
Gene Expression Analysis of nc-RNAs in Bipolar and Panic Disorders: A Pilot Study




Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Healthy Controls
2.2. Therapy and Second Sampling
2.3. RNA Extraction and cDNA Synthesis
2.4. Real Time Polymerase Chain Reaction (rt-PCR) Analysis
2.5. Statistical Analysis
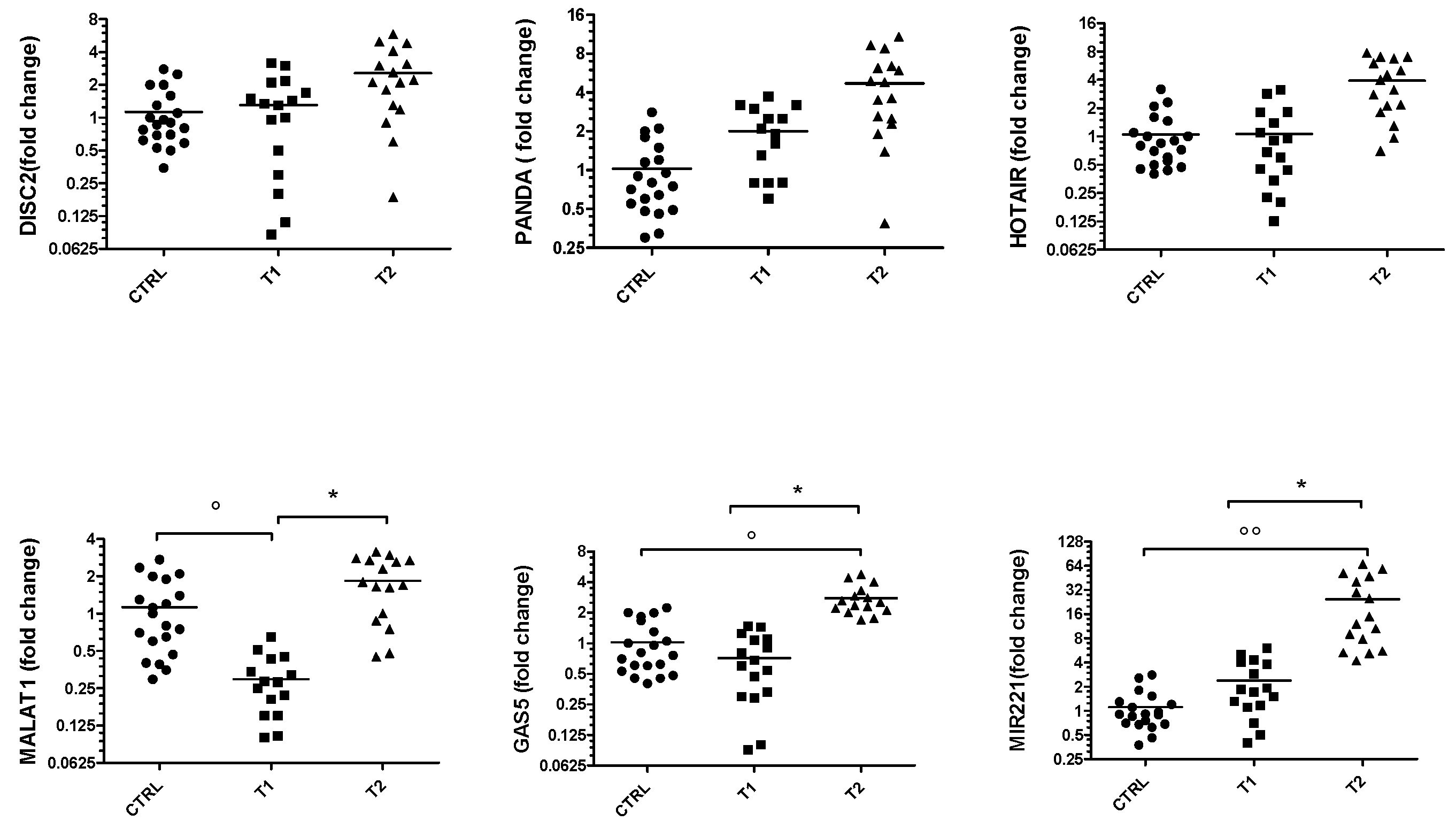
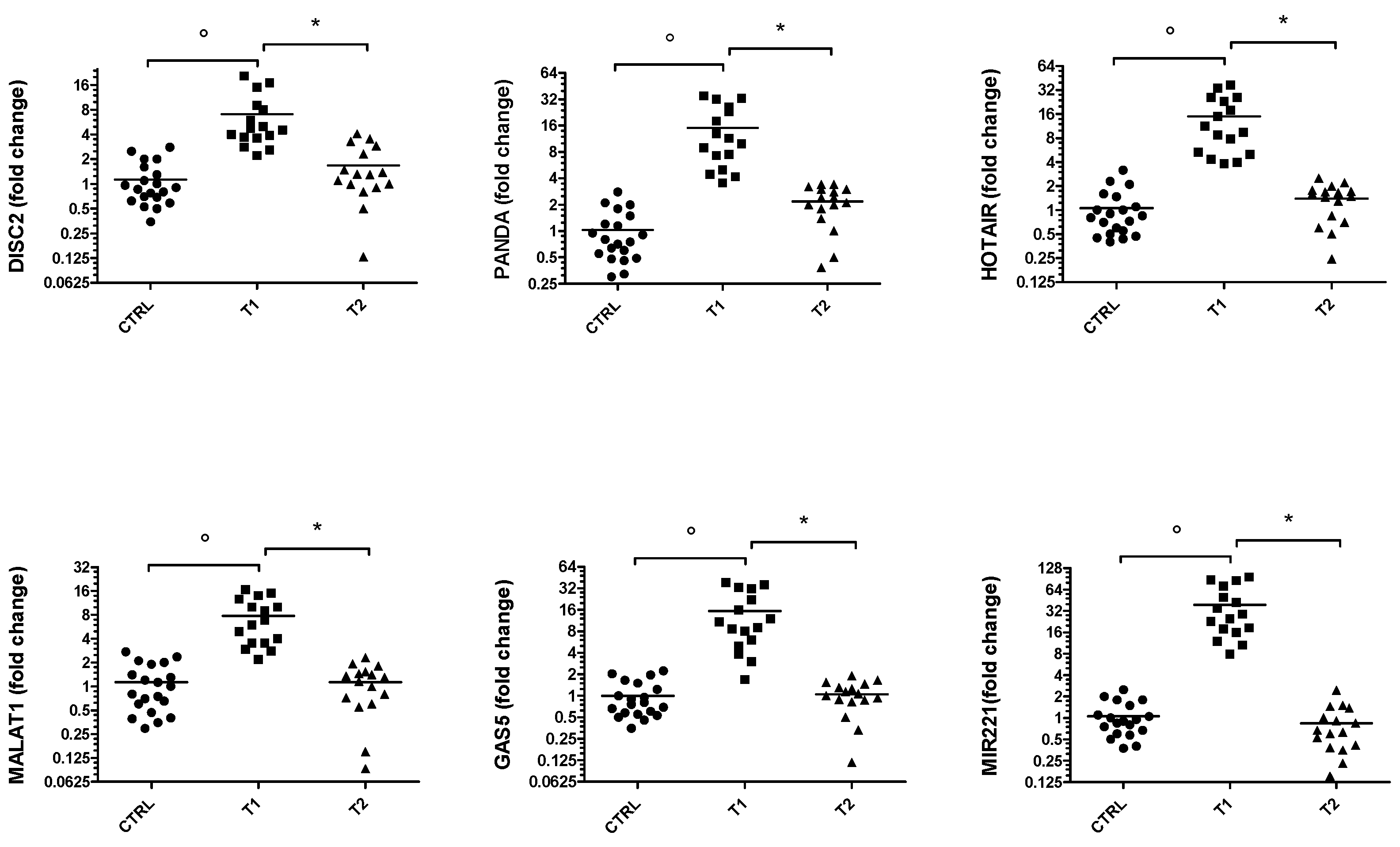
3. Results
3.1. ncRNAs Expression in BD
3.2. ncRNAs Expression in PD
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, S.; Mao, C.; Liu, S. Peptides encoded by noncoding genes: Challenges and perspectives. Signal Transduct. Target. Ther. 2019, 4, 57. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Esteller, M. Non-coding RNAs in human disease. Nat. Rev. Genet. 2011, 12, 861–874. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Veneziano, D.; Acunzo, M.; Croce, C.M. Small non-coding RNA and cancer. Carcinogenesis 2017, 38, 485–491. [Google Scholar] [PubMed]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and Functions of Long Noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [PubMed]
- Hadjicharalambous, M.R.; Lindsay, M.A. Long non-coding RNAs and the innate immune response. Non-Coding RNA 2019, 5, 34. [Google Scholar] [CrossRef] [PubMed]
- Yoshino, Y.; Dwivedi, Y. Non-Coding RNAs in Psychiatric Disorders and Suicidal Behavior. Front. Psychiatry 2020, 11, 543893. [Google Scholar] [CrossRef] [PubMed]
- Vieta, E.; Berk, M.; Schulze, T.G.; Carvalho, A.F.; Suppes, T.; Calabrese, J.R.; Gao, K.; Miskowiak, K.W.; Grande, I. Bipolar disorders. Nat. Rev. Dis. Prim. 2018, 4, 18008. [Google Scholar] [CrossRef] [PubMed]
