

Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases



Abstract
1. Introduction
2. Materials and Methods
2.1. Classification Criteria of the Disease
2.2. Appraisal of the Disease-Associated Sex-Biased miRNAs
2.3. Analysis of Evolutionary Conservation of the Disease-Associated Sex-Biased miRNAs
2.4. Analysis of TFs and the Function of the Disease-Associated Sex-Biased miRNAs
3. Results
3.1. Subsection
3.1.1. Identification of the Disease-Associated Sex-Biased miRNAs
3.1.2. Analysis of the Chromosomal Arrangement of the Disease-Associated Sex-Biased miRNA Genes
3.1.3. Analysis of the Evolutionary Conservation of the Disease-Associated Sex-Biased miRNA Genes
3.1.4. Analysis of Tissue Specificity of the Disease-Associated Sex-Biased miRNAs
3.1.5. Analysis of Transcription Factors of the Disease-Associated Sex-Biased miRNAs
3.1.6. Analysis of the Disease Spectrum Width of Disease-Associated Sex-Biased miRNAs
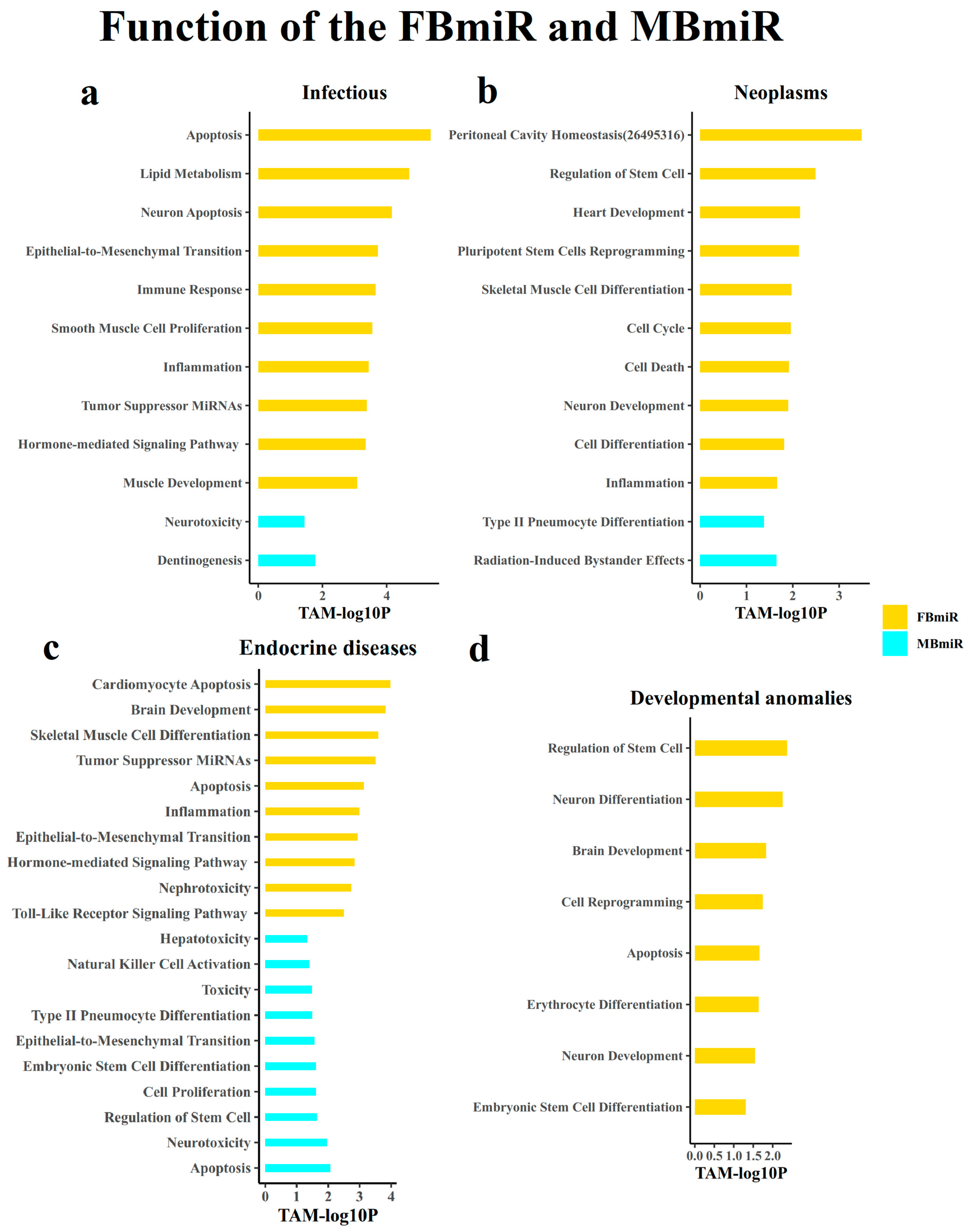
3.1.7. Analysis of the Function of the Disease-Associated Sex-Biased miRNAs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engelhardt, M.; Brioli, A.; von Lilienfeld-Toal, M. Differences due to socio-economic status, genetic background and sex in cancer and precision medicine—An intersectional approach to close the care gap for marginalized groups. Dtsch. Med. Wochenschr. 2023, 148, 528–538. [Google Scholar] [CrossRef] [PubMed]
- Bancks, M.P.; Akhabue, E.; Rana, J.S.; Reis, J.P.; Schreiner, P.J.; Yano, Y.; Lewis, C.E. Sex differences in cardiovascular risk factors before and after the development of type 2 diabetes and risk for incident cardiovascular disease. Diabetes Res. Clin. Pract. 2020, 166, 108334. [Google Scholar] [CrossRef] [PubMed]
- Tersalvi, G.; Gaiero, L.; Capriolo, M.; Cristoforetti, Y.; Salizzoni, S.; Senatore, G.; Pedrazzini, G.; Biasco, L. Sex Differences in Epidemiology, Morphology, Mechanisms, and Treatment of Mitral Valve Regurgitation. Medicina 2023, 59, 1017. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.; Qi, Y.; Ding, L.; Ding, S.; Han, Z.; Wang, Y.; Du, P. miRNA dosage control in development and human disease. Trends Cell Biol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Cui, C.; Yang, W.; Shi, J.; Zhou, Y.; Yang, J.; Cui, Q.; Zhou, Y. Identification and Analysis of Human Sex-biased MicroRNAs. Genom. Proteom. Bioinform. 2018, 16, 200–211. [Google Scholar] [CrossRef]
- Deny, M.; Popotas, A.; Hanssens, L.; Lefevre, N.; Arroba Nunez, L.A.; Ouafo, G.S.; Corazza, F.; Casimir, G.; Chamekh, M. Sex-biased expression of selected chromosome x-linked microRNAs with potent regulatory effect on the inflammatory response in children with cystic fibrosis: A preliminary pilot investigation. Front. Immunol. 2023, 14, 1114239. [Google Scholar] [CrossRef]
