
Swedish Genome-Wide Haplotype Association Analysis Suggests Breast Cancer Loci with Varying Risk-Modifying Effects

 , ,
, ,
Abstract
1. Background
2. Material and Methods
2.1. Cases and Controls
2.2. Genotyping and Quality Control
2.3. Statistics
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Available online: https://gco.iarc.who.int/media/globocan/factsheets/populations/752-sweden-fact-sheet.pdf (accessed on 2 November 2024).
- Kurian, A.W.; Ward, K.C.; Howlader, N.; Deapen, D.; Hamilton, A.S.; Mariotto, A.; Miller, D.; Penberthy, L.S.; Katz, S.J. Genetic Testing and Results in a Population-Based Cohort of Breast Cancer Patients and Ovarian Cancer Patients. J. Clin. Oncol. 2019, 37, 1305–1315. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, N.; Ryder, S.; Forbes, C.; Ross, J.; Quek, R.G. A systematic review of the international prevalence of BRCA mutation in breast cancer. Clin. Epidemiol. 2019, 11, 543–561. [Google Scholar] [CrossRef] [PubMed]
- The Breast Cancer Susceptibility Collaboration (UK); Rahman, N.; Seal, S.; Thompson, D.; Kelly, P.; Renwick, A.; Elliott, A.; Reid, S.; Spanova, K.; Barfoot, R.; et al. PALB2, which encodes a BRCA2-interacting protein, is a breast cancer susceptibility gene. Nat. Genet. 2007, 39, 165–167. [Google Scholar] [CrossRef]
- The Breast Cancer Susceptibility Collaboration (UK); Renwick, A.; Thompson, D.; Seal, S.; Kelly, P.; Chagtai, T.; Ahmed, M.; North, B.; Jayatilake, H.; Barfoot, R.; et al. ATM mutations that cause ataxia-telangiectasia are breast cancer susceptibility alleles. Nat. Genet. 2006, 38, 873–875. [Google Scholar] [CrossRef]
- Meijers-Heijboer, H.; van den Ouweland, A.; Klijn, J.; Wasielewski, M.; de Snoo, A.; Oldenburg, R.; Hollestelle, A.; Houben, M.; Crepin, E.; van Veghel-Plandsoen, M.; et al. Low-penetrance susceptibility to breast cancer due to CHEK2(*)1100delC in noncarriers of BRCA1 or BRCA2 mutations. Nat. Genet. 2002, 31, 55–59. [Google Scholar] [CrossRef]
- Michailidou, K.; Lindstrom, S.; Dennis, J.; Beesley, J.; Hui, S.; Kar, S.; Lemacon, A.; Soucy, P.; Glubb, D.; Rostamianfar, A.; et al. Association analysis identifies 65 new breast cancer risk loci. Nature 2017, 551, 92–94. [Google Scholar] [CrossRef]
- Wang, A.; Shen, J.; Rodriguez, A.A.; Saunders, E.J.; Dadaev, T.; Conti, D.V.; Rybicki, B.A.; Terao, C.; Yamaguchi, M.; Andiappan, A.K.; et al. Characterizing prostate cancer risk through multi-ancestry genome-wide discovery of 187 novel risk variants. Nat. Genet. 2023, 55, 2065–2074. [Google Scholar] [CrossRef]
- Dareng, E.O.; Coetzee, S.G.; Tyrer, J.P.; Intermaggio, M.P.; Aben, K.K.H.; Anton-Culver, H.; Bandera, E.V.; Beckmann, M.W.; Berchuck, A.; Bernardini, M.Q.; et al. Integrative multi-omics analyses to identify the genetic and functional mechanisms underlying ovarian cancer risk regions. Am. J. Hum. Genet. 2024, 111, 1061–1083. [Google Scholar] [CrossRef]
