

Genome-Wide Identification of the IQM Gene Family and Their Transcriptional Responses to Abiotic Stresses in Kiwifruit (Actinidia eriantha)
Abstract
1. Introduction
2. Materials and Methods
2.1. Identification of Kiwifruit IQM Proteins
2.2. Phylogenetic Analysis
2.3. Chromosomal Localization and Gene Duplication
2.4. Analysis of the Gene Structures, Conserved Motifs, and Cis-Acting Elements of AeIQM
2.5. Plant Materials’ Growth Conditions, Hormones, and Abiotic Treatments
2.6. RNA Isolation, Quantitative Real-Time PCR, and Heatmap
2.7. Salt Stress Tolerance Assays of IQM Genes in Yeast
2.8. Statistical Analysis
3. Results
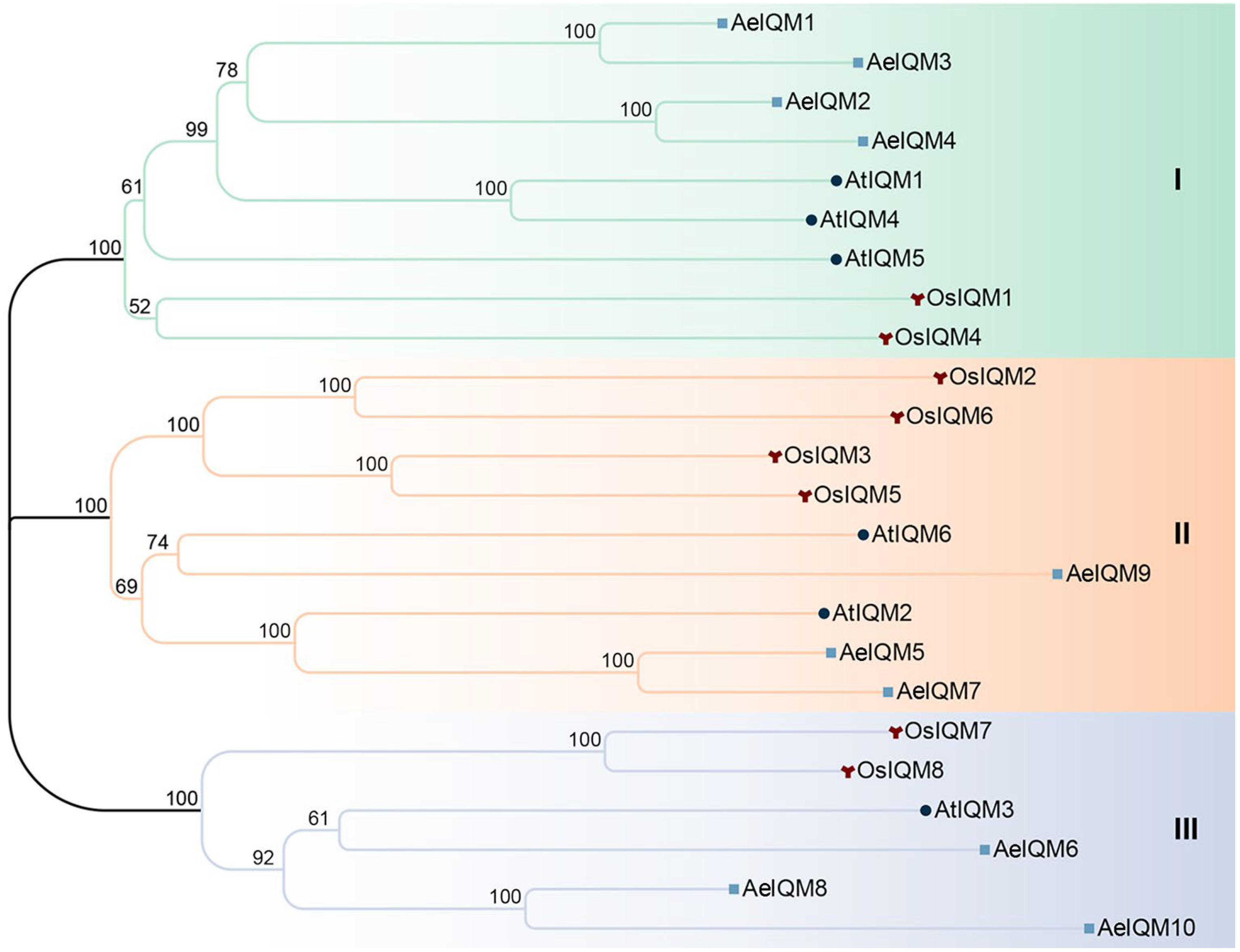
3.1. Identification and Phylogenetic Analyses of IQM Gene Family in Kiwifruit
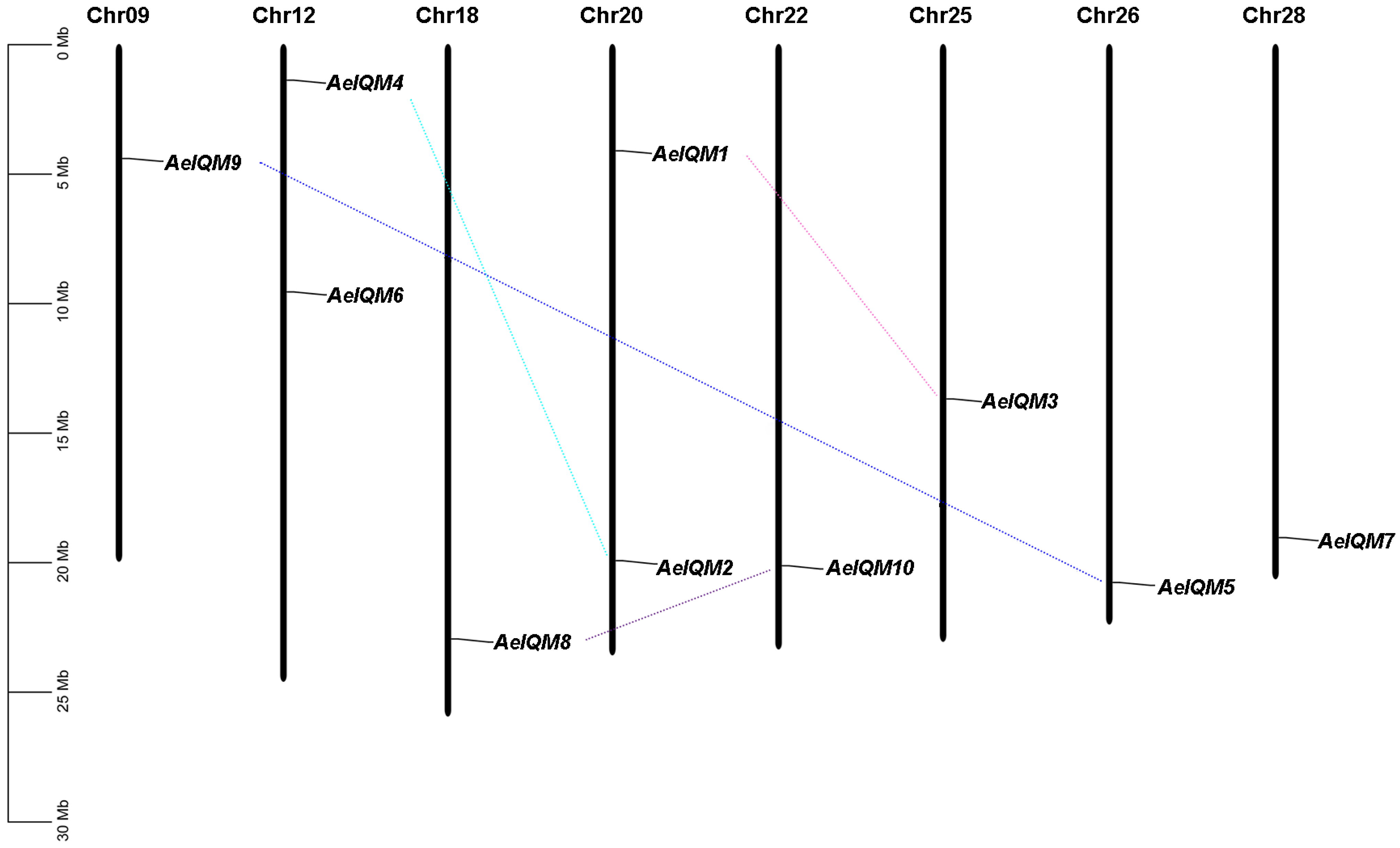
3.2. Chromosomal Locations and Gene Duplication
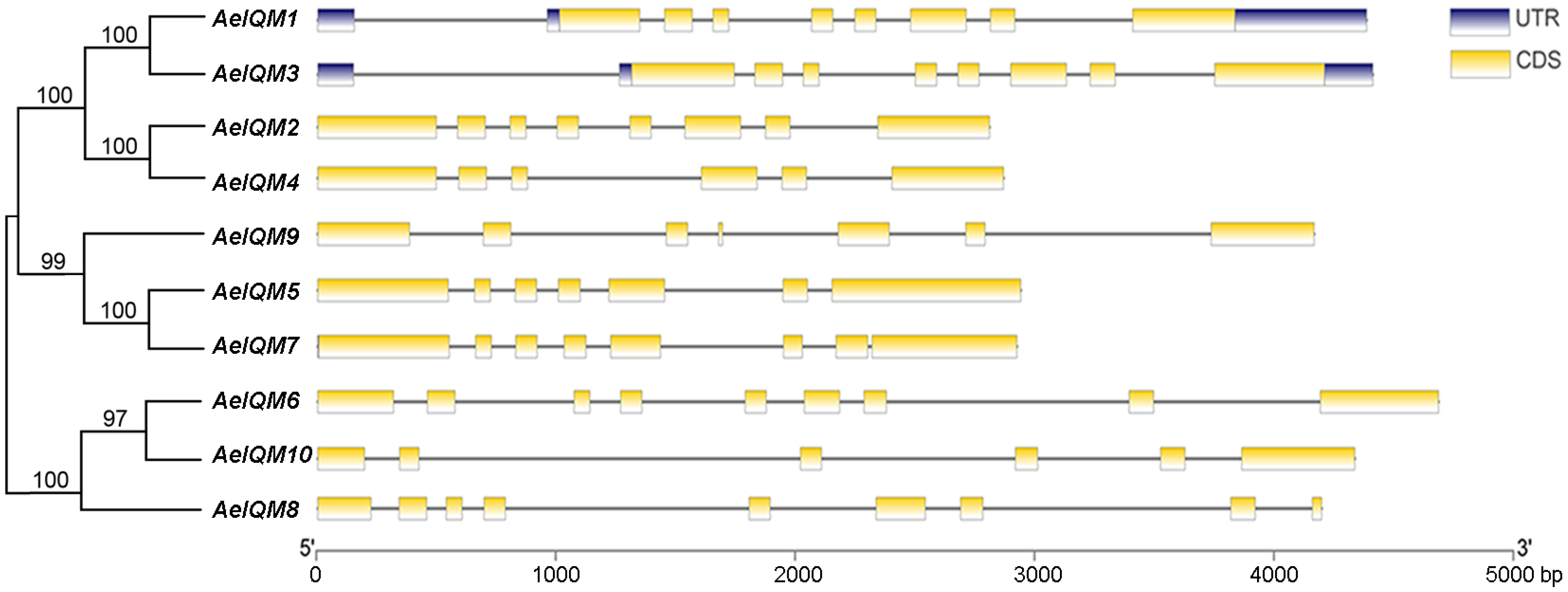
3.3. Gene Structure and Conserved Motifs of Kiwifruit IQM Proteins
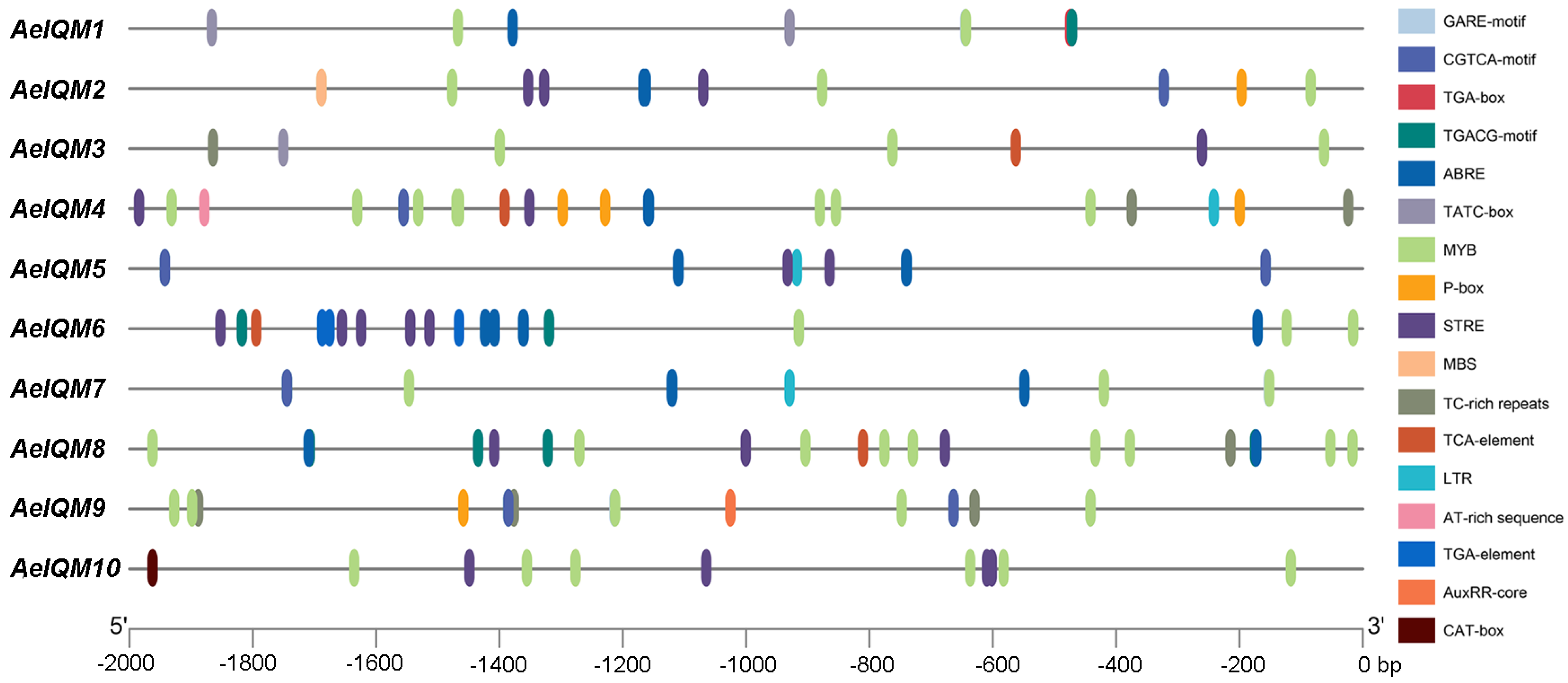
3.4. Analysis of Cis-Acting Elements of AeIQM Gene Promoters
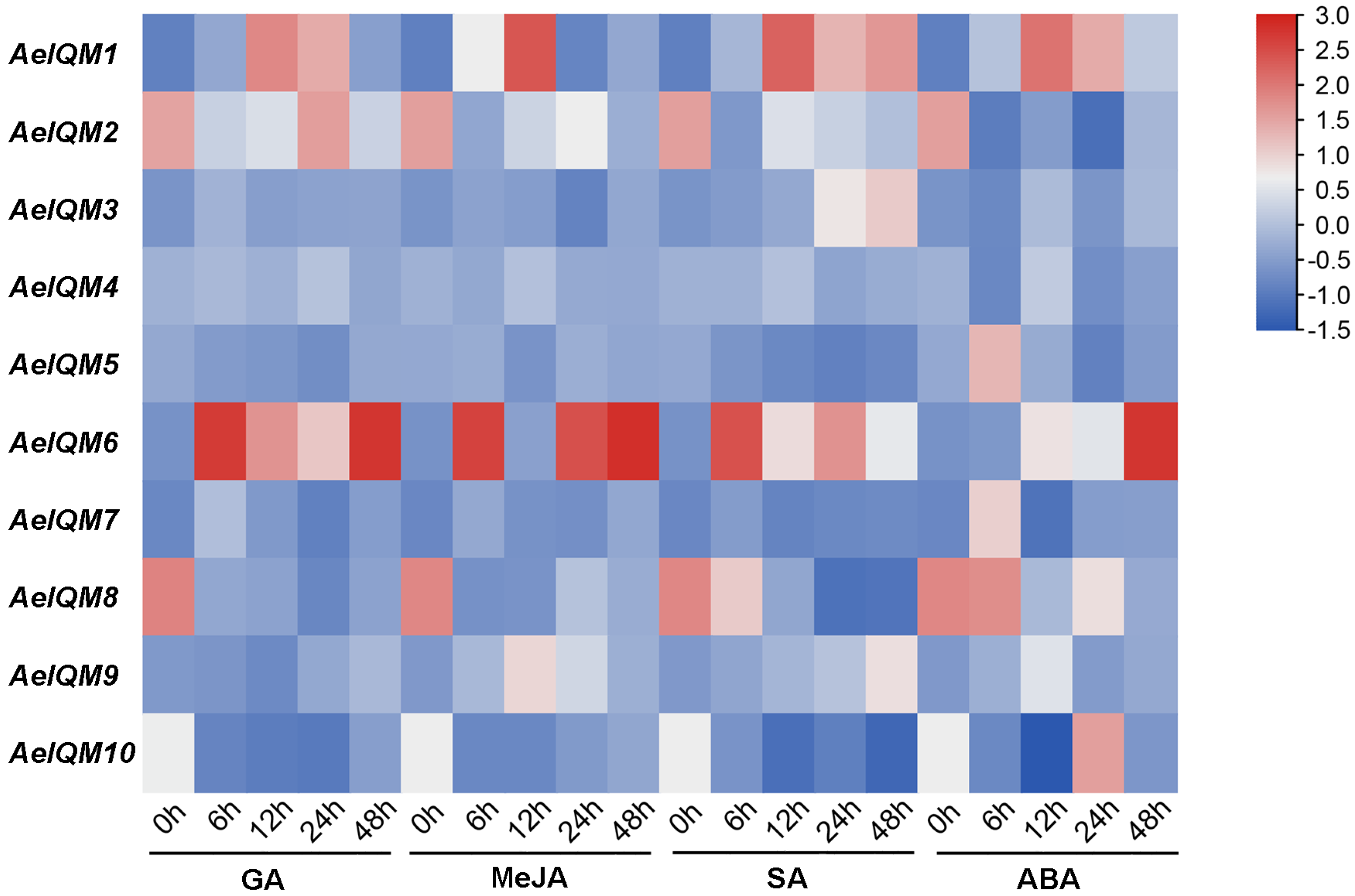
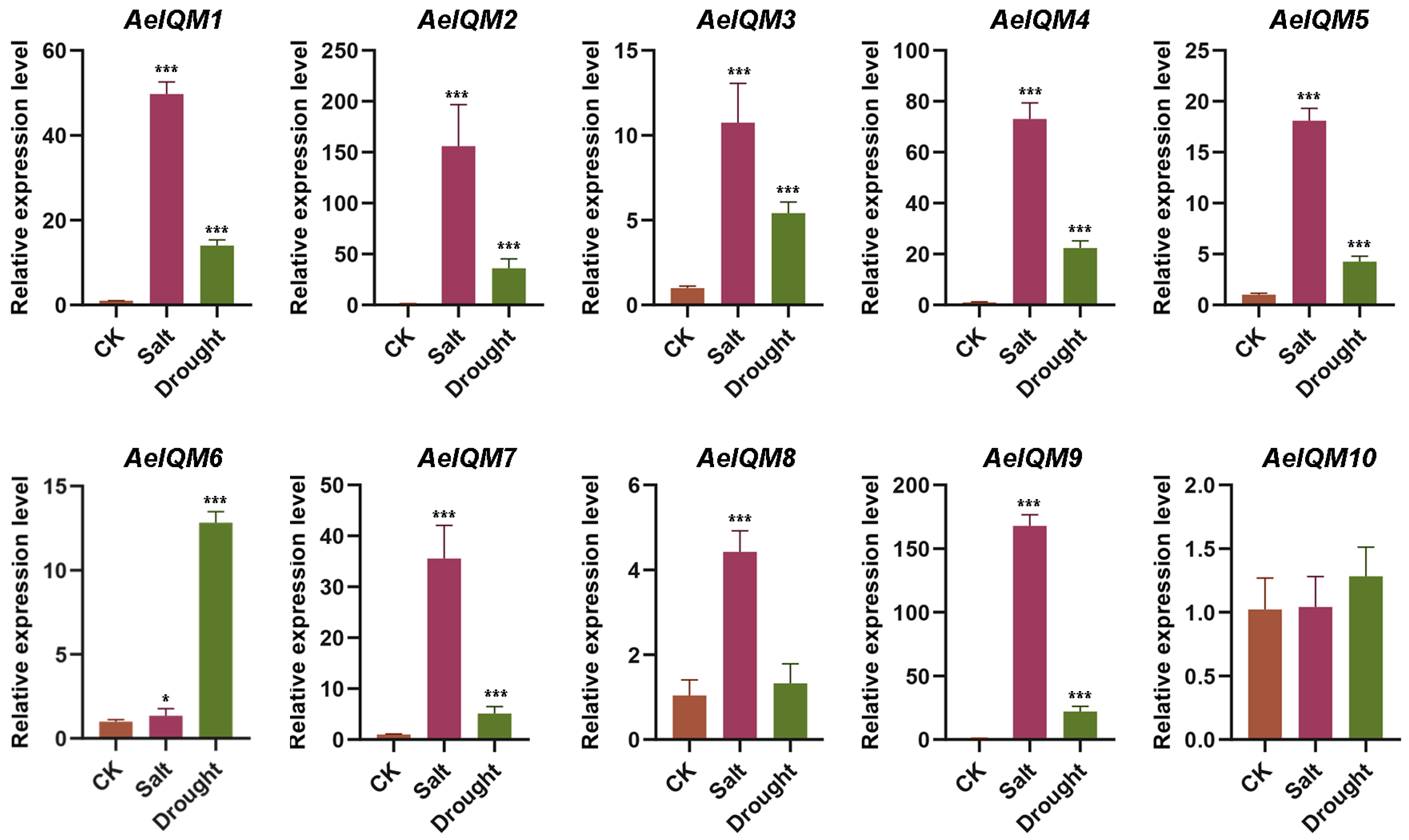
3.5. Effects of Abiotic Stress on AeIQM Gene Expression
3.6. Salt Stress Resistance Analysis of AeIQM Genes in Yeast
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.-K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Liang, W.; Ma, X.; Wan, P.; Liu, L. Plant salt-tolerance mechanism: A review. Biochem. Biophys. Res. Commun. 2018, 495, 286–291. [Google Scholar] [CrossRef]
- Bartels, D.; Sunkar, R. Drought and salt tolerance in plants. CRC Crit. Rev. Plant Sci. 2005, 24, 23–58. [Google Scholar] [CrossRef]
