
Genome-Wide Identification and Transcriptome Analysis of the Hsp70 Gene Family in Monodonta labio Reveals Its Role in Response to Nanoplastics Stress
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of the Hsp70 Gene Family
2.2. Phylogenetic Analysis of the Hsp70 Genes
2.3. Physicochemical Properties, Gene Structure Analysis, Motif Analysis, and Chromosome Localization of Hsp70 Genes
2.4. Analyzing the Subcellular Localization and Predicting Signal Peptides of Hsp70 Proteins
2.5. Expression and Analysis of Hsp70 Genes
3. Results
3.1. Genome-Wide Identification and Sequence Analysis of Hsp70 Genes in M. labio
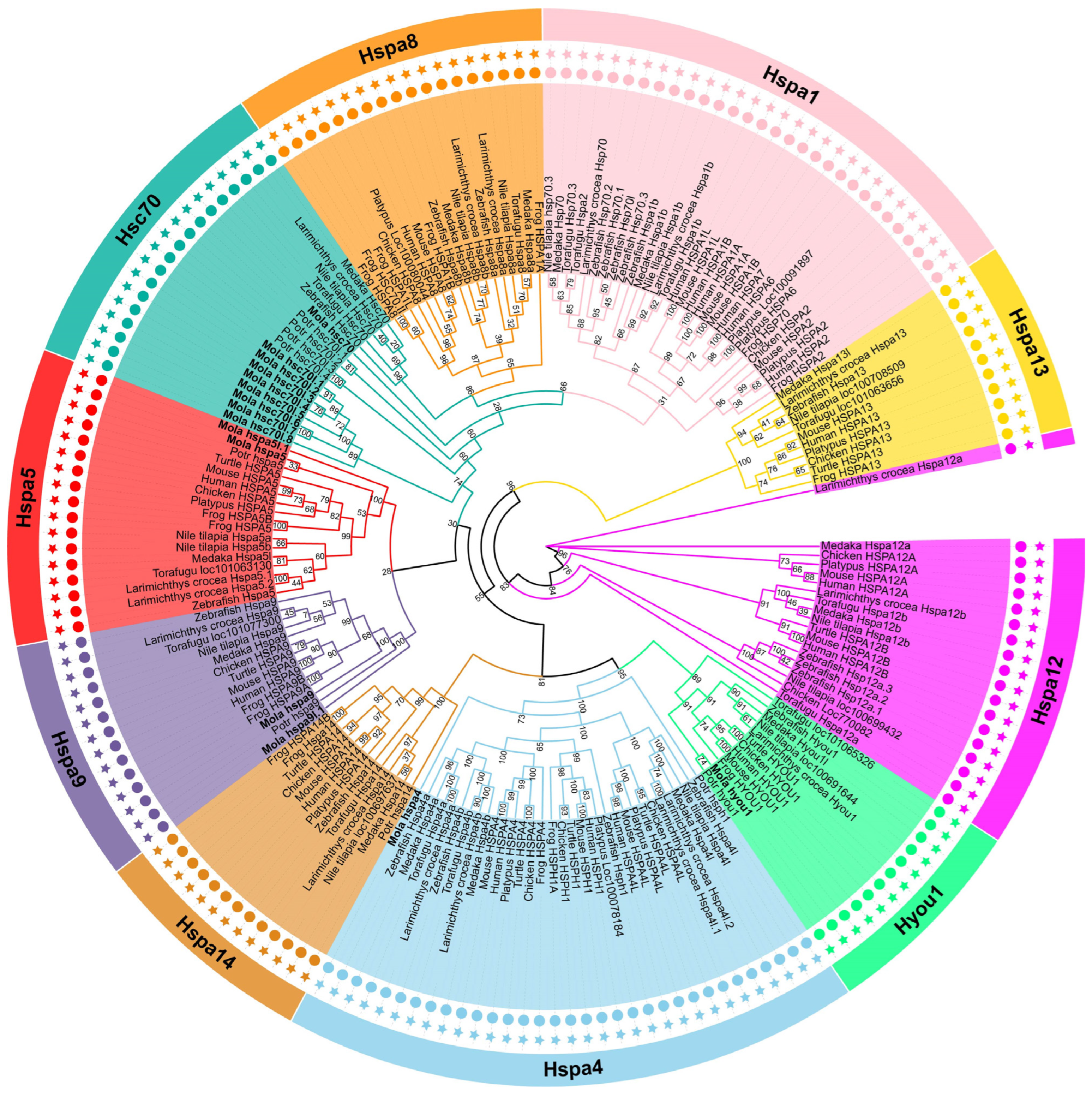
3.2. Phylogenetic Analysis of Hsp70 Genes
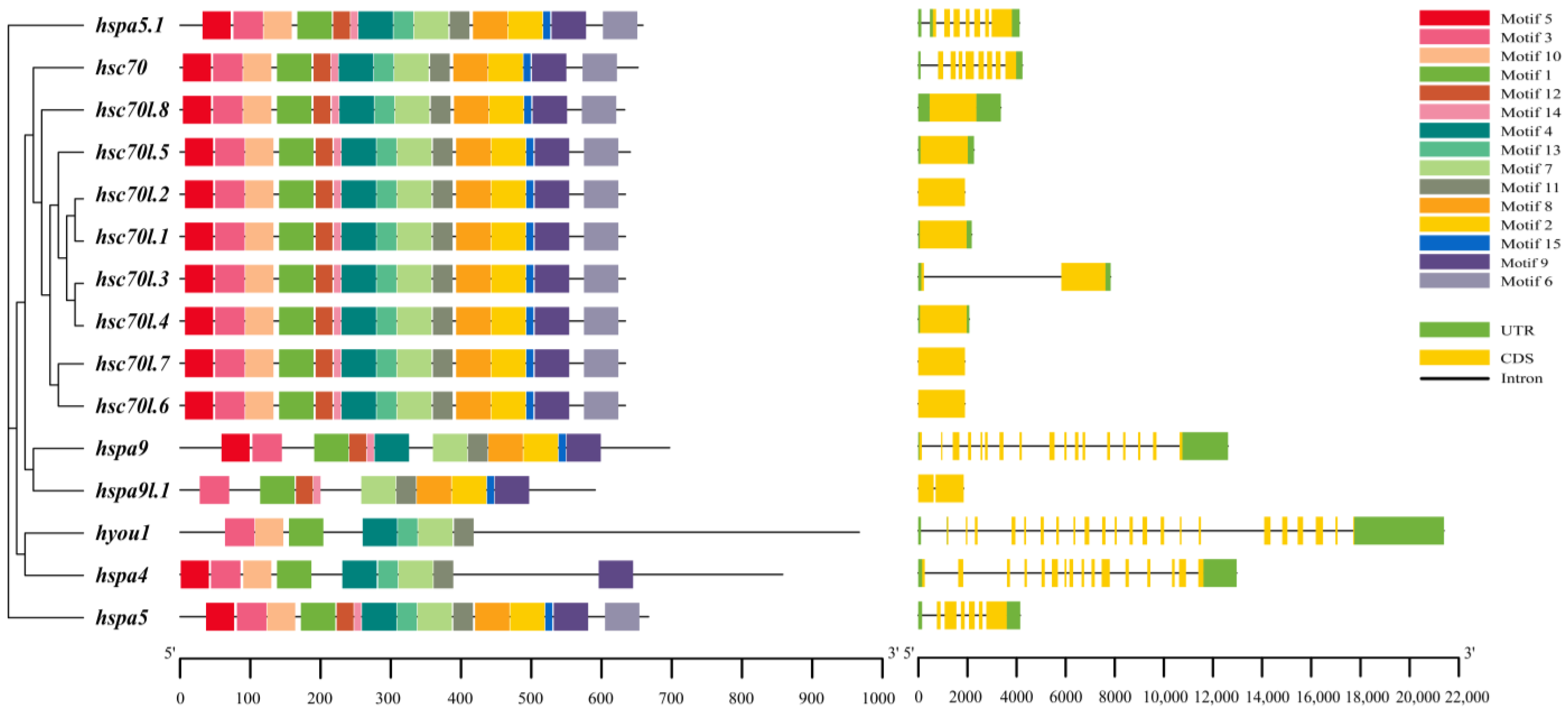
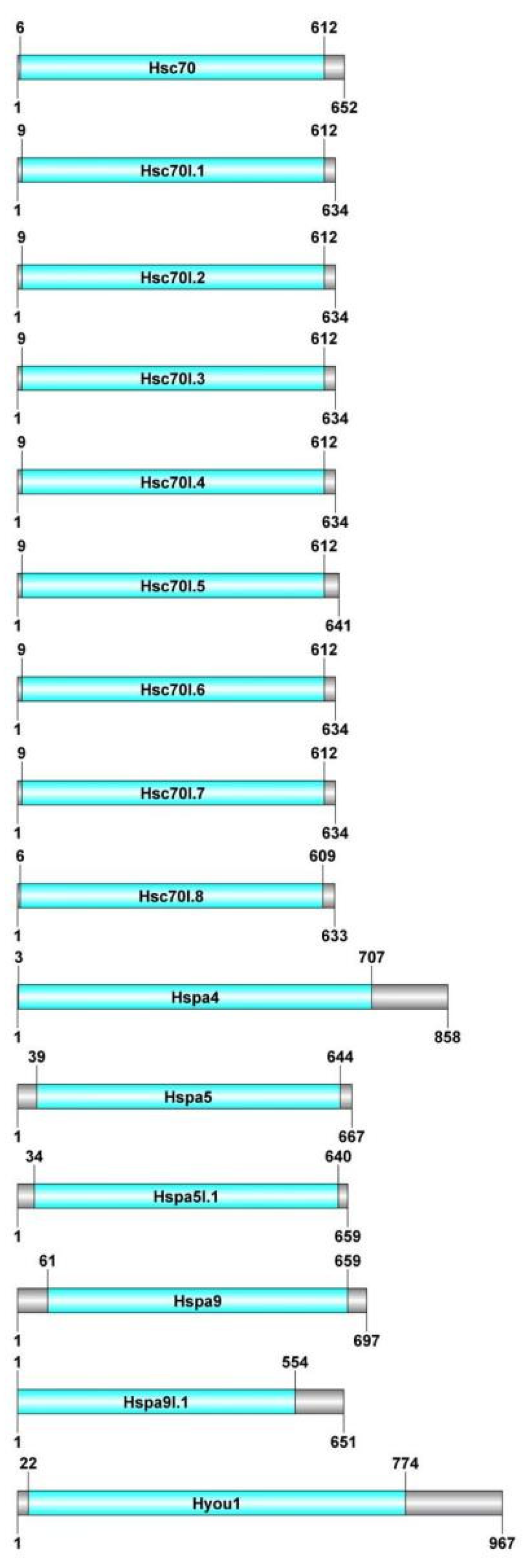
3.3. Analysis of Motif, Structure, and Conserved Domain
3.4. Secondary Structure Prediction, Subcellular Localization, and Analysis of Hsp70 Proteins
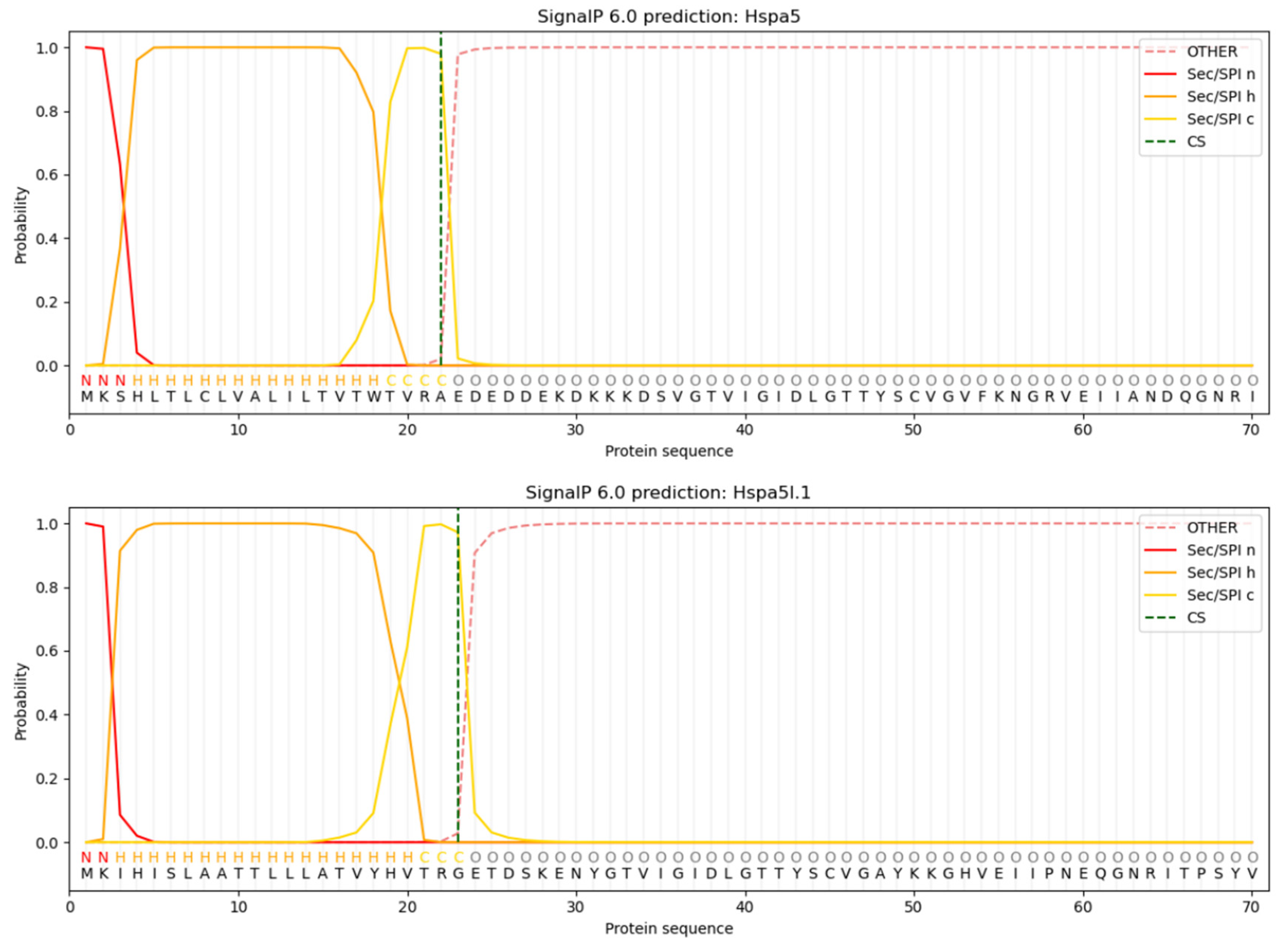
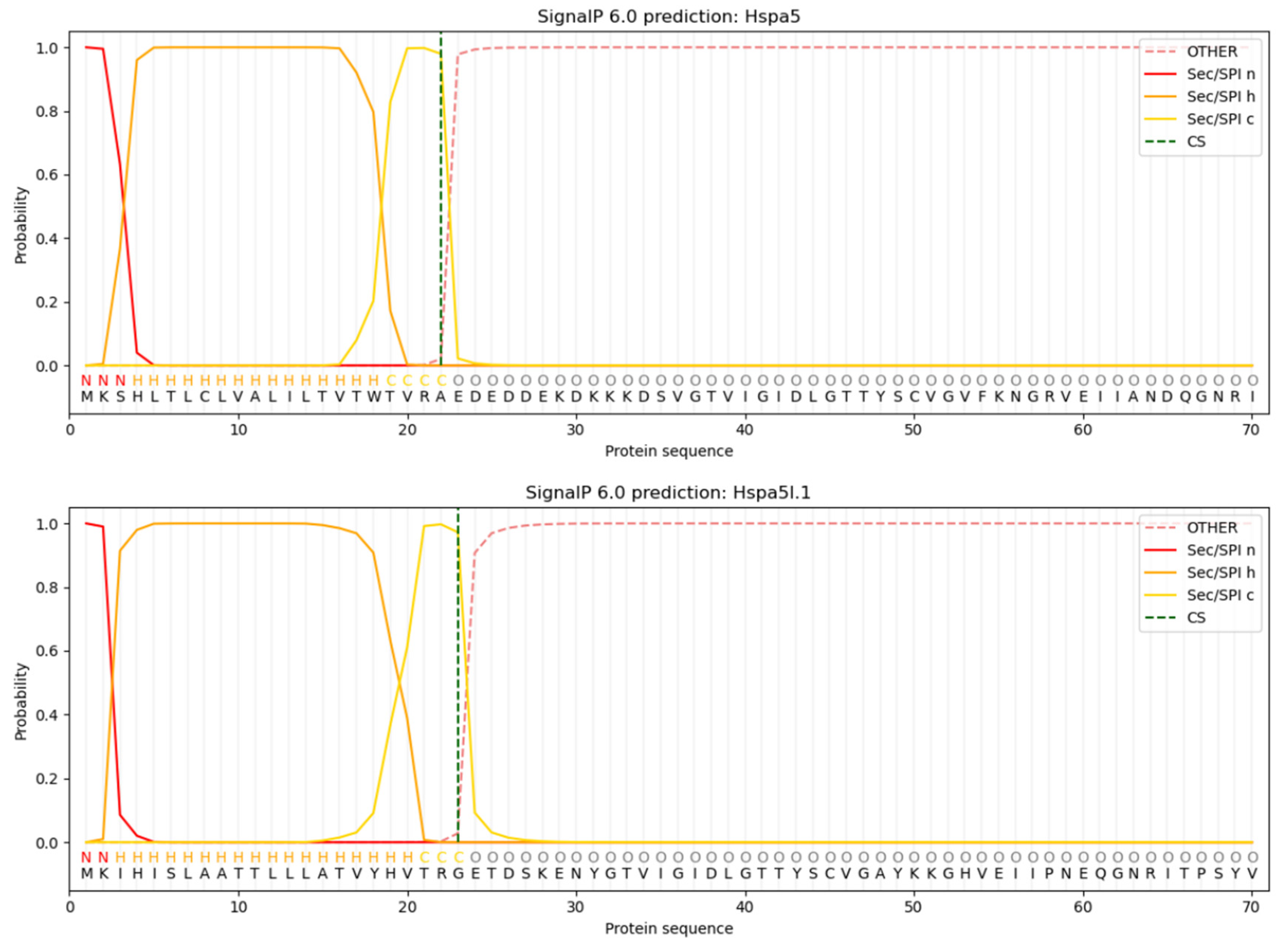
3.5. Analysis of Protein Signal Peptides Predictively
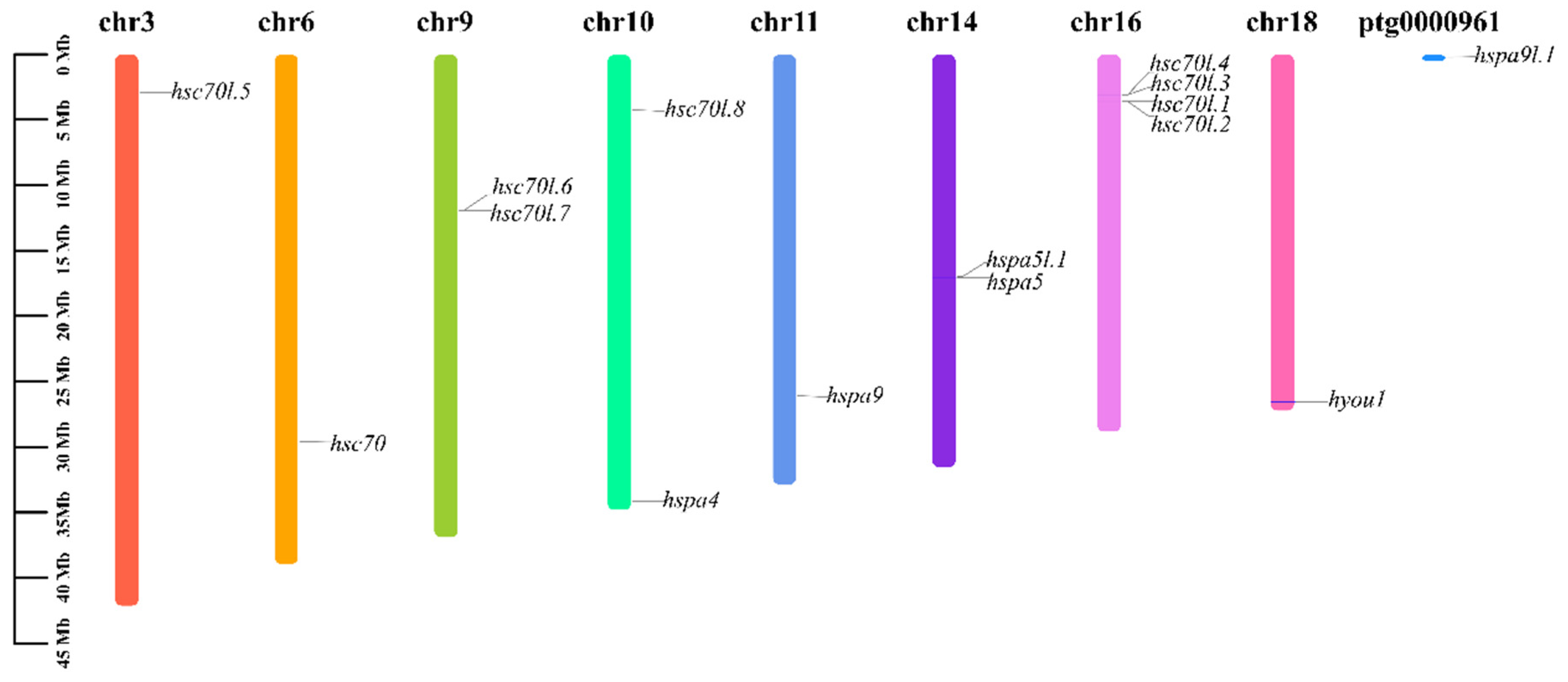
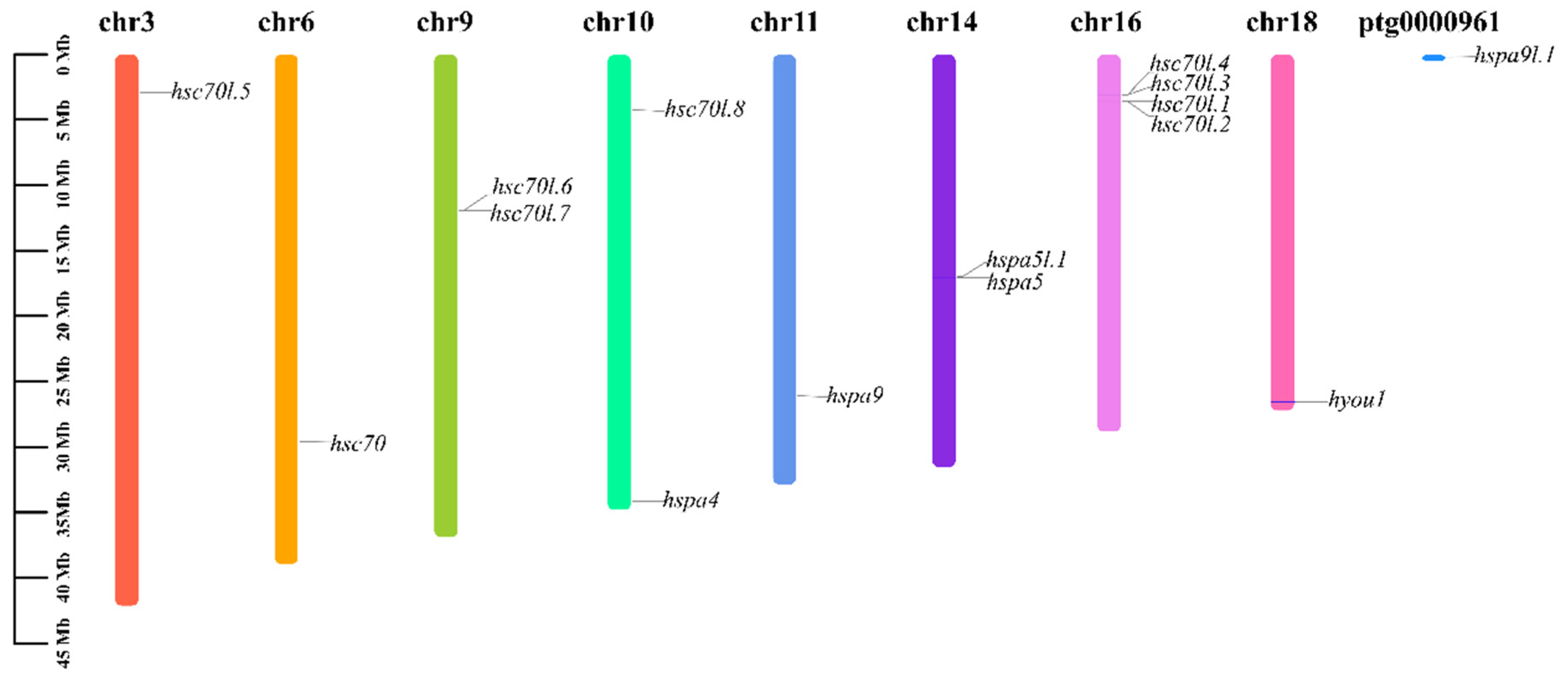
3.6. Chromosomal Location Analysis of Hsp70s
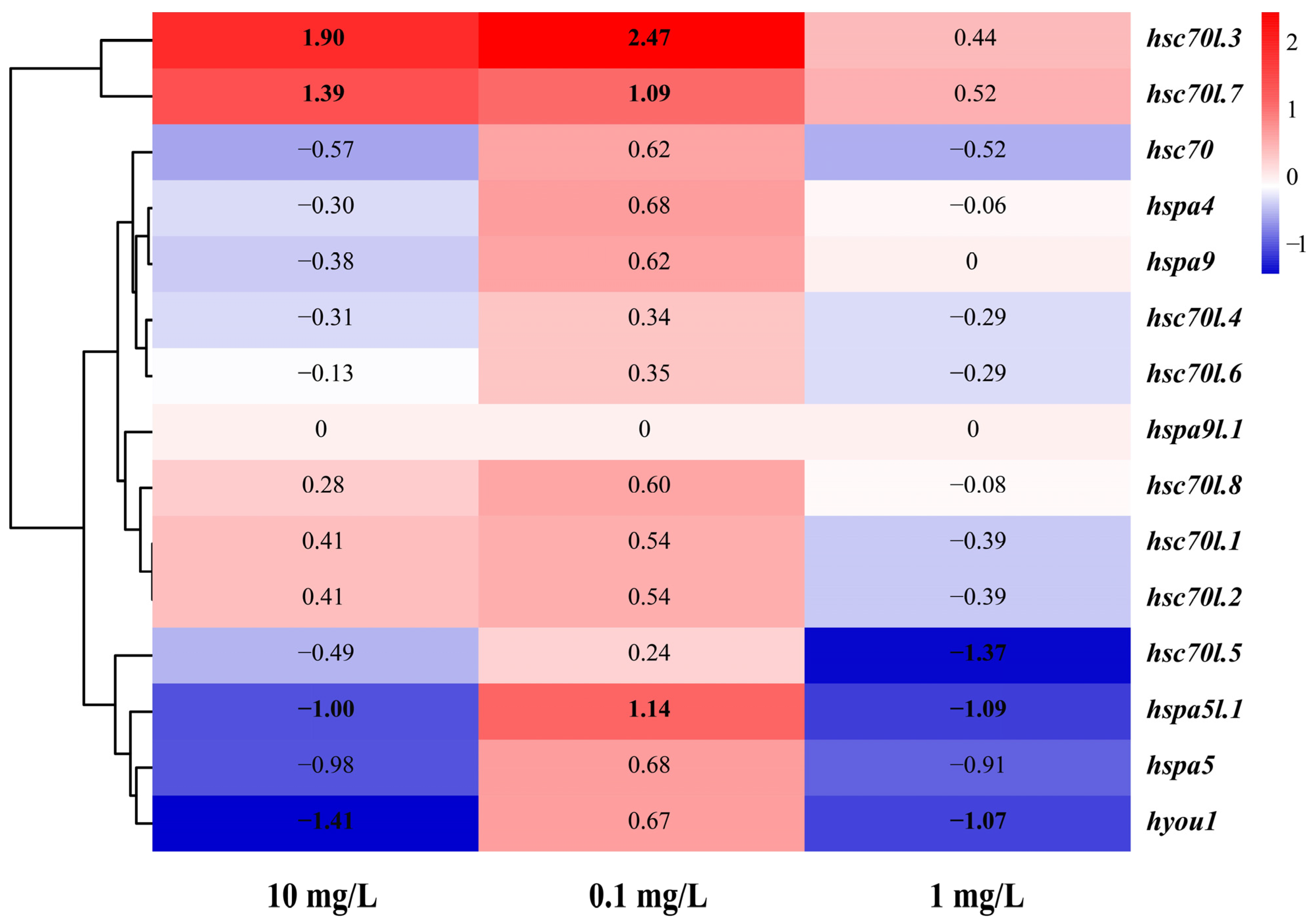
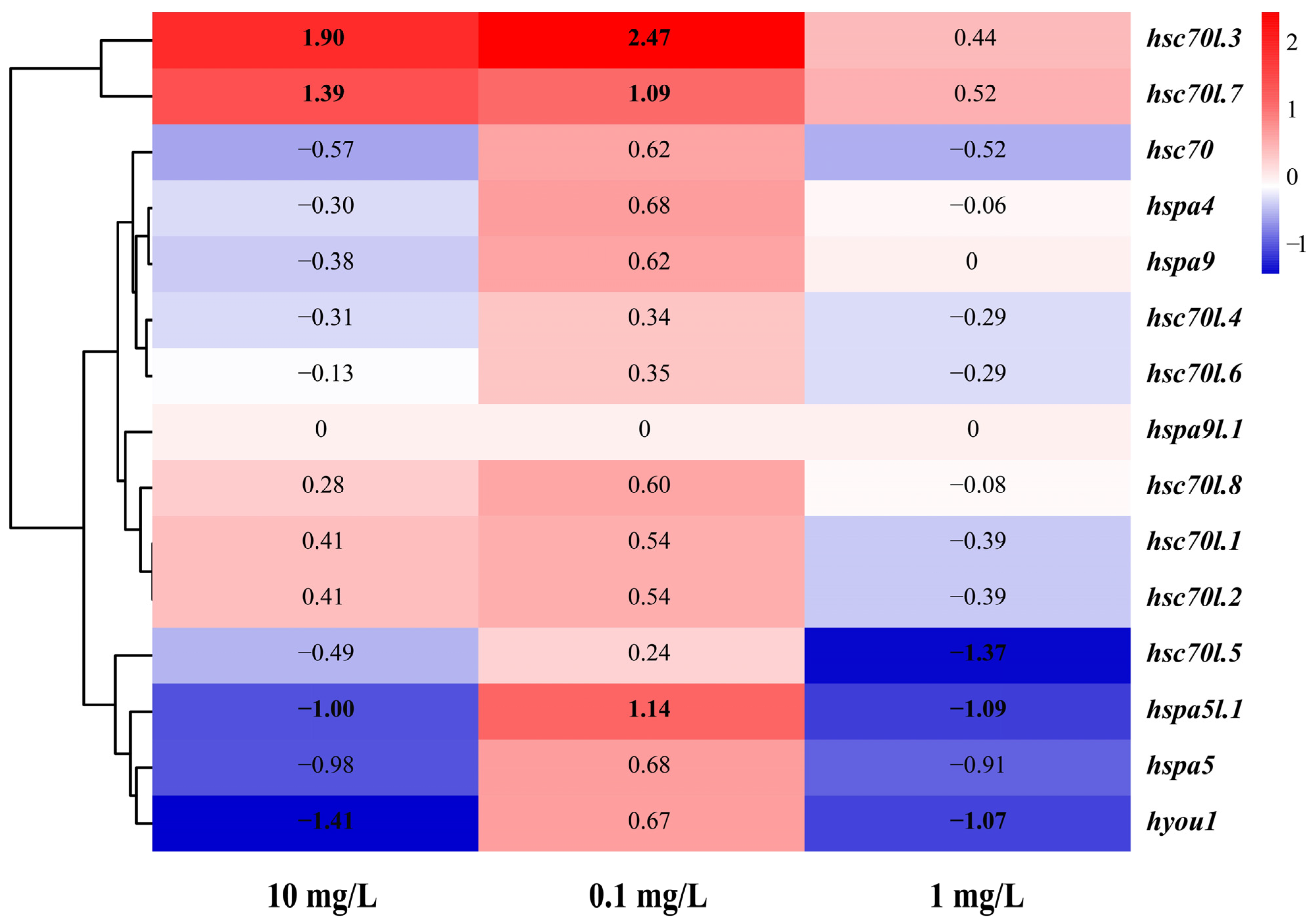
3.7. Expression of the Hsp70 Genes in M. labio Muscles under NP Stress
3.8. Selection Test on Duplicated Hsp70 Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hartl, F.U.; Bracher, A.; Hayer-Hartl, M. Molecular chaperones in protein folding and proteostasis. Nature 2011, 475, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Zatsepina, O.G.; Nikitina, E.A.; Shilova, V.Y.; Chuvakova, L.N.; Sorokina, S.; Vorontsova, J.E.; Tokmacheva, E.V.; Funikov, S.Y.; Rezvykh, A.P.; Evgen’ev, M.B. Hsp70 affects memory formation and behaviorally relevant gene