- Barnekow, E.; Liu, W.; Helgadottir, H.T.; Michailidou, K.; Dennis, J.; Bryant, P.; Thutkawkorapin, J.; Wendt, C.; Czene, K.; Hall, P.; et al. A Swedish Genome-Wide Haplotype Association Analysis Identifies a Novel Breast Cancer Susceptibility Locus in 8p21.2 and Characterizes Three Loci on Chromosomes 10, 11 and 16. Cancers 2022, 14, 1206. [Google Scholar] [CrossRef]
- Barnekow, E.; Hasslow, J.; Liu, W.; Bryant, P.; Thutkawkorapin, J.; Wendt, C.; Czene, K.; Hall, P.; Margolin, S.; Lindblom, A. A Swedish Familial Genome-Wide Haplotype Analysis Identified Five Novel Breast Cancer Susceptibility Loci on 9p24.3, 11q22.3, 15q11.2, 16q24.1 and Xq21.31. Int. J. Mol. Sci. 2023, 24, 4468. [Google Scholar] [CrossRef]
- Liu, W.; Mahdessian, H.; Helgadottir, H.; Zhou, X.; Thutkawkorapin, J.; Jiao, X.; Wolk, A.; Lindblom, A. The Swedish Low-risk Colorectal Cancer Study Group Colorectal cancer risk susceptibility loci in a Swedish population. Mol. Carcinog. 2022, 61, 288–300. [Google Scholar] [CrossRef] [PubMed]
- Gabrielson, M.; Eriksson, M.; Hammarström, M.; Borgquist, S.; Leifland, K.; Czene, K.; Hall, P. Cohort Profile: The Karolinska Mammography Project for Risk Prediction of Breast Cancer (KARMA). Int. J. Epidemiol. 2017, 46, 1740–1741g. [Google Scholar] [CrossRef] [PubMed]
- Margolin, S.; Werelius, B.; Fornander, T.; Lindblom, A. BRCA1 Mutations in a Population-based Study of Breast Cancer in Stockholm County. Genet. Test. 2004, 8, 127–132. [Google Scholar] [CrossRef] [PubMed]
- Wendt, C.; Lindblom, A.; Arver, B.; von Wachenfeldt, A.; Margolin, S. Tumour spectrum in non-BRCA hereditary breast cancer families in Sweden. Hered. Cancer Clin. Pract. 2015, 13, 15. [Google Scholar] [CrossRef]
- Illumina. Infinium OncoArray-500K. Available online: http://www.illumina.com/products/by-type/microarray-kits/infinium-oncoarray-500k.html (accessed on 3 November 2024).
- PLINK 1.9. 2018. Retrieved 24 February 2022. Available online: https://www.cog-genomics.org/plink/1.9/ (accessed on 3 November 2024).
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef]
- Available online: http://zzz.bwh.harvard.edu/plink/dist/plink-doc-1.07.pdf (accessed on 3 November 2024).
- Purcell, S. Haplotype-based association tests with GLMs. In PLINK v1.07 Whole Genome Association Analysis Toolset; Cambridge University: Cambridge, UK, 2007; Available online: https://zzz.bwh.harvard.edu/plink/haplo.shtml#hap3 (accessed on 12 December 2024).
- Patel, M.M.; Adrada, B.E. Hereditary Breast Cancer: BRCA mutations and beyond. Radiol. Clin. N. Am. 2024, 62, 627–642. [Google Scholar] [CrossRef]
- Graffeo, R.; Rana, H.; Conforti, F.; Bonanni, B.; Cardoso, M.; Paluch-Shimon, S.; Pagani, O.; Goldhirsch, A.; Partridge, A.; Lambertini, M.; et al. Moderate penetrance genes complicate genetic testing for breast cancer diagnosis: ATM, CHEK2, BARD1 and RAD51D. Breast 2022, 65, 32–40. [Google Scholar] [CrossRef]
- Hatami, M.; Zia, S.; Kanjorpor, A.; Nemati, H.; Sadeghi, M. Impact of alcohol dehydrogenase 3 (ADH3 or ADH1C) genetic variation on head and neck cancer susceptibility: A systematic review, meta-analysis, functional analysis, and trial sequential analysis. Pathol.-Res. Pract. 2024, 262, 155561. [Google Scholar] [CrossRef]
