
The Impact of Cancer-Associated Fibroblasts on the Biology and Progression of Colorectal Carcinomas

 , , , , , and
, , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Patient Cohort and Study Design
2.2. Isolation and Quantification of Nucleic Acids
2.3. Digital Gene Expression
2.4. NanoString Data Processing
2.5. Next Generation Sequencing
2.6. Immunohistochemistry
2.7. Statistical Evaluation
3. Results
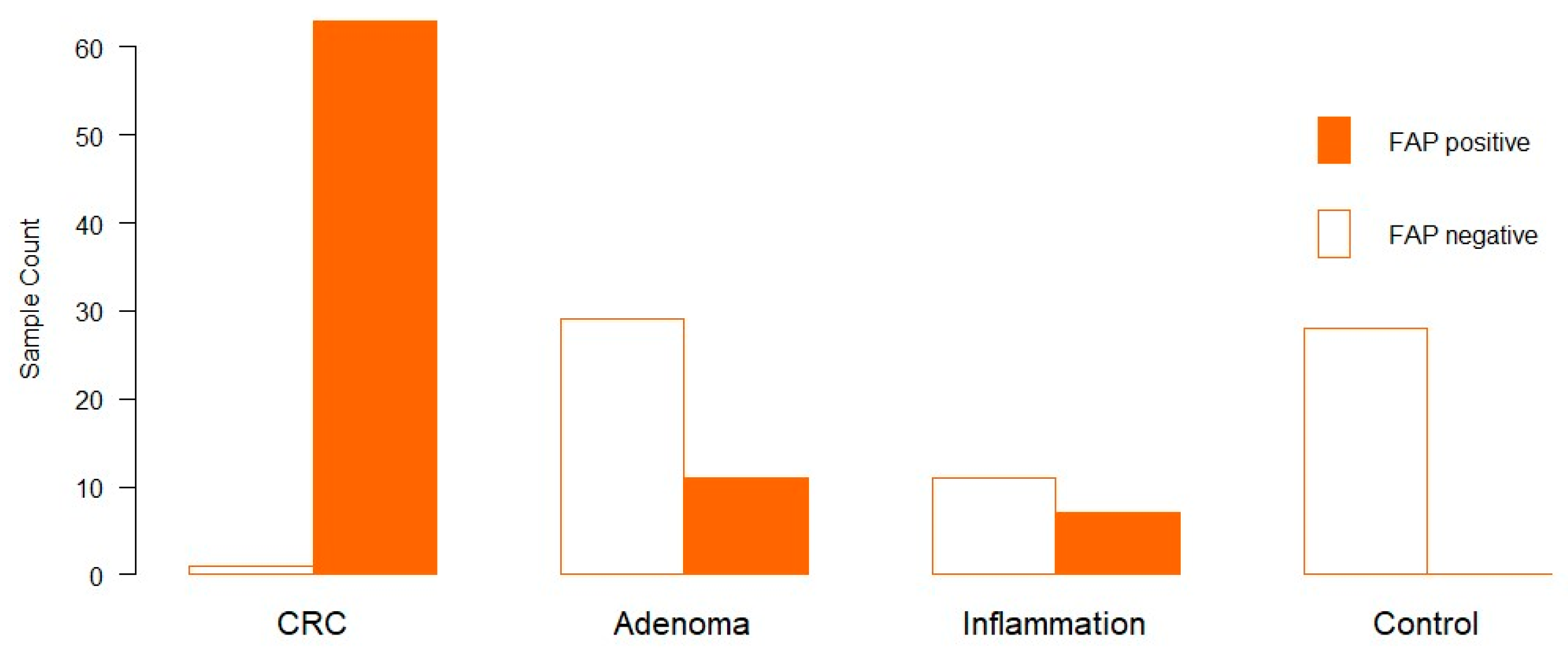
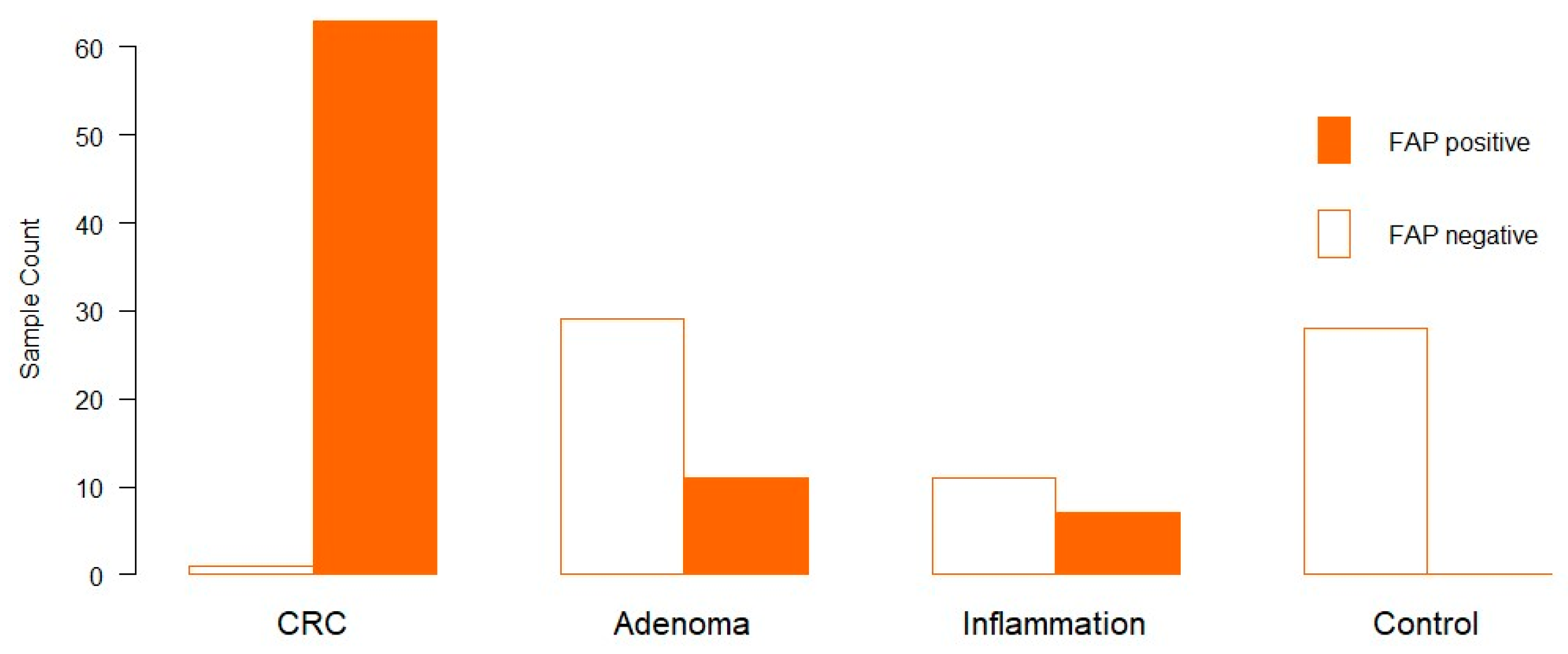
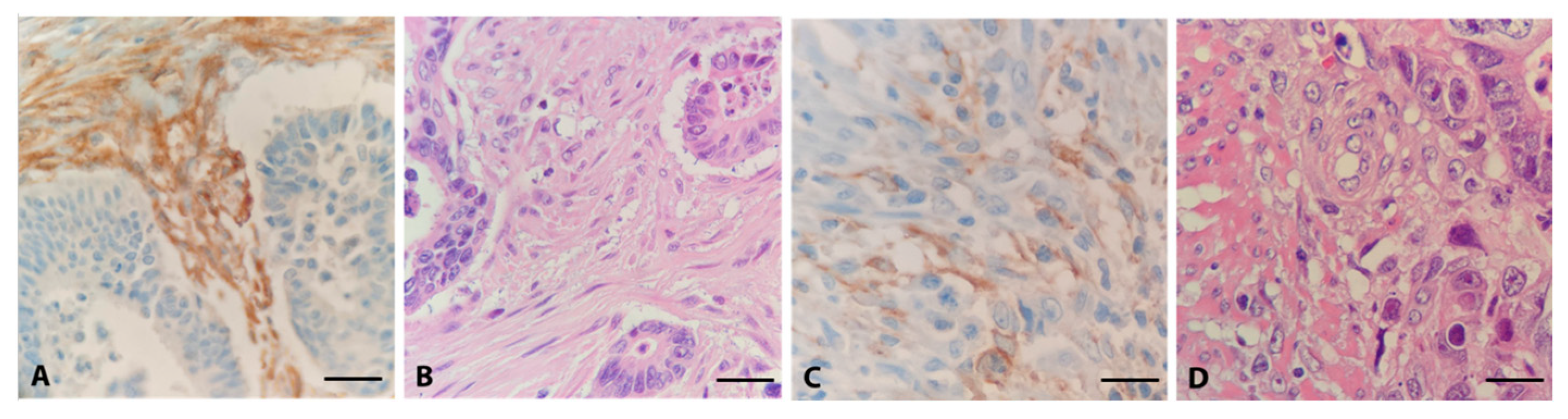
3.1. Distribution of FAP in Clinical Samples via Immunohistochemistry
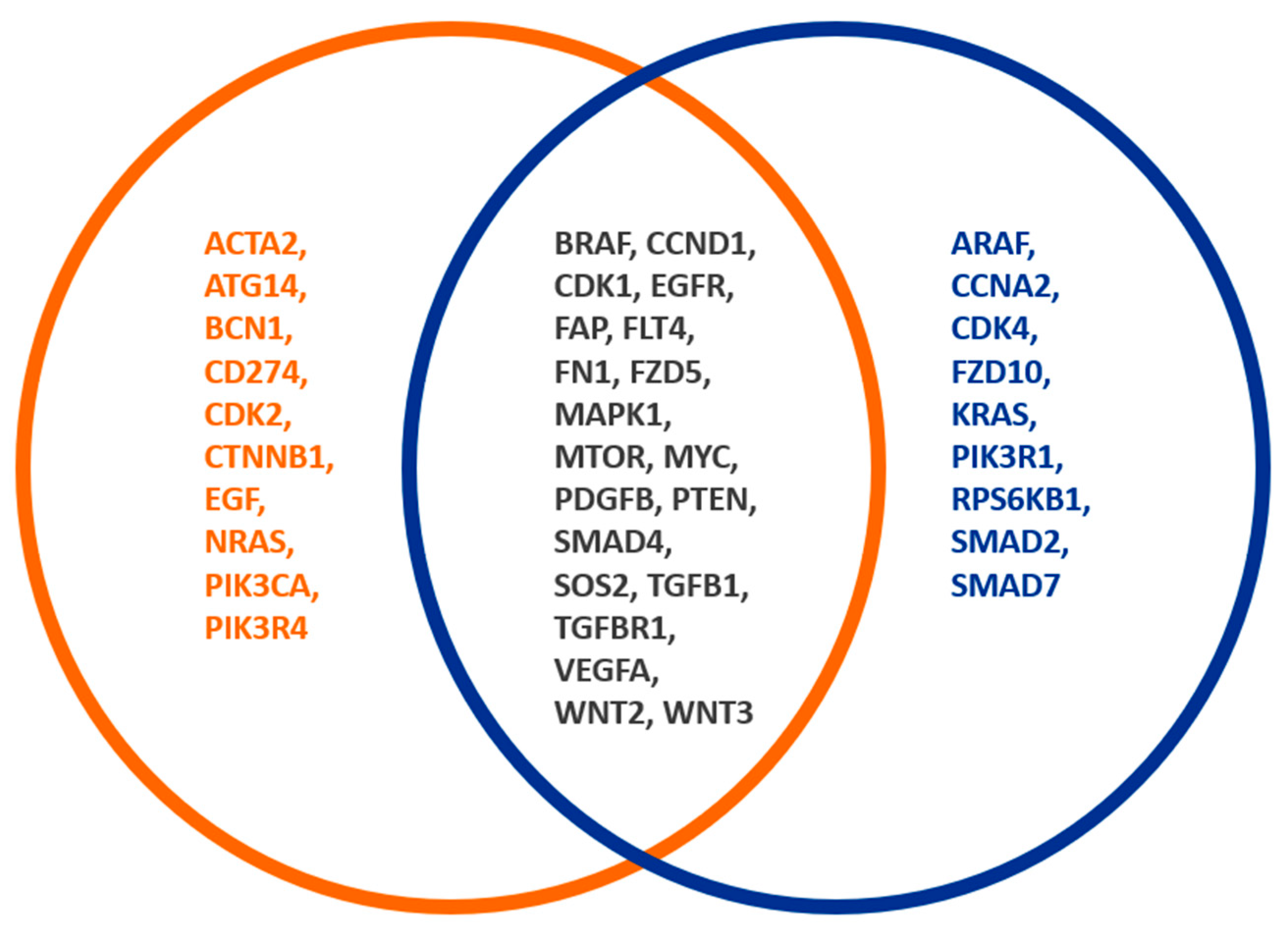
3.2. Comparison of the Gene Expression Patterns of Tumours with Low and High Infiltration by FAP-Positive Fibroblasts
3.3. Adenoma vs. CRC
4. Discussion
4.1. Active Fibroblasts Are Abundant in Colorectal Cancer
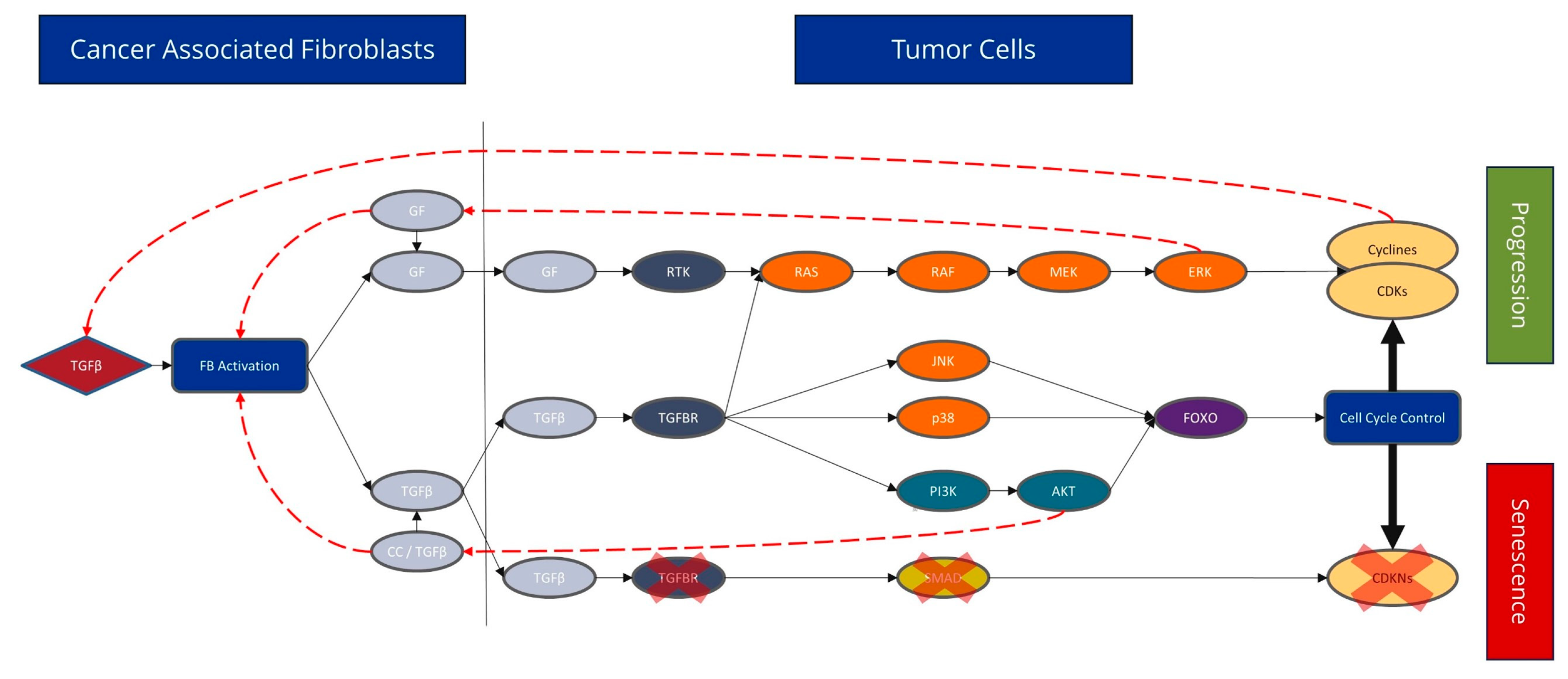
4.2. CAFs Have a Major Influence on the Cell Cycle Signalling Pathway
4.3. Different Expression Patterns of CRC and Adenomas
4.4. The Detrimental Impact of CAFs on Clinical Course, Survival Rate, and Treatment Response
4.5. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Herold, G. Innere Medizin 2021; De Gruyter: Berlin, Germany, 2021. [Google Scholar]
- Xi, Y.; Xu, P. Global colorectal cancer burden in 2020 and projections to 2040. Transl. Oncol. 2021, 14, 101174. [Google Scholar] [CrossRef] [PubMed]
- Mattiuzzi, C.; Sanchis-Gomar, F.; Lippi, G. Concise update on colorectal cancer epidemiology. Ann. Transl. Med. 2019, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Musa, M.; Ali, A. Cancer-associated fibroblasts of colorectal cancer and their markers: Updates, challenges and translational outlook. Future Oncol. 2020, 16, 2329–2344. [Google Scholar] [CrossRef] [PubMed]