expression in Drosophila melanogaster. Cell Stress Chaperones 2021, 26, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Tiwari, A.K. Molecular chaperone Hsp70 and its constitutively active form Hsc70 play an indispensable role duringeye development of drosophila Melanogaster. Mol Neurobiol. 2018, 55, 4345–4361. [Google Scholar] [PubMed]
- Kennedy, D.; Jäger, R.; Mosser, D.D.; Samali, A. Regulation of apoptosis by heat shock proteins. IUBMB Life 2014, 66, 327–338. [Google Scholar] [CrossRef] [PubMed]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response: Evolutionary and ecological physiology. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef]
- Hu, B.; Li, M.; Yu, X.; Xun, X.; Lu, W.; Li, X.; Li, Y.; Lou, J.; Wang, S.; Zhang, L.; et al. Diverse expression regulation of hsp70 genes in scallops after exposure to toxic alexandrium dinoflagellates. Chemosphere 2019, 234, 62–69. [Google Scholar] [CrossRef]
- Ciechanover, A.; Kwon, Y.T. Protein Quality Control by Molecular Chaperones in Neurodegeneration. Front. Neurosci. 2017, 11, 185. [Google Scholar] [CrossRef]
- Cong, H.; Lei, Y.; Kong, L. The mitochondrial genome of the toothed top shell snail Monodonta labio (Gastropoda: Trochidae): The first complete sequence in the subfamily monodontinae. Mitochondrial DNA B Resour. 2020, 5, 621–622. [Google Scholar] [CrossRef]
- Zhao, D.; Kong, L.-F.; Sasaki, T.; Li, Q. Shell variations in the gastropod, Monodonta labio, in the north-western pacific: The important role of temperature in the evolution process. J. Mar. Biol. Assoc. 2019, 99, 1591–1599. [Google Scholar] [CrossRef]
- Tallec, K.; Huvet, A.; Di Poi, C.; González-Fernández, C.; Lambert, C.; Petton, B.; Le Goïc, N.; Berchel, M.; Soudant, P.; Paul-Pont, I. Nanoplastics impaired oyster free living stages, gametes and embryos. Environ. Pollut. 2018, 242, 1226–1235. [Google Scholar] [CrossRef]