- Gunay, N.; Taheri, S.; Memis, M.; Yilmaz Sukranli, Z.; Sahin, T.; Demiryurek, S.; Ekici Gunay, N.; Aslan, Y.E.; Demiryurek, A.T. Male- and female-specific microRNA expression patterns in a mouse model of methanol poisoning. Food Chem. Toxicol. 2023, 174, 113666. [Google Scholar] [CrossRef]
- Kahnamoui, S.; Basu, S.; Lei, Y.; Patel, D.; Keijzer, R.; Pascoe, C.D. MicroRNA-200b deficiency is not sufficient to increase susceptibility to allergen induced airway inflammation and dysfunction in mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2023, 325, L45–L53. [Google Scholar] [CrossRef]
- Rosenberg, L.; Liu, C.; Sharma, R.; Wood, C.; Vyhlidal, C.A.; Gaedigk, R.; Kho, A.T.; Ziniti, J.P.; Celedon, J.C.; Tantisira, K.G.; et al. Intrauterine Smoke Exposure, microRNA Expression during Human Lung Development, and Childhood Asthma. Int. J. Mol. Sci. 2023, 24, 7727. [Google Scholar] [CrossRef]
- Blatkiewicz, M.; Sielatycka, K.; Piotrowska, K.; Kilanczyk, E. DHEA and Its Metabolites Reduce the Cytokines Involved in the Inflammatory Response and Fibrosis in Primary Biliary Cholangitis. Int. J. Mol. Sci. 2023, 24, 5301. [Google Scholar] [CrossRef]
- Zhou, C.; Freel, C.; Mills, O.; Yang, X.R.; Yan, Q.; Zheng, J. MicroRNA-29 Differentially Mediates Preeclampsia-Dysregulated Cellular Responses to Cytokines in Female and Male Fetal Endothelial Cells. bioRxiv 2023. [Google Scholar] [CrossRef] [PubMed]
- Tomeva, E.; Krammer, U.D.B.; Switzeny, O.J.; Haslberger, A.G.; Hippe, B. Sex-Specific miRNA Differences in Liquid Biopsies from Subjects with Solid Tumors and Healthy Controls. Epigenomes 2023, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Florijn, B.W.; Bijkerk, R.; Kruyt, N.D.; van Zonneveld, A.J.; Wermer, M.J.H. Sex-Specific MicroRNAs in Neurovascular Units in Ischemic Stroke. Int. J. Mol. Sci. 2021, 22, 11888. [Google Scholar] [CrossRef] [PubMed]
- Florijn, B.W.; Valstar, G.B.; Duijs, J.; Menken, R.; Cramer, M.J.; Teske, A.J.; Ghossein-Doha, C.; Rutten, F.H.; Spaanderman, M.E.A.; den Ruijter, H.M.; et al. Sex-specific microRNAs in women with diabetes and left ventricular diastolic dysfunction or HFpEF associate with microvascular injury. Sci. Rep. 2020, 10, 13945. [Google Scholar] [CrossRef]
- Guo, L.; Zhang, Q.; Ma, X.; Wang, J.; Liang, T. miRNA and mRNA expression analysis reveals potential sex-biased miRNA expression. Sci. Rep. 2017, 7, 39812. [Google Scholar] [CrossRef]
- Queiros, A.M.; Eschen, C.; Fliegner, D.; Kararigas, G.; Dworatzek, E.; Westphal, C.; Sanchez Ruderisch, H.; Regitz-Zagrosek, V. Sex- and estrogen-dependent regulation of a miRNA network in the healthy and hypertrophied heart. Int. J. Cardiol. 2013, 169, 331–338. [Google Scholar] [CrossRef]