- Maina, G.; Rosso, G.; Aguglia, A.; Chiodelli, D.F.; Bogetto, F. Anxiety and bipolar disorders: Epidemiological and clinical aspects. Ital. J. Psychopathol. 2011, 17, 365–375. [Google Scholar]
- Marabita, F.; De Candia, P.; Torri, A.; Tegner, J.; Abrignani, S.; Rossi, R.L. Normalization of circulating microRNA expression data obtained by quantitative real-time RT-PCR. Brief. Bioinform. 2016, 17, 204–212. [Google Scholar] [CrossRef] [PubMed]
- Iempridee, T.; Wiwithaphon, S.; Piboonprai, K.; Pratedrat, P.; Khumkhrong, P.; Japrung, D.; Temisak, S.; Laiwejpithaya, S.; Chaopotong, P.; Dharakul, T. Identification of reference genes for circulating long noncoding RNA analysis in serum of cervical cancer patients. FEBS Open Bio 2018, 8, 1844–1854. [Google Scholar] [CrossRef] [PubMed]
- Shirvani Farsani, Z.; Zahirodin, A.; Ghaderian SM, H.; Shams, J.; Naghavi Gargari, B. The role of long non-coding RNA MALAT1 in patients with bipolar disorder. Metab. Brain Dis. 2020, 35, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Sudhalkar, N.; Rosen, C.; Melbourne, J.K.; Park, M.R.; Chase, K.A.; Sharma, R.P. Long non-coding RNAs associated with heterochromatin function in immune cells in psychosis. Non-Coding RNA 2018, 4, 43. [Google Scholar] [CrossRef] [PubMed]
- Kino, T.; Hurt, D.E.; Ichijo, T.; Nader, N.; Chrousos, G.P. Noncoding RNA Gas5 is a growth arrest- and starvation-associated repressor of the glucocorticoid receptor. Sci. Signal. 2010, 3, ra8. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Watson, S.; Young, A.H. Glucocorticoid receptor dysfunction: Consequences for the pathophysiology and treatment of mood disorders. Indian J. Psychiatry 2003, 45, 5. [Google Scholar] [PubMed]