- Hu, S.; Ding, Y.; Zhu, C. Sensitivity and responses of chloroplasts to heat stress in plants. Front. Plant Sci. 2020, 11, 375. [Google Scholar] [CrossRef] [PubMed]
- Manasa, S.; Lekshmi; Planography, M.; Panigrahi, K.C.S.; Rout, G.R. Overview of cold stress regulation in plants. Bot. Rev. 2022, 88, 359–387. [Google Scholar] [CrossRef]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China Life Sci. 2020, 63, 635–674. [Google Scholar] [CrossRef]
- Mustafin, Z.S.; Zamyatin, V.I.; Konstantinov, D.K.; Doroshkov, A.V.; Lashin, S.A.; Afonnikov, D.A. Phylostratigraphic analysis shows the earliest origination of the abiotic stress associated genes in A. thaliana. Genes 2019, 10, 963. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Tuteja, N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol. Biochem. 2010, 48, 909–930. [Google Scholar] [CrossRef]
- Aldon, D.; Mbengue, M.; Mazars, C.; Galaud, J.-P. Calcium signalling in plant biotic interactions. Int. J. Mol. Sci. 2018, 19, 665. [Google Scholar] [CrossRef]
- Hu, W.; Yan, Y.; Tie, W.; Ding, Z.; Wu, C.; Ding, X.; Wang, W.; Xia, Z.; Guo, J.; Peng, M. Genome-wide analyses of calcium sensors reveal their involvement in drought stress response and storage roots deterioration after harvest in Cassava. Genes 2018, 9, 221. [Google Scholar] [CrossRef]
- Carafoli, E.; Krebs, J. Why calcium? How calcium became the best communicator. J. Biol. Chem. 2016, 291, 20849–20857. [Google Scholar] [CrossRef] [PubMed]
- Hetherington, A.M.; Brownlee, C. The generation of Ca2+ signals in plants. Annu. Rev. Plant Biol. 2004, 55, 401–427. [Google Scholar] [CrossRef] [PubMed]
- Chin, D.; Means, A.R. Calmodulin: A prototypical calcium sensor. Trends Cell Biol. 2000, 10, 322–328. [Google Scholar] [CrossRef] [PubMed]
