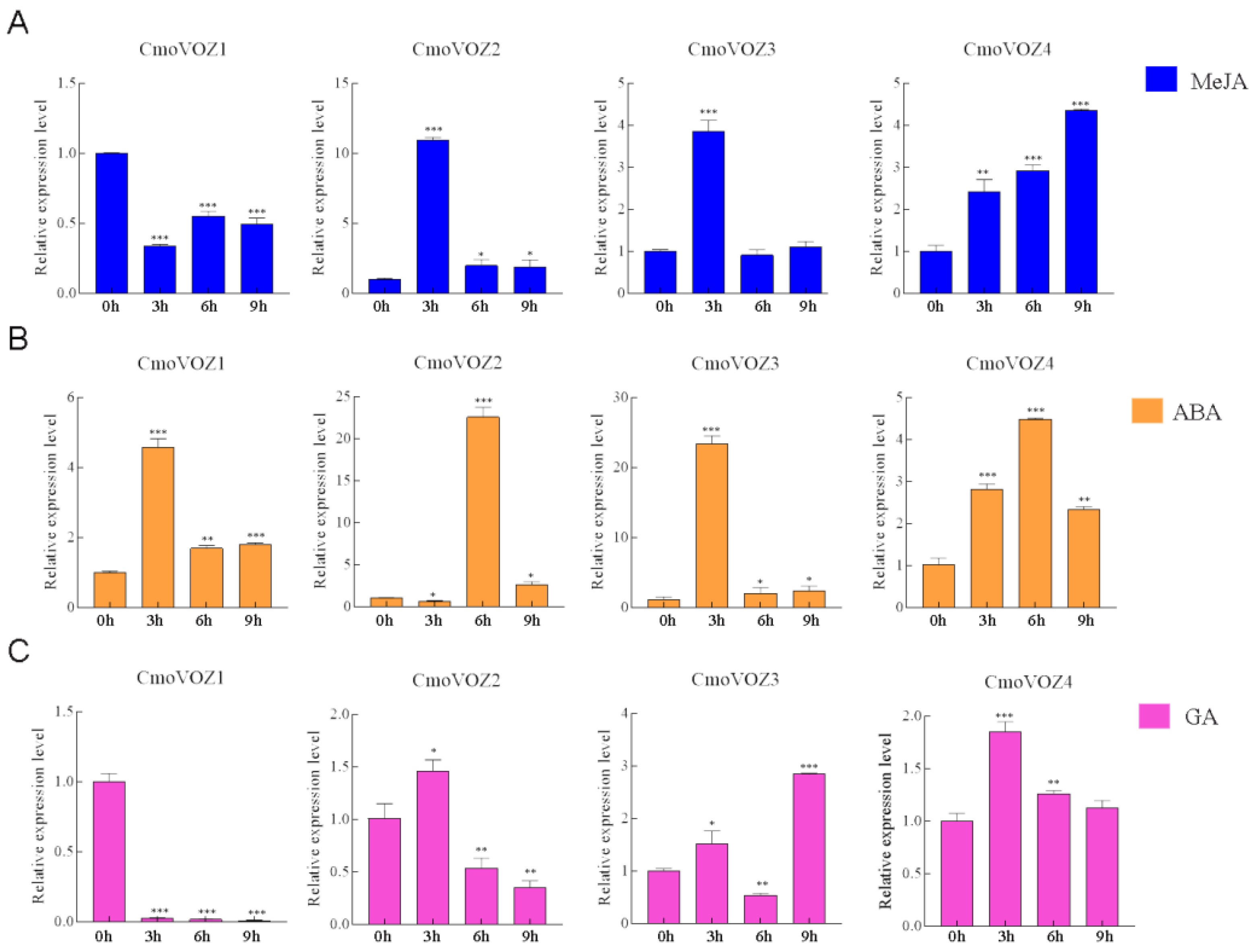
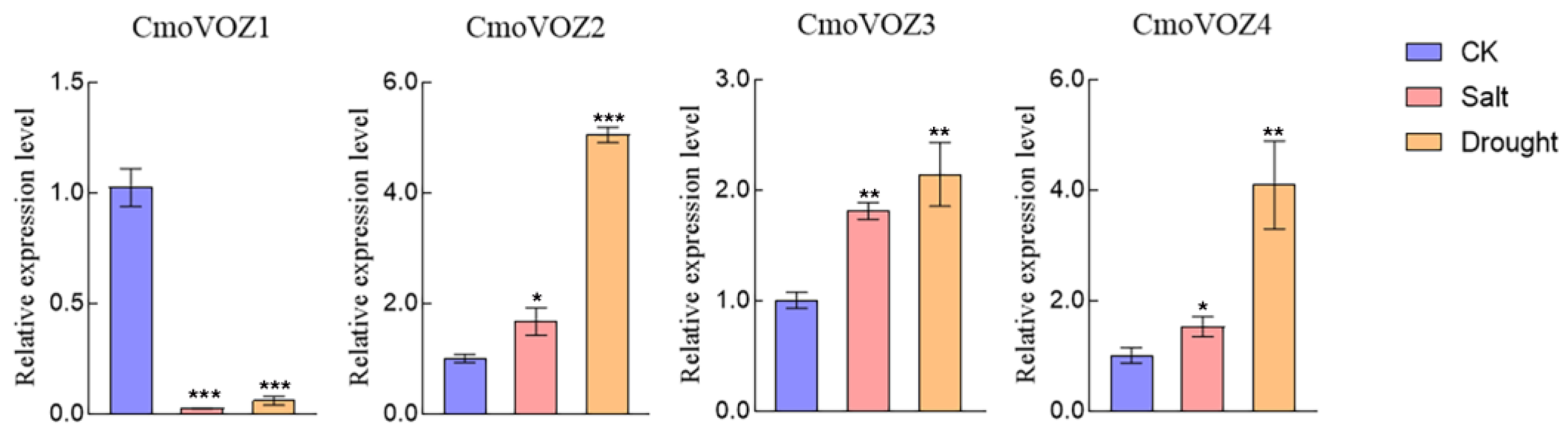
3.5. Expression Analysis of CmoVOZ Gene Family in C. moschata
3.6. Transactivation Assay and Subcellular Localization of ComVOZ2
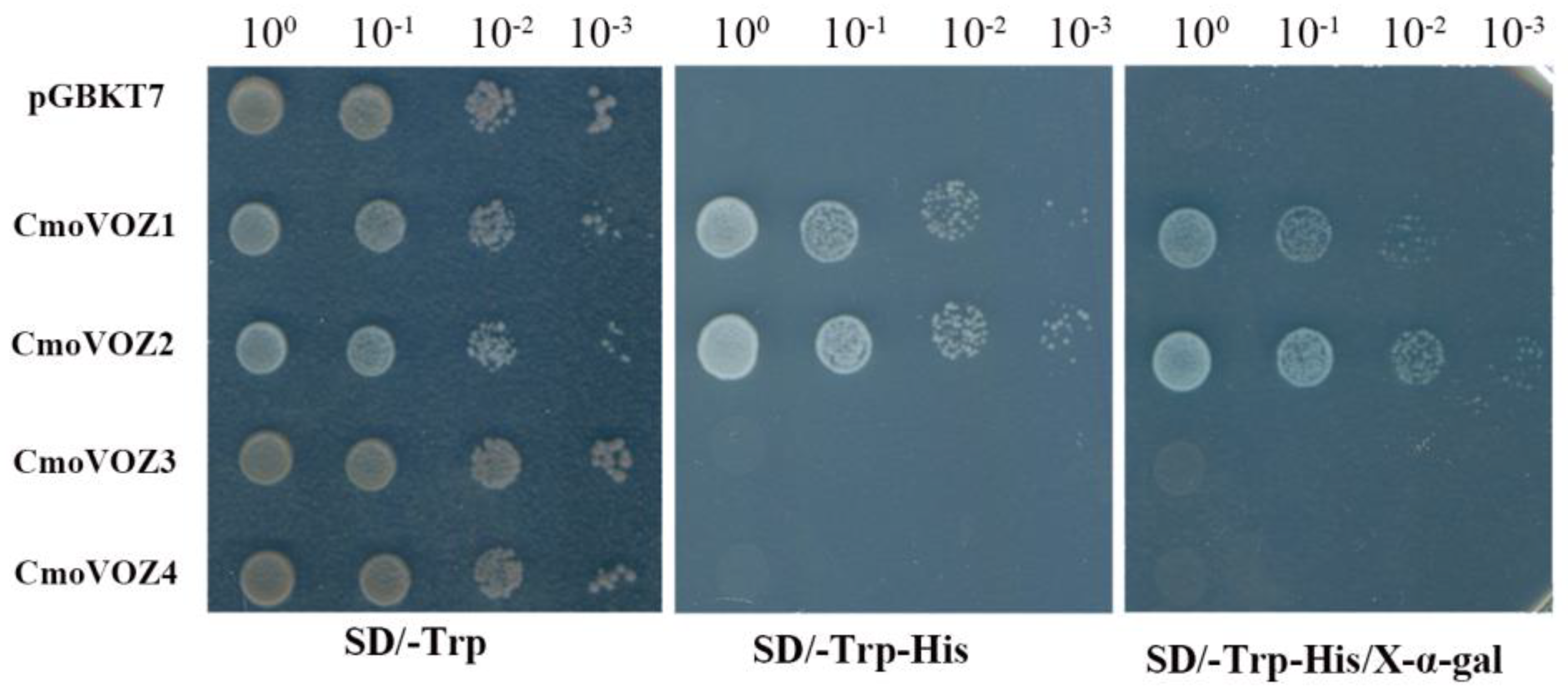
3.7. Analysis of Salt and Drought Stress Tolerance of CmoVOZ2 Gene in Yeast
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Nykiel, M.; Gietler, M.; Fidler, J.; Prabucka, B.; Labudda, M. Abiotic stress signaling and responses in plants. Plants 2023, 12, 3405. [Google Scholar] [CrossRef]
- Sarwar, R.; Geng, R.; Li, L.; Shan, Y.; Zhu, K.M.; Wang, J.; Tan, X.L. Genome-wide prediction, functional divergence, and characterization of stress-responsive BZR transcription factors in B. napus. Front. Plant Sci. 2021, 12, 790655. [Google Scholar] [CrossRef] [PubMed]
- Gupta, B.; Huang, B. Mechanism of salinity tolerance in plants: Physiological, biochemical, and molecular characterization. Int. J. Genom. 2014, 2014, 701596. [Google Scholar] [CrossRef] [PubMed]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional regulatory network of plant heat stress response. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Wang, H.; Shao, H.; Tang, X. Recent advances in utilizing transcription factors to improve plant abiotic stress tolerance by transgenic technology. Front. Plant Sci. 2016, 7, 67. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.A.; Li, M.Z.; Wang, S.M.; Yin, H.J. Revisiting the role of plant transcription factors in the battle against abiotic stress. Int. J. Mol. Sci. 2018, 19, 1634. [Google Scholar] [CrossRef] [PubMed]
- Baillo, E.H.; Kimotho, R.N.; Zhang, Z.; Xu, P. Transcription factors associated with abiotic and biotic stress tolerance and their potential for crops improvement. Genes 2019, 10, 771. [Google Scholar] [CrossRef]
- Jin, J.; Tian, F.; Yang, D.C.; Meng, Y.Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Mitsuda, N.; Hisabori, T.; Takeyasu, K.; Sato, M.H. VOZ; isolation and characterization of novel vascular plant transcription factors with a one-zinc finger from Arabidopsis thaliana. Plant Cell Physiol. 2004, 45, 845–854. [Google Scholar] [CrossRef]