- Guinney, J.; Dienstmann, R.; Wang, X.; de Reyniès, A.; Schlicker, A.; Soneson, C.; Marisa, L.; Roepman, P.; Nyamundanda, G.; Angelino, P.; et al. The consensus molecular subtypes of colorectal cancer. Nat. Med. 2015, 21, 1350–1356. [Google Scholar] [CrossRef] [PubMed]
- Fridman, W.H.; Miller, I.; Sautès-Fridman, C.; Byrne, A.T. Therapeutic Targeting of the Colorectal Tumour Stroma. Gastroenterology 2020, 158, 303–321. [Google Scholar] [CrossRef] [PubMed]
- D’Arcangelo, E.; Wu, N.C.; Cadavid, J.L.; McGuigan, A.P. The life cycle of cancer-associated fibroblasts within the tumour stroma and its importance in disease outcome. Br. J. Cancer 2020, 122, 931–942. [Google Scholar] [CrossRef]
- Kobayashi, H.; Enomoto, A.; Woods, S.L.; Burt, A.D.; Takahashi, M.; Worthley, D.L. Cancer-associated fibroblasts in gastrointestinal cancer. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 282–295. [Google Scholar] [CrossRef]
- Fitzgerald, A.A.; Weiner, L.M. The role of fibroblast activation protein in health and malignancy. Cancer Metastasis Rev. 2020, 39, 783–803. [Google Scholar] [CrossRef]
- Altmann, A.; Haberkorn, U.; Siveke, J. The latest developments in imaging of fibroblast activation protein. J. Nucl. Med. 2021, 62, 160–167. [Google Scholar] [CrossRef]
- Henry, L.R.; Lee, H.-O.; Lee, J.S.; Klein-Szanto, A.; Watts, P.; Ross, E.A.; Chen, W.-T.; Cheng, J.D. Clinical implications of fibroblast activation protein in patients with colon cancer. Clin. Cancer Res. 2007, 13, 1736–1741. [Google Scholar] [CrossRef]
- Remmele, W.; Stegner, H.E. Recommendation for uniform definition of an immunoreactive score (IRS) for immunohistochemical estrogen receptor detection (ER-ICA) in breast cancer tissue. Pathologe 1987, 8, 138–140. [Google Scholar] [PubMed]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Liao, Y.; Wang, J.; Jaehnig, E.J.; Shi, Z.; Zhang, B. WebGestalt 2019: Gene set analysis toolkit with revamped UIs and APIs. Nucleic Acids Res. 2019, 47, W199–W205. [Google Scholar] [CrossRef]