- DeFalco, T.A.; Bender, K.W.; Snedden, W.A. Breaking the code: Ca2+ sensors in plant signalling. Biochem. J. 2009, 425, 27–40. [Google Scholar] [CrossRef]
- Yang, T.; Poovaiah, B.W. Calcium/calmodulin-mediated signal network in plants. Trends Plant Sci. 2003, 8, 505–512. [Google Scholar] [CrossRef]
- Zhou, Y.; Chen, Y.; Yamamoto, K.T.; Duan, J.; Tian, C.-E. Sequence and expression analysis of the Arabidopsis IQM family. Acta Physiol. Plant. 2010, 32, 191–198. [Google Scholar] [CrossRef]
- Abel, S.; Savchenko, T.; Levy, M. Genome-wide comparative analysis of the IQD gene families in Arabidopsis thaliana and Oryza sativa. BMC Evol. Biol. 2005, 5, 72. [Google Scholar] [CrossRef]
- Talke, I.N.; Blaudez, D.; Maathuis, F.J.M.; Sanders, D. CNGCs: Prime targets of plant cyclic nucleotide signalling? Trends Plant Sci. 2003, 8, 286–293. [Google Scholar] [CrossRef]
- Bouché, N.; Scharlat, A.; Snedden, W.; Bouchez, D.; Fromm, H. A novel family of calmodulin-binding transcription activators in multicellular organisms. J. Biol. Chem. 2002, 277, 21851–21861. [Google Scholar] [CrossRef]
- Reddy, A.S.N.; Day, I.S. Analysis of the myosins encoded in the recently completed Arabidopsis thaliana genome sequence. Genome Biol. 2001, 2, research0024. [Google Scholar] [CrossRef]
- Lv, T.; Liu, Q.; Xiao, H.; Fan, T.; Zhou, Y.; Wang, J.; Tian, C.-e. Genome-wide identification and analysis of the IQM gene family in soybean. Front. Plant Sci. 2023, 13, 1093589. [Google Scholar] [CrossRef]