- Celesnik, H.; Ali, G.S.; Robison, F.M.; Reddy, A.S. Arabidopsis thaliana VOZ (Vascular plant one-zinc finger) transcription factors are required for proper regulation of flowering time. Biol. Open 2013, 2, 424–431. [Google Scholar] [CrossRef]
- Li, B.; Zheng, J.C.; Wang, T.T.; Min, D.H.; Wei, W.L.; Chen, J.; Zhou, Y.B.; Chen, M.; Xu, Z.S.; Ma, Y.Z. Expression analyses of soybean VOZ transcription factors and the role of GmVOZ1G in drought and salt stress tolerance. Int. J. Mol. Sci. 2020, 21, 2177. [Google Scholar] [CrossRef]
- Shi, P.; Jiang, R.; Li, B.; Wang, D.; Fang, D.; Yin, M.; Yin, M.; Gu, M. Genome-wide analysis and expression profiles of the VOZ gene family in Quinoa (Chenopodium quinoa). Genes 2022, 13, 1695. [Google Scholar] [CrossRef]
- Uluisik, S.; Kiyak, A.; Kurt, F.; Filiz, E. Genome-wide identification of the VOZ transcription factors in tomato (Solanum lycopersicum): Their functions during fruit ripening and their responses to salinity stress. J. Hortic. Sci. Biotechnol. 2023, 98, 468–482. [Google Scholar] [CrossRef]
- Prasad, K.; Xing, D.; Reddy, A.S.N. Vascular plant one-zinc-finger (VOZ) transcription factors are positive regulators of salt tolerance in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 3731. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Chairattanawat, C.; Vo, K.T.X.; Liu, J.; Zhang, J.; Pan, T.; Kim, D.-Y.; Martinoia, E.; Zhong, C.-Y.; Wang, M.-H.; et al. VOZ1 and VOZ2 transcription factors regulate arsenic tolerance and distribution in rice and Arabidopsis. Front. Plant Sci. 2023, 14, 1209860. [Google Scholar] [CrossRef] [PubMed]
- Nakai, Y.; Nakahira, Y.; Sumida, H.; Takebayashi, K.; Nagasawa, Y.; Yamasaki, K.; Akiyama, M.; Ohme-Takagi, M.; Fujiwara, S.; Shiina, T.; et al. Vascular plant one-zinc-finger protein 1/2 transcription factors regulate abiotic and biotic stress responses in Arabidopsis. Plant J. 2013, 73, 761–775. [Google Scholar] [CrossRef]