- Li, Z.; Han, Z. Transcriptional Response of Short-Term Nanoplastic Exposure in Monodonta labio. Mar. Pollut. Bull. 2022, 182, 114005. [Google Scholar] [CrossRef]
- Setälä, O.; Fleming-Lehtinen, V.; Lehtiniemi, M. Ingestion and transfer of microplastics in the planktonic food web. Environ. Pollut. 2014, 185, 77–83. [Google Scholar] [CrossRef]
- Gupta, S.C.; Sharma, A.; Mishra, M.; Mishra, R.K.; Chowdhuri, D.K. Heat shock proteins in toxicology: How close and how far? Life Sci. 2010, 86, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Sun, S.; Gao, T.; Han, Z. The Hsp70 gene family in Boleophthalmus pectinirostris: Genome-wide identification and expression analysis under high ammonia stress. Animals 2019, 9, 36. [Google Scholar] [CrossRef] [PubMed]
- Xu, K.; Xu, H.; Han, Z. Genome-wide identification of Hsp70 genes in the large yellow croaker (Larimichthys crocea) and their regulated expression under cold and heat stress. Genes 2018, 9, 590. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.; Deng, Z.; Xu, S.; Zhang, H.; Han, Z. Genome-wide identification and low-salinity stress analysis of the Hsp70 gene family in swimming crab (Portunus trituberculatus). Int. J. Biol. Macromol. 2022, 208, 126–135. [Google Scholar] [CrossRef]
- Gao, L.; Yuan, Z.; Yu, S.; Yang, Y.; Li, Y.; He, C. Genome-wide identification of HSP70/110 genes in sea cucumber Apostichopus Japonicus and comparative analysis of their involvement in aestivation. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 28, 162–171. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, T.; Koyanagi, R.; Gyoja, F.; Kanda, M.; Hisata, K.; Fujie, M.; Goto, H.; Yamasaki, S.; Nagai, K.; Morino, Y.; et al. Bivalve-specific gene expansion in the pearl oyster genome: Implications of adaptation to a sessile lifestyle. Zool. Lett. 2016, 2, 3. [Google Scholar] [CrossRef] [PubMed]