- Zhang, X.; Qiao, G.; Lu, P. Modulation of Fibroblast Growth Factor Signaling Is Essential for Mammary Epithelial Morphogenesis. PLoS ONE 2014, 9, e92735. [Google Scholar] [CrossRef]
- Cui, F.; Wu, D.; Wang, W.; He, X.; Wang, M. Variants of FGFR2 and their associations with breast cancer risk: A HUGE systematic review and meta-analysis. Breast Cancer Res. Treat. 2016, 155, 313–335. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, S.; Kovacs, Z.; Yuan, S.H.; Siszler, G.; Kögl, M.; Summer, H.; Geerts, A.; Golz, S.; Shioda, T.; Methner, A. TOX3 is a neuronal survival factor that induces transcription depending on the presence of CITED1 or phosphorylated CREB in the transcriptionally active complex. J. Cell Sci. 2011, 124, 252–260. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.S.; Mo, J.L.; Hu, L.M.; Gong, C.M.; Liu, T.; Hong, W.X.; Yin, J.Y.; Liu, Z.Q.; Zhou, H.H. Association between CASC16 rs4784227 polymorphism and breast cancer susceptibility: A meta-analysis. Medicine 2021, 100, e26215. [Google Scholar] [CrossRef] [PubMed]
- Easton, D.F.; Pooley, K.A.; Dunning, A.M.; Pharoah, P.D.; Thompson, D.; Ballinger, D.G.; Struewing, J.P.; Morrison, J.; Field, H.; Luben, R.; et al. Genome-wide association study identifies novel breast cancer susceptibility loci. Nature 2007, 447, 1087–1093. [Google Scholar] [CrossRef] [PubMed]
- Gudmundsson, J.; Sulem, P.; Manolescu, A.; Amundadottir, L.T.; Gudbjartsson, D.; Helgason, A.; Rafnar, T.; Bergthorsson, J.T.; A Agnarsson, B.; Baker, A.; et al. Genome-wide association study identifies a second prostate cancer susceptibility variant at 8q24. Nat. Genet. 2007, 39, 631–637. [Google Scholar] [CrossRef]
- Eeles, R.A.; Kote-Jarai, Z.; Giles, G.G.; Al Olama, A.A.; Guy, M.; Jugurnauth, S.K.; Mulholland, S.; Leongamornlert, D.A.; Edwards, S.M.; Morrison, J.; et al. Multiple newly identified loci associated with prostate cancer susceptibility. Nat. Genet. 2008, 40, 316–321. [Google Scholar] [CrossRef]
- Zanke, B.W.; Greenwood, C.M.; Rangrej, J.; Kustra, R.; Tenesa, A.; Farrington, S.M.; Prendergast, J.; Olschwang, S.; Chiang, T.; Crowdy, E.; et al. Genome-wide association scan identifies a colorectal cancer susceptibility locus on chromosome 8q24. Nat. Genet. 2007, 39, 989–994. [Google Scholar] [CrossRef]
- Broderick, P.; Carvajal-Carmona, L.; Pittman, A.M.; Webb, E.; Howarth, K.; Rowan, A.; Lubbe, S.; Spain, S.; Sullivan, K.; Fielding, S.; et al. A genome-wide association study shows that common alleles of SMAD7 influence colorectal cancer risk. Nat. Genet. 2007, 39, 1315–1317. [Google Scholar] [CrossRef]
- Hung, R.J.; McKay, J.D.; Gaborieau, V.; Boffetta, P.; Hashibe, M.; Zaridze, D.; Mukeria, A.; Szeszenia-Dabrowska, N.; Lissowska, J.; Rudnai, P.; et al. A susceptibility locus for lung cancer maps to nicotinic acetylcholine receptor subunit genes on 15q25. Nature 2008, 452, 633–637. [Google Scholar] [CrossRef]