- Fan, T.; Lv, T.; Xie, C.; Zhou, Y.; Tian, C. Genome-wide analysis of the IQM gene family in rice (Oryza sativa L.). Plants 2021, 10, 1949. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.-P.; Duan, J.; Fujibe, T.; Yamamoto, K.T.; Tian, C.-E. AtIQM1, a novel calmodulin-binding protein, is involved in stomatal movement in Arabidopsis. Plant Mol. Biol. 2012, 79, 333–346. [Google Scholar] [CrossRef] [PubMed]
- Lv, T.; Li, X.; Fan, T.; Luo, H.; Xie, C.; Zhou, Y.; Tian, C.E. The calmodulin-binding protein IQM1 interacts with CATALASE2 to affect pathogen defense. Plant Physiol. 2019, 181, 1314–1327. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.P.; Wu, J.H.; Xiao, W.H.; Chen, W.; Chen, Q.H.; Fan, T.; Xie, C.P.; Tian, C.-E. Arabidopsis IQM4, a novel calmodulin-binding protein, is involved with seed dormancy and germination in Arabidopsis. Front. Plant Sci. 2018, 9, 721. [Google Scholar] [CrossRef]
- Gong, L.-P.; Cheng, J.-Z.; Zhou, Y.-P.; Huang, X.-L.; Tian, C.-E. Disruption of IQM5 delays flowering possibly through modulating the juvenile-to-adult transition. Acta Physiol. Plant. 2016, 39, 21. [Google Scholar] [CrossRef]
- Chartzoulakis, K.S.; Therios, I.N.; Misopolinos, N.D.; Noitsakis, B.I. Growth, ion content and photosynthetic performance of salt-stressed kiwifruit plants. Irrig. Sci. 1995, 16, 23–28. [Google Scholar] [CrossRef]
- Sotiropoulos, T.E.; Therios, I.N.; Dimassi, K.N. Uptake of boron by kiwifruit plants under various levels of shading and salinity. J. Plant Nutr. 2005, 27, 1979–1989. [Google Scholar] [CrossRef]
- Klages, K.; Boldingh, H.; Smith, G.S. Accumulation of myo-inositol in Actinidia seedlings subjected to salt stress. Ann. Bot. 1999, 84, 521–527. [Google Scholar] [CrossRef]