- Brocchieri, L.; Conway de Macario, E.; Macario, A.J.L. Hsp70 Genes in the Human Genome: Conservation and Differentiation Patterns Predict a Wide Array of Overlapping and Specialized Functions. BMC Evol. Biol. 2008, 8, 19. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Xun, X.; Kong, Y.; Wang, S.; Yang, Z.; Li, Y.; Kong, D.; Wang, S.; Zhang, L.; Hu, X.; et al. Hsp70 gene expansions in the scallop Patinopecten yessoensis and their expression regulation after exposure to the toxic dinoflagellate alexandrium catenella. Fish Shellfish Immunol. 2016, 58, 266–273. [Google Scholar] [CrossRef]
- Zhang, G.; Fang, X.; Guo, X.; Li, L.; Luo, R.; Xu, F.; Yang, P.; Zhang, L.; Wang, X.; Qi, H.; et al. The oyster genome reveals stress adaptation and complexity of shell formation. Nature 2012, 490, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Eddy, S.R. Accelerated profile HMM searches. PLoS Comput. Biol. 2011, 7, e1002195. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef] [PubMed]
- Song, L.; Li, C.; Xie, Y.; Liu, S.; Zhang, J.; Yao, J.; Jiang, C.; Li, Y.; Liu, Z. Genome-wide identification of Hsp70 genes in channel catfish and their regulated expression after bacterial infection. Fish Shellfish Immunol. 2016, 49, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML-VI-HPC: Maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 2006, 22, 2688–2690. [Google Scholar] [CrossRef] [PubMed]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant. 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Wen, L.; Gao, X.; Jin, C.; Xue, Y.; Yao, X. DOG 1.0: Illustrator of protein domain structures. Cell Res. 2009, 19, 271–273. [Google Scholar] [CrossRef] [PubMed]