- Coto-Llerena, M.; Ercan, C.; Kancherla, V.; Taha-Mehlitz, S.; Eppenberger-Castori, S.; Soysal, S.D.; Ng, C.K.Y.; Bolli, M.; von Flüe, M.; Nicolas, G.P.; et al. High Expression of FAP in Colorectal Cancer Is Associated With Angiogenesis and Immunoregulation Processes. Front. Oncol. 2020, 10, 979. [Google Scholar] [CrossRef] [PubMed]
- Wikberg, M.L.; Edin, S.; Lundberg, I.V.; Van Guelpen, B.; Dahlin, A.M.; Rutegård, J.; Stenling, R.; Öberg, Å.; Palmqvist, R. High intratumoural expression of fibroblast activation protein (FAP) in colon cancer is associated with poorer patient prognosis. Tumour Biol. 2013, 34, 1013–1020. [Google Scholar] [CrossRef] [PubMed]
- Garin-Chesa, P.; Old, L.J.; Rettig, W.J. Cell surface glycoprotein of reactive stromal fibroblasts as a potential antibody target in human epithelial cancers. Proc. Natl. Acad. Sci. USA 1990, 87, 7235–7239. [Google Scholar] [CrossRef]
- Henriksson, M.L.; Edin, S.; Dahlin, A.M.; Oldenborg, P.A.; Öberg, Å.; Van Guelpen, B.; Rutegård, J.; Stenling, R.; Palmqvist, R. Colorectal cancer cells activate adjacent fibroblasts resulting in FGF1/FGFR3 signalling and increased invasion. Am. J. Pathol. 2011, 178, 1387–1394. [Google Scholar] [CrossRef]
- Ma, T.H.; Gao, C.C.; Xie, R.; Yang, X.Z.; Dai, W.J.; Zhang, J.L.; Yan, W.; Wu, S.N. Predictive values of FAP and HGF for tumour angiogenesis and metastasis in colorectal cancer. Neoplasma 2017, 64, 880–886. [Google Scholar] [CrossRef]
- Kuzet, S.E.; Gaggioli, C. Fibroblast activation in cancer: When seed fertilizes soil. Cell Tissue Res. 2016, 365, 607–619. [Google Scholar] [CrossRef]
- Kalluri, R. The biology and function of fibroblasts in cancer. Nat. Rev. Cancer 2016, 16, 582–598. [Google Scholar] [CrossRef]
- Liu, T.; Zhou, L.; Li, D.; Andl, T.; Zhang, Y. Cancer-Associated Fibroblasts Build and Secure the Tumour Microenvironment. Front. Cell Dev. Biol. 2019, 7, 60. [Google Scholar] [CrossRef]