- Zhong, Y.P.; Qi, X.-J.; Chen, J.-Y.; Li, Z.; Bai, D.-F.; Wei, C.-G.; Fang, J.-B. Growth and physiological responses of four kiwifruit genotypes to salt stress and resistance evaluation. J. Integr. Agric. 2019, 18, 83–95. [Google Scholar] [CrossRef]
- Abid, M.; Zhang, Y.J.; Li, Z.; Bai, D.F.; Zhong, Y.P.; Fang, J.B. Effect of salt stress on growth, physiological and biochemical characters of four kiwifruit genotypes. Sci. Hortic. 2020, 271, 109473. [Google Scholar] [CrossRef]
- Goodstein, D.M.; Shu, S.; Howson, R.; Neupane, R.; Hayes, R.D.; Fazo, J.; Mitros, T.; Dirks, W.; Hellsten, U.; Putnam, N.; et al. Phytozome: A comparative platform for green plant genomics. Nucleic Acids Res. 2012, 40, D1178–D1186. [Google Scholar] [CrossRef] [PubMed]
- Lei, B.; Song, M.; Li, X.; Dang, X.; Qin, R.; Zhu, S.; An, X.; Liu, Q.; Yao, X.; Nie, Y.; et al. SMART v1.0: A database for small molecules with functional implications in plants. Interdiscip. Sci. 2022, 14, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Chuguransky, S.; Williams, L.; Qureshi, M.; Salazar, G.A.; Sonnhammer, E.L.L.; Tosatto, S.C.E.; Paladin, L.; Raj, S.; Richardson, L.J.; et al. Pfam: The protein families database in 2021. Nucleic Acids Res. 2021, 49, D412–D419. [Google Scholar] [CrossRef] [PubMed]
- Duvaud, S.; Gabella, C.; Lisacek, F.; Stockinger, H.; Ioannidis, V.; Durinx, C. Expasy, the Swiss bioinformatics resource portal, as designed by its users. Nucleic Acids Res. 2021, 49, W216–W227. [Google Scholar] [CrossRef] [PubMed]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mole. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Bailey, T.L.; Williams, N.; Misleh, C.; Li, W.W. MEME: Discovering and analyzing DNA and protein sequence motifs. Nucleic Acids Res. 2006, 34, W369–W373. [Google Scholar] [CrossRef]