- Waterhouse, A.M.; Procter, J.B.; Martin, D.M.A.; Clamp, M.; Barton, G.J. Jalview version 2—A multiple sequence alignment editor and analysis workbench. Bioinformatics 2009, 25, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Zhang, Y.; Zhang, Z.; Zhu, J.; Yu, J. KaKs_Calculator 2.0: A toolkit incorporating gamma-series methods and sliding window strategies. Genom. Proteom. Bioinform. 2010, 8, 77–80. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Li, Z.-D.; Li, D.-T.; Jiang, M.-X.; Zhang, C.-X. HSP70/DNAJ family of genes in the brown planthopper, Nilaparvata lugens: Diversity and function. Genes 2021, 12, 394. [Google Scholar] [CrossRef] [PubMed]
- Rehman, A.; Atif, R.M.; Qayyum, A.; Du, X.; Hinze, L.; Azhar, M.T. Genome-wide identification and characterization of HSP70 gene family in four species of cotton. Genomics 2020, 112, 4442–4453. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, D.; Upadhyay, R.C.; Chaudhary, U.B.; Kumar, R.; Singh, S.; Ashutosh Polley, S.; Mukherjee, A.; Das, T.K.; De, S. Seasonal variation in expression pattern of genes under HSP70. Cell Stress Chaperones 2014, 19, 401–408. [Google Scholar] [CrossRef]
- Sung, D.Y.; Vierling, E.; Guy, C.L. Comprehensive expression profile analysis of the arabidopsis Hsp70 gene family. Plant Physiol. 2001, 126, 789–800. [Google Scholar] [CrossRef] [PubMed]
- Jung, K.-H.; Gho, H.-J.; Nguyen, M.X.; Kim, S.-R.; An, G. Genome-wide expression analysis of HSP70 family genes in rice and identification of a cytosolic HSP70 gene highly induced under heat stress. Funct. Integr. Genom. 2013, 13, 391–402. [Google Scholar] [CrossRef] [PubMed]
- Freeling, M. Bias in plant gene content following different sorts of duplication: Tandem, whole-genome, segmental, or by transposition. Annu. Rev. Plant Biol. 2009, 60, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Jin, S.; Shou, C.; Han, Z. Hsp70 Gene Family in Sebastiscus marmoratus: The Genome-Wide Identification and Transcriptome Analysis under Thermal Stress. Genes 2023, 14, 1779. [Google Scholar] [CrossRef] [PubMed]