- Selote, D.; Matthiadis, A.; Gillikin, J.W.; Sato, M.H.; Long, T.A. The E3 ligase BRUTUS facilitates degradation of VOZ1/2 transcription factors. Plant Cell Environ. 2018, 41, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Koguchi, M.; Yamasaki, K.; Hirano, T.; Sato, M.H. Vascular plant one-zinc-finger protein 2 is localized both to the nucleus and stress granules under heat stress in Arabidopsis. Plant Signal Behav. 2017, 12, e1295907. [Google Scholar] [CrossRef]
- Nakai, Y.; Fujiwara, S.; Kubo, Y.; Sato, M.H. Overexpression of VOZ2 confers biotic stress tolerance but decreases abiotic stress resistance in Arabidopsis. Plant Signal Behav. 2013, 8, e23358. [Google Scholar] [CrossRef]
- Kumar, S.; Choudhary, P.; Gupta, M.; Nath, U. Vascular plant one-zine finger1 (VOZ1) and VOZ2 Interact with constans and promote photoperiodic flowering transition. Plant Physiol. 2018, 176, 2917–2930. [Google Scholar] [CrossRef]
- Schaefer, H.; Renner, S.S. Phylogenetic relationships in the order Cucurbitales and a new classification of the gourd family (Cucurbitaceae). Taxon 2011, 60, 122–138. [Google Scholar] [CrossRef]
- Huang, S.; Li, R.; Zhang, Z.; Li, L.; Gu, X.; Fan, W.; Lucas, W.J.; Wang, X.; Xie, B.; Ni, P.; et al. The genome of the cucumber, Cucumis sativus L. Nat. Genet. 2009, 41, 1275–1281. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, J.; Sun, H.; Salse, J.; Lucas, W.J.; Zhang, H.; Zheng, Y.; Mao, L.; Ren, Y.; Wang, Z.; et al. The draft genome of watermelon (Citrullus lanatus) and resequencing of 20 diverse accessions. Nat. Genet. 2013, 45, 51–58. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, Y.; McGregor, C.; Liu, S.; Luan, F.; Gao, M.; Weng, Y. Genetic architecture of fruit size and shape variation in cucurbits: A comparative perspective. TAG. Theoretical and applied genetics. Theor. Appl. Genet. 2020, 133, 1–21. [Google Scholar] [CrossRef]
- Perkins-Veazie, P.; Collins, J.K.; Davis, A.R.; Roberts, W. Carotenoid content of 50 watermelon cultivars. J. Agric. Food Chem. 2006, 54, 2593–2597. [Google Scholar] [CrossRef]