- Meder, B.; Backes, C.; Haas, J.; Leidinger, P.; Stähler, C.; Großmann, T.; Vogel, B.; Frese, K.; Giannitsis, E.; Katus, H.A.; et al. Influence of the confounding factors age and sex on microRNA profiles from peripheral blood. Clin. Chem. 2014, 60, 1200–1208. [Google Scholar] [CrossRef]
- Rounge, T.B.; Umu, S.U.; Keller, A.; Meese, E.; Ursin, G.; Tretli, S.; Lyle, R.; Langseth, H. Circulating small non-coding RNAs associated with age, sex, smoking, body mass and physical activity. Sci. Rep. 2018, 8, 17650. [Google Scholar] [CrossRef]
- Sosa, E.; Flores, L.; Yan, W.; McCarrey, J.R. Escape of X-linked miRNA genes from meiotic sex chromosome inactivation. Development 2015, 142, 3791–3800. [Google Scholar] [CrossRef]
- Ullah Jan, P.F.; Kousar, S.; Mahmood, A.; Nadeem, S.; Malik, K.; Safir, W.; Shujaat, N.; Khan, F.U.; Shahid, M. Risk Loci For Chronic Obstructive Disease Reside On Chromosome 14: A Case-Control Study On The Pakistani Population. J. Ayub Med. Coll. Abbottabad 2023, 35, 203–209. [Google Scholar] [CrossRef]
- Gonzalez-Espinoza, A.; Zamora-Fuentes, J.; Hernandez-Lemus, E.; Espinal-Enriquez, J. Gene Co-Expression in Breast Cancer: A Matter of Distance. Front. Oncol. 2021, 11, 726493. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Qin, F.; Lai, X.; Yang, T.; Yu, J.; Wei, C.; Wei, L.; Li, J. Exploring heterogeneity of tumor immune cells and adrenal cells in aldosterone-producing adenomas using single-cell RNA-seq and investigating differences by sex. Heliyon 2023, 9, e14357. [Google Scholar] [CrossRef] [PubMed]
- Conesa, A.; Gotz, S.; Garcia-Gomez, J.M.; Terol, J.; Talon, M.; Robles, M. Blast2GO: A universal tool for annotation, visualization and analysis in functional genomics research. Bioinformatics 2005, 21, 3674–3676. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, Q.; Deng, M.; Miao, J.; Guo, Y.; Gao, W.; Cui, Q. An analysis of human microRNA and disease associations. PLoS ONE 2008, 3, e3420. [Google Scholar] [CrossRef]
- Liang, Y.; Ridzon, D.; Wong, L.; Chen, C. Characterization of microRNA expression profiles in normal human tissues. BMC Genom. 2007, 8, 166. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Xia, L.; Zhang, L.; Zhao, K.; Li, C. MiRNA-144-5p down-modulates CDCA3 to regulate proliferation and apoptosis of lung adenocarcinoma cells. Mutat. Res. 2022, 825, 111798. [Google Scholar] [CrossRef]
- Shi, Y.; Massague, J. Mechanisms of TGF-β signaling from cell membrane to the nucleus. Cell 2003, 113, 685–700. [Google Scholar] [CrossRef]
- Poppelaars, F.; Gaya da Costa, M.; Faria, B.; Eskandari, S.K.; Damman, J.; Seelen, M.A. A functional TGFB1 polymorphism in the donor associates with long-term graft survival after kidney transplantation. Clin. Kidney J. 2022, 15, 278–286. [Google Scholar] [CrossRef]