- Glasauer, S.M.K.; Neuhauss, S.C.F. Whole-Genome Duplication in Teleost Fishes and Its Evolutionary Consequences. Mol. Genet. Genom. 2014, 289, 1045–1060. [Google Scholar] [CrossRef] [PubMed]
- Holland, P.W.H.; Marlétaz, F.; Maeso, I.; Dunwell, T.L.; Paps, J. New Genes from Old: Asymmetric Divergence of Gene Duplicates and the Evolution of Development. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2017, 372, 20150480. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhang, Y.; Liu, Y.; Zhang, Y.; Xiao, S.; Yu, Z. Co-Expression of Heat Shock Protein (HSP) 40 and HSP70 in Pinctada martensii Response to Thermal, Low Salinity and Bacterial Challenges. Fish Shellfish Immunol. 2016, 48, 239–243. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Gene Name | Gene ID | CDS * Length (bp) | Protein Length (aa) | Hsp70 Domain Location (aa) | MW * (kDa) | PI * | Chromosome | Location |
|---|---|---|---|---|---|---|---|---|---|
| 1 | hsc70 | Mlab0082280.1 | 1959 | 652 | 6-612 | 71,385.57 | 5.24 | chr6 | 29612442:29616687 |
| 2 | hsc70l.1 | Mlab0108740.1 | 1905 | 634 | 9-612 | 69,628.69 | 5.51 | chr16 | 3599462:3601643 |
| 3 | hsc70l.2 | Mlab0108730.1 | 1905 | 634 | 9-612 | 69,628.69 | 5.51 | chr16 | 3603044:3604948 |
| 4 | hsc70l.3 | Mlab0108940.1 | 1905 | 634 | 9-612 | 69,483.42 | 5.39 | chr16 | 3103368:3111202 |
| 5 | hsc70l.4 | Mlab0108950.1 | 1905 | 634 | 9-612 | 69,582.56 | 5.45 | chr16 | 3099140:3101222 |
| 6 | hsc70l.5 | Mlab0079900.1 | 1926 | 641 | 9-612 | 70,145.10 | 5.64 | chr3 | 2896965:2899237 |
| 7 | hsc70l.6 | Mlab0033790.1 | 1905 | 634 | 9-612 | 69,631.49 | 5.56 | chr9 | 11909551:11911455 |
| 8 | hsc70l.7 | Mlab0033800.1 | 1905 | 634 | 9-612 | 69,608.45 | 5.51 | chr9 | 11912615:11914519 |