- Brown, K.M.; MacGregor, S.; Montgomery, G.W.; Craig, D.W.; Zhao, Z.Z.; Iyadurai, K.; Henders, A.K.; Homer, N.; Campbell, M.J.; Stark, M.; et al. Common sequence variants on 20q11.22 confer melanoma susceptibility. Nat. Genet. 2008, 40, 838–840. [Google Scholar] [CrossRef]
- Hess, T.; Maj, C.; Gehlen, J.; Borisov, O.; Haas, S.L.; Gockel, I.; Vieth, M.; Piessen, G.; Alakus, H.; Vashist, Y.; et al. Dissecting the genetic heterogeneity of gastric cancer. EBioMedicine 2023, 92, 104616. [Google Scholar] [CrossRef]
- Glubb, D.M.; Thompson, D.J.; Aben, K.K.; Alsulimani, A.; Amant, F.; Annibali, D.; Attia, J.; Barricarte, A.; Beckmann, M.W.; Berchuck, A.; et al. Cross-cancer genome-wide association study of endometrial cancer and epithelial ovarian cancer identifies genetic risk regions associated with risk of both cancers. Cancer Epidemiol. Biomark. Prev. 2021, 30, 217–228. [Google Scholar] [CrossRef]
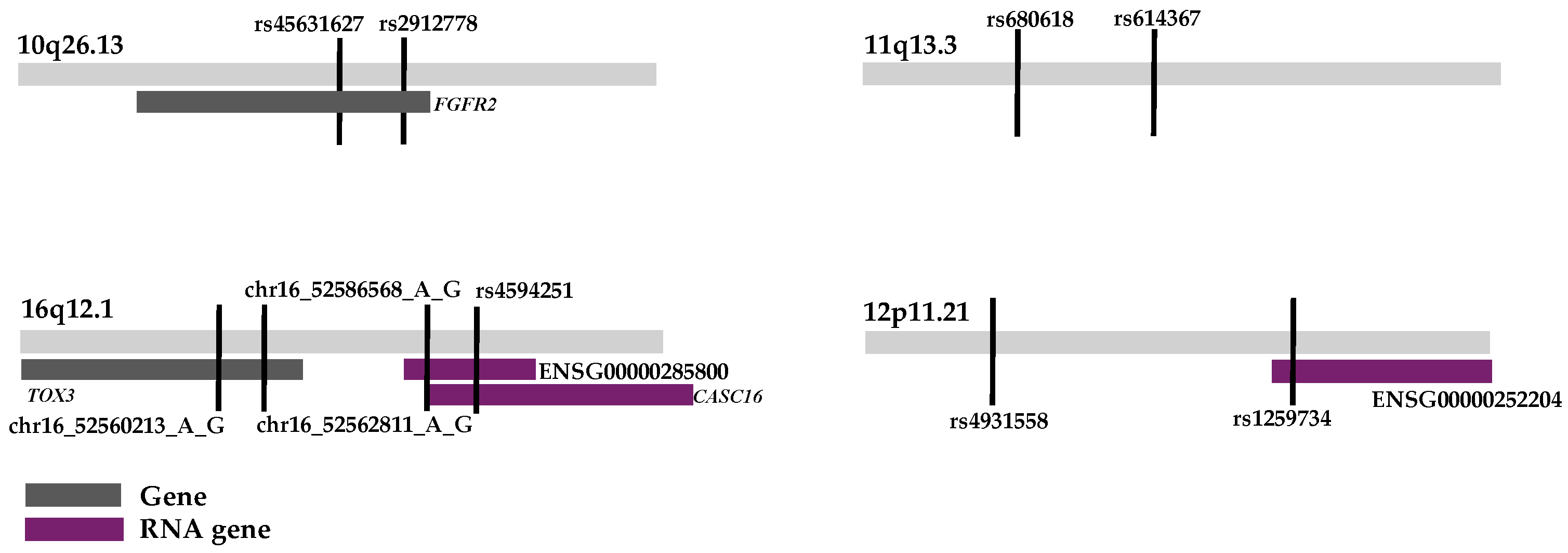

{kind=link}
| Locus | BP1 | BP2 | Haplotype | F | OR | p | Genes |
|---|---|---|---|---|---|---|---|
| 10q26.13 | 121579038 | 121579140 | GG | 0.4 | 1 | 1.96 × 10−15 | FGFR2 |
| 11q13.3 | 69502113 | 69513996 | GGAAAAGGGA | 0.1 | 1 | 2.30 × 10−9 | RNA |
| 12p11.21 | 31804562 | 31877649 | GAGGGGGGAAGCG | 0.1 | 1 | 4.55 × 10−8 | RNA |
| 16q12.1 | 52552656 | 52563932 | GGG | 0.3 | 1 | 1.32 × 10−14 | CASC16 |
| 16q12.1 | 52526301 | 52528899 | AG | 0.3 | 1 | 2.96 × 10−14 | TOX3 |
| Locus | GWAS Sporadic (n = 2550) | GWAS Familial (n = 650) | GWAS All (n = 3200) | |||
|---|---|---|---|---|---|---|
| OR | p-Value | OR | p-Value | OR | p-Value | |
| 10q26.13 | 1.33 | 1.96 × 10−15 | 1.53 ** | 1.70 × 10−12 | 1.36 * | 1.00 × 10−20 |
| 11q13.3 | 1.39 | 2.30 × 10−9 | 1.66 ** | 2.58 × 10−9 | 1.44 * | 6.37 × 10−13 |
| 12p11.21 | 1.42 | 4.55 × 10−8 | 1.08 | 0.524 | 1.35 | 7.92 × 10−7 |
| 16q12.1 | 1.34 | 1.32 × 10−14 | 1.45 ** | 4.39 × 10−9 | 1.37 * | 2.38 × 10−18 |
| 16q12.1 | 1.34 | 2.96 × 10−14 | 1.49 ** | 3.97E × 10−10 | 1.37 * | 1.71 × 10−18 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vermani, L.; Barnekow, E.; Liu, W.; Wendt, C.; Hall, P.; Margolin, S.; Lindblom, A. Swedish Genome-Wide Haplotype Association Analysis Suggests Breast Cancer Loci with Varying Risk-Modifying Effects. Genes 2024, 15, 1616. https://doi.org/10.3390/genes15121616
Vermani L, Barnekow E, Liu W, Wendt C, Hall P, Margolin S, Lindblom A. Swedish Genome-Wide Haplotype Association Analysis Suggests Breast Cancer Loci with Varying Risk-Modifying Effects. Genes. 2024; 15(12):1616. https://doi.org/10.3390/genes15121616
Chicago/Turabian StyleVermani, Litika, Elin Barnekow, Wen Liu, Camilla Wendt, Per Hall, Sara Margolin, and Annika Lindblom. 2024. "Swedish Genome-Wide Haplotype Association Analysis Suggests Breast Cancer Loci with Varying Risk-Modifying Effects" Genes 15, no. 12: 1616. https://doi.org/10.3390/genes15121616
APA StyleVermani, L., Barnekow, E., Liu, W., Wendt, C., Hall, P., Margolin, S., & Lindblom, A. (2024). Swedish Genome-Wide Haplotype Association Analysis Suggests Breast Cancer Loci with Varying Risk-Modifying Effects. Genes, 15(12), 1616. https://doi.org/10.3390/genes15121616