- Ling, C.; Liu, Y.; Yang, Z.; Xu, J.; Ouyang, Z.; Yang, J.; Wang, S. Genome-wide identification of HSF gene family in kiwifruit and the function of AeHSFA2b in salt tolerance. Int. J. Mol. Sci. 2023, 24, 15638. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Flagel, L.E.; Wendel, J.F. Gene duplication and evolutionary novelty in plants. New Phytol. 2009, 183, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Ling, C.; Liu, Y.; Zhang, H.; Hussain, Q.; Lyu, S.; Wang, S.; Liu, Y. Genome-wide expression profiling analysis of kiwifruit GolS and RFS genes and identification of AcRFS4 function in raffinose accumulation. Int. J. Mol. Sci. 2022, 23, 8836. [Google Scholar] [CrossRef]
- Abid, M.; Wang, Z.; Feng, C.; Luo, J.; Zhang, Y.; Tu, J.; Cai, X.; Gao, P. Genome-wide identification and structural characterization of growth-regulating factors (GRFs) in Actinida eriantha and Actinidia chinensis. Plants 2022, 11, 1633. [Google Scholar] [CrossRef]
- Pandey, G.K.; Cheong, Y.H.; Kim, K.N.; Grant, J.J.; Li, L.; Hung, W.; D’Angelo, C.; Weinl, S.; Kudla, J.; Luan, S. The calcium sensor calcineurin B-like 9 modulates abscisic acid sensitivity and biosynthesis in Arabidopsis. Plant Cell 2004, 16, 1912–1924. [Google Scholar] [CrossRef]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef]
- Huang, S.; Ding, J.; Deng, D.; Tang, W.; Sun, H.; Liu, D.; Zhang, L.; Niu, X.; Zhang, X.; Meng, M.; et al. Draft genome of the kiwifruit Actinidia chinensis. Nat. Commun. 2013, 4, 2640. [Google Scholar] [CrossRef]