| 9 | hsc70l.8 | Mlab0114000.1 | 1902 | 633 | 6-609 | 69,727.77 | 5.55 | chr10 | 4284735:4288100 |
| 10 | hspa4 | Mlab0150810.1 | 2577 | 858 | 3-707 | 96,570.18 | 5.18 | chr10 | 34117494:34130454 |
| 11 | hspa5 | Mlab0102200.1 | 2004 | 667 | 39-644 | 73,546.11 | 5.03 | chr14 | 17073207:17077361 |
| 12 | hspa5l.1 | Mlab0102100.1 | 1980 | 659 | 34-640 | 72,963.32 | 5.38 | chr14 | 16986162:16990290 |
| 13 | hspa9 | Mlab0147970.1 | 2094 | 697 | 61-659 | 76,155.32 | 5.92 | chr11 | 26113170:26125780 |
| 14 | hspa9l.1 | Mlab0204400.1 | 1776 | 591 | 1-554 | 64,170.27 | 4.86 | ptg000096l | 225489:227341 |
| 15 | hyou1 | Mlab0200990.1 | 2904 | 967 | 22-774 | 109,317.80 | 5.48 | chr18 | 26557938:26579334 |
| Protein | α Helix | β Turn | Random Coil | Extended Strand | Subcellular Location Prediction |
|---|---|---|---|---|---|
| Hsc70 | 40.95% | 7.21% | 33.44% | 18.40% | Cytoplasm |
| Hsc70l.1 | 41.48% | 7.89% | 31.55% | 19.09% | Cytoplasm |
| Hsc70l.2 | 41.48% | 7.89% | 31.55% | 19.09% | Cytoplasm |
| Hsc70l.3 | 41.64% | 7.41% | 32.18% | 18.77% | Cytoplasm |
| Hsc70l.4 | 41.64% | 7.41% | 32.02% | 18.93% | Cytoplasm |
| Hsc70l.5 | 42.28% | 6.40% | 32.61% | 18.72% | Nucleus |
| Hsc70l.6 | 42.43% | 7.10% | 31.39% | 19.09% | Cytoplasm |
| Hsc70l.7 | 42.90% | 7.10% | 31.23% | 18.77% | Cytoplasm |
| Hsc70l.8 | 42.81% | 6.79% | 32.07% | 18.33% | Cytoplasm |
| Hspa4 | 44.06% | 3.03% | 38.81% | 14.10% | Nucleus |
| Hspa5 | 43.03% | 7.20% | 31.63% | 18.14% | Endoplasmic reticulum |
| Hspa5l.1 | 42.49% | 6.98% | 31.26% | 19.27% | Mitochondrion |
| Hspa9 | 44.19% | 7.32% | 29.56% | 18.94% | Mitochondrion |
| Hspa9l.1 | 42.47% | 6.77% | 29.44% | 21.32% | Cytoplasm |
| Hyou1 | 47.05% | 4.65% | 33.92% | 14.37% | Endoplasmic reticulum |
| Gene | log2FC | ||
|---|---|---|---|
| 0.1 mg/L | 1 mg/L | 10 mg/L | |