- Han, X.N.; Liu, C.Y.; Liu, Y.L.; Xu, Q.M.; Li, X.R.; Yang, S.L. New triterpenoids and other constituents from the fruits of Benincasa hispida (Thunb.) Cogn. J. Agric. Food Chem. 2013, 61, 12692–12699. [Google Scholar] [CrossRef] [PubMed]
- Thoennissen, N.H.; Iwanski, G.B.; Doan, N.B.; Okamoto, R.; Lin, P.; Abbassi, S.; Song, J.H.; Yin, D.; Toh, M.; Xie, W.D.; et al. Cucurbitacin B induces apoptosis by inhibition of the JAK/STAT pathway and potentiates antiproliferative effects of gemcitabine on pancreatic cancer cells. Cancer Res. 2009, 69, 5876–5884. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Bao, J.; Guo, J.; Ding, Q.; Lu, J.; Huang, M.; Wang, Y. Biological activities and potential molecular targets of cucurbitacins: A focus on cancer. Anti-Cancer Drugs 2012, 23, 777–787. [Google Scholar] [CrossRef] [PubMed]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary gnetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, The Swiss Bioinformatics Resource Portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Xu, M.; Wang, Y.; Zhang, M.; Chen, M.; Ni, Y.; Xu, X.; Xu, S.; Li, Y.; Zhang, X. Genome-wide identification of BES1 gene family in six Cucurbitaceae species and its expression analysis in Cucurbita moschata. Int. J. Mol. Sci. 2023, 24, 2287. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Zhu, W.; Jiao, D.; Zhang, J.; Xue, C.; Chen, M.; Yang, Q. Genome-wide identification and analysis of BES1/BZR1 transcription factor family in potato (Solanum tuberosum L.). Plant Growth Regul. 2020, 92, 375–387. [Google Scholar] [CrossRef]
- Zhang, W.; Yang, G.; Zou, X.; Mu, D.; Li, H.; Zang, D.; Wang, Y. Expression of ethylene response factors (ERFs) From Betula platyphylla and the confer salt and drought tolerance analysis in a yeast. J. Plant Biochem. Biotechnol. 2017, 26, 35–42. [Google Scholar] [CrossRef]
- Gao, B.; Chen, M.; Li, X.; Liang, Y.; Zhu, F.; Liu, T.; Zhang, D.; Wood, A.J.; Oliver, M.J.; Zhang, J. Evolution by duplication: Paleopolyploidy events in plants reconstructed by deciphering the evolutionary history of VOZ transcription factors. BMC Plant Biol. 2018, 18, 256. [Google Scholar] [CrossRef]