- Wagener, C.; Müller, O. Molekulare Onkologie: Entstehung, Progression, Klinische Aspekte; Georg Thieme: Stuttgart, Germany, 2009. [Google Scholar]
- Ikeda, K.; Monden, T.; Tsujie, M.; Izawa, H.; Yamamoto, H.; Ohnishi, T.; Ohue, M.; Sekimoto, M.; Tomita, N.; Monden, M. Cyclin D, CDK4 and p16 expression in colorectal cancer. Nihon Rinsho. Jpn. J. Clin. Med. 1996, 54, 1054–1059. [Google Scholar]
- Zhang, P.; Kawakami, H.; Liu, W.; Zeng, X.; Strebhardt, K.; Tao, K.; Huang, S.; Sinicrope, F.A. Targeting CDK1 and MEK/ERK overcomes apoptotic resistance in BRAF-mutant human colorectal cancer. Mol. Cancer Res. 2018, 16, 378–389. [Google Scholar] [CrossRef]
- Elbadawy, M.; Usui, T.; Yamawaki, H.; Sasaki, K. Emerging Roles of C-Myc in Cancer Stem Cell-Related Signalling and Resistance to Cancer Chemotherapy: A Potential Therapeutic Target Against Colorectal Cancer. Int. J. Mol. Sci. 2019, 20, 2340. [Google Scholar] [CrossRef]
- Kramer, N.; Schmöllerl, J.; Unger, C.; Nivarthi, H.; Rudisch, A.; Unterleuthner, D.; Scherzer, M.; Riedl, A.; Artaker, M.; Crncec, I.; et al. Autocrine WNT2 signalling in fibroblasts promotes colorectal cancer progression. Oncogene 2017, 36, 5460–5472. [Google Scholar] [CrossRef]
- Gok Yavuz, B.; Gunaydin, G.; Gedik, M.E.; Kosemehmetoglu, K.; Karakoc, D.; Ozgur, F.; Guc, D. Cancer associated fibroblasts sculpt tumour microenvironment by recruiting monocytes and inducing immunosuppressive PD-1(+) TAMs. Sci. Rep. 2019, 9, 3172. [Google Scholar] [CrossRef] [PubMed]
- Freeman, P.; Mielgo, A. Cancer-Associated Fibroblast Mediated Inhibition of CD8+ Cytotoxic T Cell Accumulation in Tumours: Mechanisms and Therapeutic Opportunities. Cancers 2020, 12, 2687. [Google Scholar] [CrossRef] [PubMed]
- Sahai, E.; Astsaturov, I.; Cukierman, E.; DeNardo, D.G.; Egeblad, M.; Evans, R.M.; Fearon, D.; Greten, F.R.; Hingorani, S.R.; Hunter, T.; et al. A framework for advancing our understanding of cancer-associated fibroblasts. Nat. Rev. Cancer 2020, 20, 174–186. [Google Scholar] [CrossRef] [PubMed]