- Mosly, D.; MacLeod, K.; Moir, N.; Turnbull, A.; Sims, A.H.; Langdon, S.P. Variation in IL6ST cytokine family function and the potential of IL6 trans-signalling in ERalpha positive breast cancer cells. Cell Signal 2023, 103, 110563. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tissue | Disease | Dataset ID | Number of Samples | Number of miRNAs | ||
|---|---|---|---|---|---|---|
| Female | Male | FBmiR | MBmiR | |||
| Peripheral blood | pulmonary tuberculosis and sarcoidosis | GSE34608 | 28 | 54 | 26 | 5 |
| whole blood leukocytes | GSE134358 | 167 | 308 | 136 | 191 | |
| lung cancer | GSE137140 | 749 | 997 | 1189 | 216 | |
| cytogenetically normal acute myeloid leukemia | GSE142699 | 11 | 13 | 4 | 0 | |
| Dementia | GSE167559 | 41 | 43 | 0 | 15 | |
| transposition of the great arteries and systemic left and right ventricles | GSE215940 | 9 | 22 | 3 | 0 | |
| Brain | surgical specimens of primary glioblastoma multiform | GSE25631 | 28 | 54 | 26 | 5 |
| pediatric brain tumors | GSE42657 | 24 | 33 | 16 | 3 | |
| glioma-related epilepsy | GSE199759 | 1 | 8 | 0 | 0 | |
| temporal lobe epilepsy with hippocampal sclerosis | GSE205661 | 3 | 3 | 0 | 0 | |
| Neoplasms | human osteosarcoma (biopsies) | GSE39040 | 35 | 30 | 11 | 6 |
| human osteosarcoma | GSE39052 | 15 | 11 | 16 | 15 | |
| locally advanced rectal cancer (LARC) | GSE68204 | 12 | 25 | 0 | 0 | |
| HB tumors | GSE75283 | 31 | 27 | 9 | 4 | |
| hepatocellular carcinoma with portal vein tumor thrombosis | GSE76903 | 9 | 51 | 88 | 17 | |
| non-small cell lung cancer | GSE102286 | 38 | 52 | 5 | 0 | |
| squamous cell carcinoma | GSE124678 | 9 | 23 | 7 | 0 | |
| colorectal cancer | GSE126093 | 4 | 6 | 3 | 1 | |
| Small Cell Bladder Cancer | GSE145259 | 3 | 19 | 0 | 0 | |
| Stage I and II Clear Cell Renal Cell Carcinomas | GSE155209 | 2 | 18 | 0 | 0 | |
| Other | insulin resistance in primary human adipocytes | GSE41223 | 28 | 54 | 26 | 5 |
| congenital aniridia | GSE137995 | 15 | 5 | 55 | 25 | |
| diabetic retinopathy | GSE160308 | 29 | 27 | 9 | 0 | |
| healing and non-healing wounds | GSE174661 | 6 | 4 | 7 | 0 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, B.; Cui, C.; Cui, Q. Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases. Genes 2023, 14, 1688. https://doi.org/10.3390/genes14091688
Zhong B, Cui C, Cui Q. Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases. Genes. 2023; 14(9):1688. https://doi.org/10.3390/genes14091688
Chicago/Turabian StyleZhong, Bitao, Chunmei Cui, and Qinghua Cui. 2023. "Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases" Genes 14, no. 9: 1688. https://doi.org/10.3390/genes14091688
APA StyleZhong, B., Cui, C., & Cui, Q. (2023). Identification and Analysis of Sex-Biased MicroRNAs in Human Diseases. Genes, 14(9), 1688. https://doi.org/10.3390/genes14091688