- Long, M.; Rosenberg, C.; Gilbert, W. Intron phase correlations and the evolution of the intron/exon structure of genes. Proc. Natl. Acad. Sci. USA 1995, 92, 12495–12499. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Guo, C.; Shan, H.; Kong, H. Divergence of duplicate genes in exon-intron structure. Proc. Natl. Acad. Sci. USA 2012, 109, 1187–1192. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Taxon (Species) | Ploidy | N a | N b | Gene Name | Gene ID | CDS (bp) | aa | Mw (kDa) | pI | Ref. |
|---|---|---|---|---|---|---|---|---|---|---|
| C. argyrosperma | diploid | 10 | 4 | CargVOZ1 | Carg07755 | 1452 | 483 | 54.03 | 5.88 | |
| CargVOZ2 | Carg15320 | 1482 | 493 | 55.20 | 6.22 | |||||
| CargVOZ3 | Carg03109 | 1464 | 487 | 54.47 | 5.67 | |||||
| CargVOZ4 | Carg05122 | 1287 | 420 | 47.01 | 5.17 | |||||
| C. lanatus | diploid | 11 | 2 | ClaVOZ1 | Cla97C01G023230.1 | 1452 | 483 | 53.95 | 5.87 | |
| ClaVOZ2 | Cla97C02G036280.1 | 1422 | 473 | 52.94 | 5.83 | |||||
| C. melo | diploid | 12 | 2 | CmVOZ1 | MELO3C007248.1 | 1452 | 483 | 54.06 | 6.04 | |
| CmVOZ2 | MELO3C019321.1 | 1404 | 440 | 49.23 | 6.44 | |||||
| C. moschata | allotetraploid | 20 | 4 | CmoVOZ1 | CmoCh07G002120.1 | 1452 | 483 | 54.03 | 5.88 | |
| CmoVOZ2 | CmoCh03G012970.1 | 1452 | 576 | 64.76 | 6.42 | |||||
| CmoVOZ3 | CmoCh20G006520.1 | 1464 | 487 | 54.49 | 5.64 | |||||
| CmoVOZ4 | CmoCh02G001990.1 | 1287 | 428 | 47.83 | 5.18 | |||||
| C. sativus | diploid | 7 | 3 | CsVOZ1 | CsaV3_6G050500.1 | 1452 | 483 | 53.98 | 6.04 | |
| CsVOZ2 | CsaV3_2G034550.1 | 1398 | 465 | 52.20 | 5.69 | |||||
| CsVOZ3 | CsaV3_4G028820.1 | 252 | 83 | 9.52 | 9.69 | |||||
| L. siceraria | diploid | 11 | 2 | LsiVOZ1 | Lsi01G002760.1 | 1482 | 493 | 55.07 | 5.88 | |
| LsiVOZ2 | Lsi10G000870.1 | 1593 | 530 | 59.30 | 5.76 | |||||
| A. thaliana | diploid | 5 | 2 | AtVOZ1 | At1g28520 | 1461 | 486 | 54.08 | 5.73 | [9,16,17] |
| AtVOZ2 | At2g42400 | 1353 | 450 | 50.57 | 5.17 | |||||
| Oryza sativa | diploid | 12 | 2 | OsVOZ1 | LOC_Os01g54930 | 1290 | 429 | 48.11 | 5.10 | [15] |