- Capparelli, C.; Chiavarina, B.; Whitaker-Menezes, D.; Pestell, T.G.; Pestell, R.G.; Hulit, J.; Andò, S.; Howell, A.; Martinez-Outschoorn, U.E.; Sotgia, F.; et al. CDK inhibitors (p16/p19/p21) induce senescence and autophagy in cancer-associated fibroblasts, “fueling” tumour growth via paracrine interactions, without an increase in neo-angiogenesis. Cell Cycle 2012, 11, 3599–3610. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cao, Y.; Townsend, C.M.; Ko, T.C. TGF-β signalling in colon cancer cells. World J. Surg. 2005, 29, 306–311. [Google Scholar] [CrossRef] [PubMed]
- Unterleuthner, D.; Neuhold, P.; Schwarz, K.; Janker, L.; Neuditschko, B.; Nivarthi, H.; Crncec, I.; Kramer, N.; Unger, C.; Hengstschläger, M.; et al. Cancer-associated fibroblast-derived WNT2 increases tumour angiogenesis in colon cancer. Angiogenesis 2020, 23, 159–177. [Google Scholar] [CrossRef] [PubMed]
- Bettington, M.L.; Walker, N.I.; Rosty, C.; Brown, I.S.; Clouston, A.D.; McKeone, D.M.; Pearson, S.-A.; Klein, K.; A Leggett, B.; Whitehall, V.L. A clinicopathological and molecular analysis of 200 traditional serrated adenomas. Mod. Pathol. 2015, 28, 414–427. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Yang, S.; Mack, C.; Xu, H.; Huang, C.S.; Mulcahy, E.; Amorosino, M.; Farraye, F.A. Comparison of microsatellite instability, CpG island methylation phenotype, BRAF and KRAS status in serrated polyps and traditional adenomas indicates separate pathways to distinct colorectal carcinoma end points. Am. J. Surg. Pathol. 2006, 30, 1491–1501. [Google Scholar] [CrossRef]
- Sekine, S.; Yamashita, S.; Yamada, M.; Hashimoto, T.; Ogawa, R.; Yoshida, H.; Taniguchi, H.; Kojima, M.; Ushijima, T.; Saito, Y. Clinicopathological and molecular correlations in traditional serrated adenoma. J. Gastroenterol. 2020, 55, 418–427. [Google Scholar] [CrossRef] [PubMed]