- Chen, H.; Wang, N.; Burmeister, M.; McInnis, M.G. MicroRNA expression changes in lymphoblastoid cell lines in response to lithium treatment. Int. J. Neuropsychopharmacol. 2009, 12, 975–981. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
| Gene | Sequences |
|---|---|
| GAS5 | F:CATTGGCACACAGGCATTAG R:TACCCAAGCAAGTCATCCATG |
| DISC2 | F:CAGCCTCCCAAGTAGCTAGGAT R:CTGTAATCCCAGCACTTTGGAA |
| PANDA | F: GCCTGTTCCTCAATCCAAGA R: TTGCTTCTGGGCAGAACTTG |
| MALAT1 | F: GGAAAGCGAGTGGTTGGTAA R:ATCCCTTTACACCTCAGTACGA |
| HOTAIR | F: GCACTCACAGACAGAGGTTTA R:CTCTGTACTCCCGTTCCCTAGA |
| GADPH | F: CACCAGGGCTGCTTTTAACTCT R: ATCTCGCTCCTGGAAGATGGT |
| miR-221-5p | F: TGAACATCCAGGTCTGGG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bella, F.; Muscatello, M.R.A.; D’Ascola, A.; Campo, S. Gene Expression Analysis of nc-RNAs in Bipolar and Panic Disorders: A Pilot Study. Genes 2023, 14, 1778. https://doi.org/10.3390/genes14091778
Bella F, Muscatello MRA, D’Ascola A, Campo S. Gene Expression Analysis of nc-RNAs in Bipolar and Panic Disorders: A Pilot Study. Genes. 2023; 14(9):1778. https://doi.org/10.3390/genes14091778
Chicago/Turabian StyleBella, Fabrizio, Maria Rosaria Anna Muscatello, Angela D’Ascola, and Salvatore Campo. 2023. "Gene Expression Analysis of nc-RNAs in Bipolar and Panic Disorders: A Pilot Study" Genes 14, no. 9: 1778. https://doi.org/10.3390/genes14091778
APA StyleBella, F., Muscatello, M. R. A., D’Ascola, A., & Campo, S. (2023). Gene Expression Analysis of nc-RNAs in Bipolar and Panic Disorders: A Pilot Study. Genes, 14(9), 1778. https://doi.org/10.3390/genes14091778