- Wang, J.-P.; Yu, J.-G.; Li, J.; Sun, P.-C.; Wang, L.; Yuan, J.-Q.; Meng, F.-B.; Sun, S.-R.; Li, Y.-X.; Lei, T.-Y.; et al. Two likely auto-tetraploidization events shaped kiwifruit genome and contributed to establishment of the actinidiaceae family. iScience 2018, 7, 230–240. [Google Scholar] [CrossRef]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486–487. [Google Scholar] [CrossRef] [PubMed]
- Wani, S.H.; Kumar, V.; Shriram, V.; Sah, S.K. Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J. 2016, 4, 162–176. [Google Scholar] [CrossRef]
- Kohli, A.; Sreenivasulu, N.; Lakshmanan, P.; Kumar, P.P. The phytohormone crosstalk paradigm takes center stage in understanding how plants respond to abiotic stresses. Plant Cell Rep. 2013, 32, 945–957. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, H.; Song, C.; Zhu, J.K.; Shabala, S. Mechanisms of plant responses and adaptation to soil salinity. Innovation 2020, 1, 100017. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Gene ID | Chr. | Location Coordinates | CDS Length (bp) | Protein Length (aa) | Molecular Weight (kDa) | pI |
|---|---|---|---|---|---|---|---|
| AeIQM1 | DTZ79_20g03230 | Chr20 | 4097436–4101769 (−) | 1446 | 481 | 53.96 | 8.57 |
| AeIQM2 | DTZ79_20g13100 | Chr20 | 19923399–19926174 (+) | 1644 | 547 | 61.58 | 8.39 |
| AeIQM3 | DTZ79_25g04840 | Chr25 | 13677555–13681914 (−) | 1572 | 523 | 58.30 | 8.70 |
| AeIQM4 | DTZ79_12g01010 | Chr12 | 1370000–1372833 (−) | 1467 | 488 | 54.24 | 9.10 |
| AeIQM5 | DTZ79_26g12800 | Chr26 | 20764403–20767307 (+) | 1899 | 632 | 71.54 | 6.98 |
| AeIQM6 | DTZ79_12g06920 | Chr12 | 9544437–9549067 (−) | 1503 | 500 | 56.05 | 6.63 |
| AeIQM7 | DTZ79_28g12820 | Chr28 | 19031981–19034869 (+) | 1806 | 601 | 68.15 | 8.71 |
| AeIQM8 | DTZ79_18g12720 | Chr18 | 22944547–22948694 (+) | 1017 | 338 | 39.15 | 7.99 |