- Hillig, R.C.; Sautier, B.; Schroeder, J.; Moosmayer, D.; Hilpmann, A.; Stegmann, C.M.; Werbeck, N.D.; Briem, H.; Boemer, U.; Weiske, J.; et al. Discovery of potent SOS1 inhibitors that block RAS activation via disruption of the RAS-SOS1 interaction. Proc. Natl. Acad. Sci. USA 2019, 116, 2551–2560. [Google Scholar] [CrossRef]
- Dang, H.; Harryvan, T.J.; Hawinkels, L. Fibroblast Subsets in Intestinal Homeostasis, Carcinogenesis, Tumour Progression, and Metastasis. Cancers 2021, 13, 183. [Google Scholar] [CrossRef]
- Bu, L.; Baba, H.; Yoshida, N.; Miyake, K.; Yasuda, T.; Uchihara, T.; Tan, P.; Ishimoto, T. Biological heterogeneity and versatility of cancer-associated fibroblasts in the tumour microenvironment. Oncogene 2019, 38, 4887–4901. [Google Scholar] [CrossRef] [PubMed]
- Heneberg, P. Paracrine tumour signalling induces transdifferentiation of surrounding fibroblasts. Crit. Rev. Oncol. Hematol. 2016, 97, 303–311. [Google Scholar] [CrossRef]
- Shin, N.; Son, G.M.; Shin, D.-H.; Kwon, M.-S.; Park, B.-S.; Kim, H.-S.; Ryu, D.; Kang, C.-D. Cancer-Associated Fibroblasts and Desmoplastic Reactions Related to Cancer Invasiveness in Patients With Colorectal Cancer. Ann. Coloproctol. 2019, 35, 36–46. [Google Scholar] [CrossRef]
- Son, G.M.; Kwon, M.-S.; Shin, D.-H.; Shin, N.; Ryu, D.; Kang, C.-D. Comparisons of cancer-associated fibroblasts in the intratumoural stroma and invasive front in colorectal cancer. Medicine 2019, 98, e15164. [Google Scholar] [CrossRef]
- Brennen, W.N.; Isaacs, J.T.; Denmeade, S.R. Rationale behind targeting fibroblast activation protein-expressing carcinoma-associated fibroblasts as a novel chemotherapeutic strategy. Mol. Cancer Ther. 2012, 11, 257–266. [Google Scholar] [CrossRef]