| hsc70 | 0.62 | −0.57 | −0.52 |
| hsc70l.1 | 0.54 | 0.41 | −0.39 |
| hsc70l.2 | 0.54 | 0.41 | −0.39 |
| hsc70l.3 | 2.47 | 1.90 | 0.44 |
| hsc70l.4 | 0.34 | −0.31 | −0.29 |
| hsc70l.5 | 0.24 | −0.49 | −1.37 |
| hsc70l.6 | 0.35 | −0.13 | −0.29 |
| hsc70l.7 | 1.09 | 1.39 | 0.52 |
| hsc70l.8 | 0.60 | 0.28 | −0.08 |
| hspa4 | 0.68 | −0.30 | −0.06 |
| hspa5 | 0.68 | −0.98 | −0.91 |
| hspa5l.1 | 1.14 | −1.00 | −1.09 |
| hspa9 | 0.62 | −0.38 | 0.00 |
| hspa9l.1 | 0.00 | 0.00 | 0.00 |
| hyou1 | 0.67 | −1.41 | −1.07 |
| Gene-Pair | Ka | Ks | Ka/Ks |
|---|---|---|---|
| hsc70l.2–hsc70l.1 | 0 | 0 | 0 |
| hsc70l.3–hsc70l.4 | 0.0008 | 0.0064 | 0.1244 |
| hsc70l.6–hsc70l.7 | 0.0007 | 0.0139 | 0.0495 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Han, X.; Sun, S.; Jin, S.; Liu, M.; Han, Z. Genome-Wide Identification and Transcriptome Analysis of the Hsp70 Gene Family in Monodonta labio Reveals Its Role in Response to Nanoplastics Stress. Genes 2024, 15, 291. https://doi.org/10.3390/genes15030291
He J, Han X, Sun S, Jin S, Liu M, Han Z. Genome-Wide Identification and Transcriptome Analysis of the Hsp70 Gene Family in Monodonta labio Reveals Its Role in Response to Nanoplastics Stress. Genes. 2024; 15(3):291. https://doi.org/10.3390/genes15030291
Chicago/Turabian StyleHe, Jingjing, Xiaolu Han, Shaolei Sun, Shihuai Jin, Mengyuan Liu, and Zhiqiang Han. 2024. "Genome-Wide Identification and Transcriptome Analysis of the Hsp70 Gene Family in Monodonta labio Reveals Its Role in Response to Nanoplastics Stress" Genes 15, no. 3: 291. https://doi.org/10.3390/genes15030291
APA StyleHe, J., Han, X., Sun, S., Jin, S., Liu, M., & Han, Z. (2024). Genome-Wide Identification and Transcriptome Analysis of the Hsp70 Gene Family in Monodonta labio Reveals Its Role in Response to Nanoplastics Stress. Genes, 15(3), 291. https://doi.org/10.3390/genes15030291