| OsVOZ2 | LOC_Os05g43950 | 1926 | 641 | 69.90 | 5.39 | |||||
| Glycine max | tetraploid | 9 | 6 | GmVOZ1A | Glyma.07G202700 | 1434 | 477 | 53.12 | 5.87 | [11] |
| GmVOZ1C | Glyma.13G172900 | 1437 | 478 | 53.35 | 5.71 | |||||
| GmVOZ1E | Glyma.11G211900 | 1380 | 459 | 51.16 | 5.83 | |||||
| GmVOZ1G | Glyma.10G068900 | 1398 | 465 | 51.63 | 6.21 | |||||
| GmVOZ2B | Glyma.06G285800 | 1437 | 478 | 54.02 | 5.56 | |||||
| GmVOZ2D | Glyma.12G120100 | 1440 | 479 | 53.85 | 5.26 | |||||
| C. quinoa | allotetraploid | 9 | 4 | CqVOZ1 | AUR62014889 | 1455 | 484 | 54.31 | 5.70 | [12] |
| CqVOZ2 | AUR62024758 | 1569 | 522 | 58.08 | 5.48 | |||||
| CqVOZ3 | AUR62031095 | 1230 | 409 | 45.93 | 7.56 | |||||
| CqVOZ4 | AUR62037692 | 1566 | 521 | 57.93 | 5.49 | |||||
| S. lycopersicum | diploid | 12 | 2 | SIVOZ1 | Solyc02g077450 | 1404 | 467 | 52.08 | 5.26 | [13] |
| SIVOZ2 | Solyc10g008880 | 1434 | 477 | 53.54 | 6.00 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Zhang, Z.; Jiao, Y.; Tu, Y.; Zhang, X. Genome-Wide Identification of Vascular Plant One-Zinc-Finger Gene Family in Six Cucurbitaceae Species and the Role of CmoVOZ2 in Salt and Drought Stress Tolerance. Genes 2024, 15, 307. https://doi.org/10.3390/genes15030307
Xu M, Zhang Z, Jiao Y, Tu Y, Zhang X. Genome-Wide Identification of Vascular Plant One-Zinc-Finger Gene Family in Six Cucurbitaceae Species and the Role of CmoVOZ2 in Salt and Drought Stress Tolerance. Genes. 2024; 15(3):307. https://doi.org/10.3390/genes15030307
Chicago/Turabian StyleXu, Minyan, Zhi Zhang, Yuhuan Jiao, Yaling Tu, and Xin Zhang. 2024. "Genome-Wide Identification of Vascular Plant One-Zinc-Finger Gene Family in Six Cucurbitaceae Species and the Role of CmoVOZ2 in Salt and Drought Stress Tolerance" Genes 15, no. 3: 307. https://doi.org/10.3390/genes15030307
APA StyleXu, M., Zhang, Z., Jiao, Y., Tu, Y., & Zhang, X. (2024). Genome-Wide Identification of Vascular Plant One-Zinc-Finger Gene Family in Six Cucurbitaceae Species and the Role of CmoVOZ2 in Salt and Drought Stress Tolerance. Genes, 15(3), 307. https://doi.org/10.3390/genes15030307