- Kakarla, S.; Song, X.T.; Gottschalk, S. Cancer-associated fibroblasts as targets for immunotherapy. Immunotherapy 2012, 4, 1129–1138. [Google Scholar] [CrossRef] [PubMed]
- Zafari, N.; Khosravi, F.; Rezaee, Z.; Esfandyari, S.; Bahiraei, M.; Bahramy, A.; Ferns, G.A.; Avan, A. The role of the tumour microenvironment in colorectal cancer and the potential therapeutic approaches. J. Clin. Lab. Anal. 2022, 36, e24585. [Google Scholar] [CrossRef] [PubMed]
- Thoma, O.M.; Neurath, M.F.; Waldner, M.J. Cyclin-Dependent Kinase Inhibitors and Their Therapeutic Potential in Colorectal Cancer Treatment. Front. Pharmacol. 2021, 12, 757120. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Li, W.; Lin, S.; Liu, B.; Wu, P.; Li, L. Fibroblast diversity and plasticity in the tumour microenvironment: Roles in immunity and relevant therapies. Cell Commun. Signal 2023, 21, 234. [Google Scholar] [CrossRef]
- Tong, J.; Tan, X.; Hao, S.; Ermine, K.; Lu, X.; Liu, Z.; Jha, A.; Yu, J.; Zhang, L. Inhibition of multiple CDKs potentiates colon cancer chemotherapy via p73-mediated DR5 induction. Oncogene 2023, 42, 869–880. [Google Scholar] [CrossRef]
- Piringer, G.; Decker, J.; Trommet, V.; Kühr, T.; Heibl, S.; Dörfler, K.; Thaler, J. Ongoing complete response after treatment cessation with dabrafenib, trametinib, and cetuximab as third-line treatment in a patient with advanced BRAF(V600E) mutated, microsatellite-stable colon cancer: A case report and literature review. Front. Oncol. 2023, 13, 1166545. [Google Scholar] [CrossRef]
- Scanu, A.M.; De Miglio, M.R. Therapeutic Landscapes in Colorectal Carcinoma. Medicina 2023, 59, 821. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Cycle | PI3K Signalling Pathway | MAPK Pathway | WNT Signalling Pathway | Growth Factors | TGF-β | Fibroblast Markers |
|---|---|---|---|---|---|---|
| CDK1 | PIK3C3 | MAP2K1 | CTNNB1 | EGFR | TGFB1 | ACTA2 |
| CDK2 | ATG14 | KRAS | Wnt1 | TGFA | TGFBR1 | FAP |
| CDK4 | PIK3R5 | MAPK1 | Wnt2 | EGFR | TGFBR2 | FN1 |