| AeIQM9 | DTZ79_09g03830 | Chr09 | 4394286–4398402 (−) | 1320 | 439 | 50.55 | 5.32 |
| AeIQM10 | DTZ79_22g10670 | Chr22 | 20117392–20121676 (+) | 1026 | 341 | 38.22 | 8.98 |
| NO. | Paralogous Pairs | Ka | Ks | Ka/Ks | Duplicate Type |
|---|---|---|---|---|---|
| 1 | AeIQM1–AeIQM3 | 0.081680638 | 0.209879171 | 0.389179342 | Segmental duplication |
| 2 | AeIQM2–AeIQM4 | 0.051740668 | 0.152338558 | 0.339642625 | Segmental duplication |
| 3 | AeIQM7–AeIQM9 | 0.412184526 | 1.815557259 | 0.227029208 | Segmental duplication |
| 4 | AeIQM8–AeIQM10 | 0.152649613 | 0.210881697 | 0.723863739 | Segmental duplication |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, M.; Zhang, Z.; Ling, C.; Jiao, Y.; Zhang, X. Genome-Wide Identification of the IQM Gene Family and Their Transcriptional Responses to Abiotic Stresses in Kiwifruit (Actinidia eriantha). Genes 2024, 15, 147. https://doi.org/10.3390/genes15020147
Xu M, Zhang Z, Ling C, Jiao Y, Zhang X. Genome-Wide Identification of the IQM Gene Family and Their Transcriptional Responses to Abiotic Stresses in Kiwifruit (Actinidia eriantha). Genes. 2024; 15(2):147. https://doi.org/10.3390/genes15020147
Chicago/Turabian StyleXu, Minyan, Zhi Zhang, Chengcheng Ling, Yuhuan Jiao, and Xin Zhang. 2024. "Genome-Wide Identification of the IQM Gene Family and Their Transcriptional Responses to Abiotic Stresses in Kiwifruit (Actinidia eriantha)" Genes 15, no. 2: 147. https://doi.org/10.3390/genes15020147
APA StyleXu, M., Zhang, Z., Ling, C., Jiao, Y., & Zhang, X. (2024). Genome-Wide Identification of the IQM Gene Family and Their Transcriptional Responses to Abiotic Stresses in Kiwifruit (Actinidia eriantha). Genes, 15(2), 147. https://doi.org/10.3390/genes15020147