| CDK6 | PIK3R4 | BRAF | CD47 | HGF | CD44 | |
| MDM2 | PIK3R1 | CHRM3 | CD274 | FGF1 | SMAD4 | |
| TP53 | PIK3CA | HRAS | ABCB1 | VEGFA | SMAD2 | |
| WEE1 | RPS6KB1 | NRAS | Wnt3 | FLT1 | SMAD7 | |
| Myt1 | BECN1 | ARAF | FZD10 | FLT4 | MYC | |
| CDKN2A | AKT1 | SOS1 | FZD2 | KDR | ||
| CDKN1B | MTOR | SOS2 | FZD5 | PDGFB | ||
| CDKN1A | PTEN | NF1 | IGF1 | |||
| CCND1 | RICTOR | MAP2K2 | VEGFC | |||
| CCNA2 | RPTOR | RAF1 | MET | |||
| CCNB3 | ||||||
| CCNE1 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Henrich, L.M.; Greimelmaier, K.; Wessolly, M.; Klopp, N.A.; Mairinger, E.; Krause, Y.; Berger, S.; Wohlschlaeger, J.; Schildhaus, H.-U.; Baba, H.A.; et al. The Impact of Cancer-Associated Fibroblasts on the Biology and Progression of Colorectal Carcinomas. Genes 2024, 15, 209. https://doi.org/10.3390/genes15020209
Henrich LM, Greimelmaier K, Wessolly M, Klopp NA, Mairinger E, Krause Y, Berger S, Wohlschlaeger J, Schildhaus H-U, Baba HA, et al. The Impact of Cancer-Associated Fibroblasts on the Biology and Progression of Colorectal Carcinomas. Genes. 2024; 15(2):209. https://doi.org/10.3390/genes15020209
Chicago/Turabian StyleHenrich, Larissa Maria, Kristina Greimelmaier, Michael Wessolly, Nick Alexander Klopp, Elena Mairinger, Yvonne Krause, Sophia Berger, Jeremias Wohlschlaeger, Hans-Ulrich Schildhaus, Hideo Andreas Baba, and et al. 2024. "The Impact of Cancer-Associated Fibroblasts on the Biology and Progression of Colorectal Carcinomas" Genes 15, no. 2: 209. https://doi.org/10.3390/genes15020209
APA StyleHenrich, L. M., Greimelmaier, K., Wessolly, M., Klopp, N. A., Mairinger, E., Krause, Y., Berger, S., Wohlschlaeger, J., Schildhaus, H.-U., Baba, H. A., Mairinger, F. D., & Borchert, S. (2024). The Impact of Cancer-Associated Fibroblasts on the Biology and Progression of Colorectal Carcinomas. Genes, 15(2), 209. https://doi.org/10.3390/genes15020209