


Y-Chromosome Haplotype Report among Eight Italian Horse Breeds

 , , ,
, , , {kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
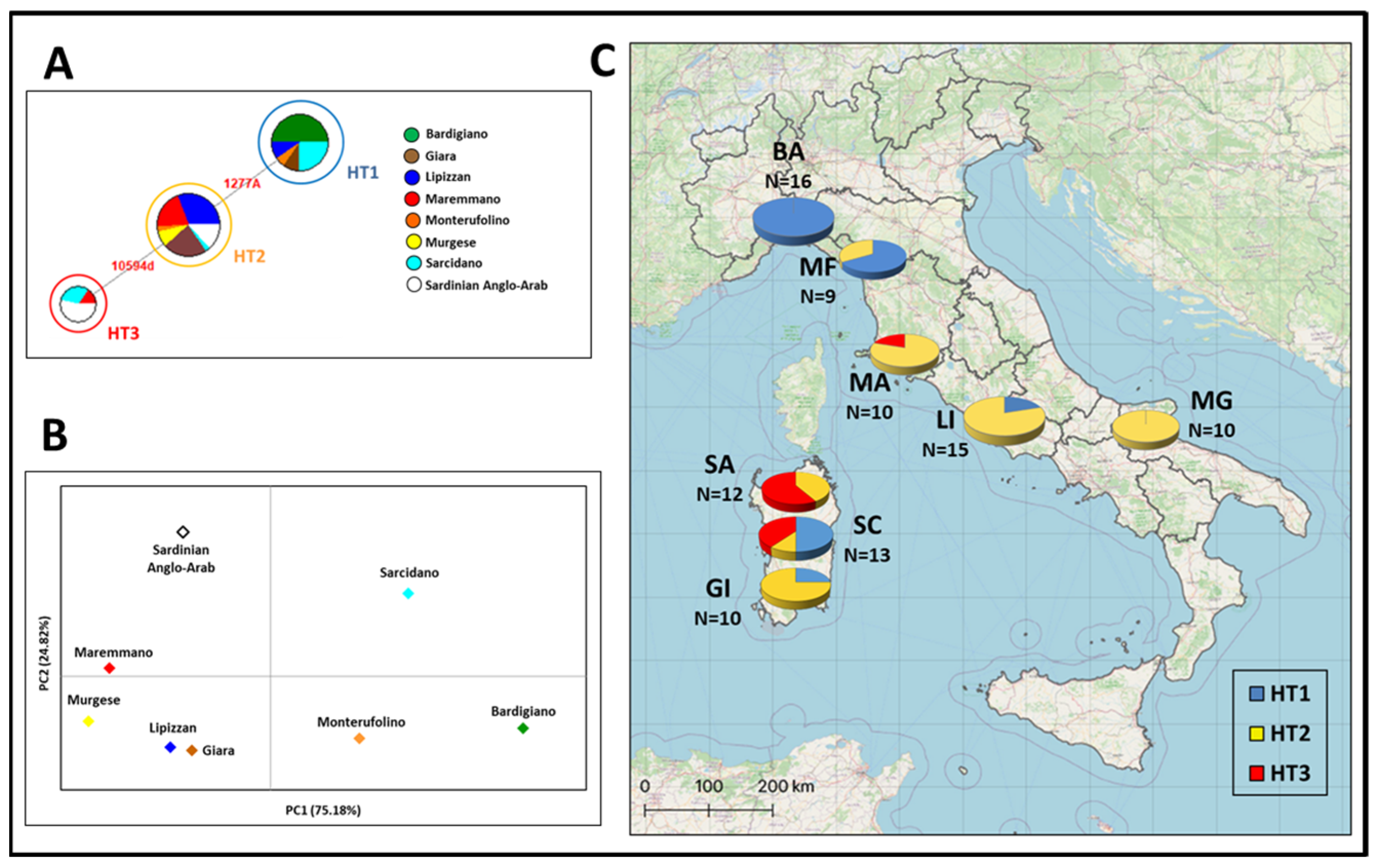
3.1. Y-Chromosome Haplotypes in the Italian Breeds
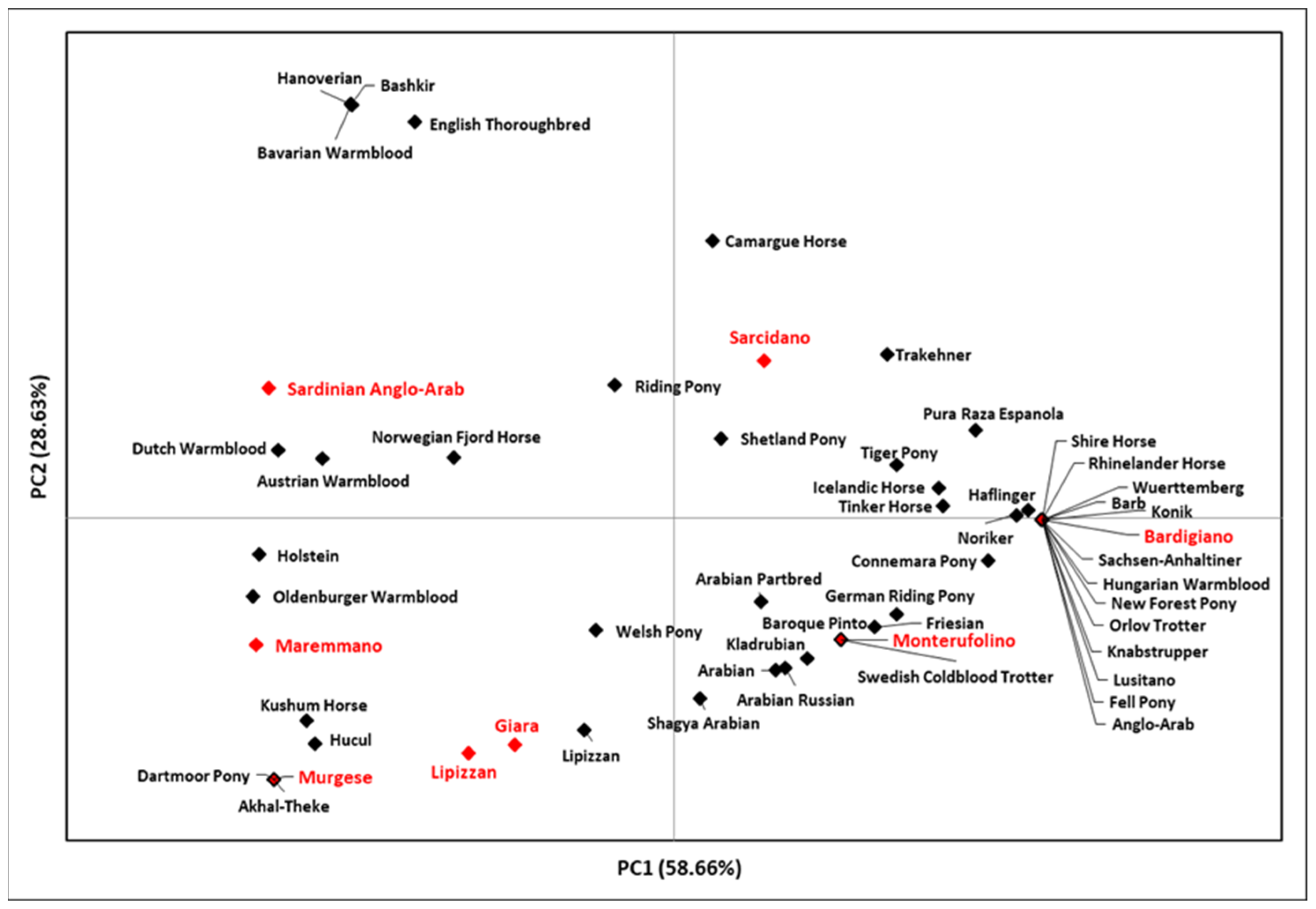
3.2. The Italian Y-Chromosome Haplotypes in the Eurasian and Mediterranean Contexts
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Niemi, M.; Bläuer, A.; Iso-Touru, T.; Nyström, V.; Harjula, J.; Taavitsainen, J.-P.; Storå, J.; Lidén, K.; Kantanen, J. Mitochondrial DNA and Y-Chromosomal Diversity in Ancient Populations of Domestic Sheep (Ovis aries) in Finland: Comparison with Contemporary Sheep Breeds. Genet. Sel. Evol. 2013, 45, 2. [Google Scholar] [CrossRef] [PubMed]
- Ajmone-Marsan, P.; Colli, L.; Han, J.L.; Achilli, A.; Lancioni, H.; Joost, S.; Crepaldi, P.; Pilla, F.; Stella, A.; Taberlet, P.; et al. The Characterization of Goat Genetic Diversity: Towards a Genomic Approach. Small Rumin. Res. 2014, 121, 58–72. [Google Scholar] [CrossRef]
- Machová, K.; Málková, A.; Vostrý, L. Sheep Post-Domestication Expansion in the Context of Mitochondrial and Y Chromosome Haplogroups and Haplotypes. Genes 2022, 13, 613. [Google Scholar] [CrossRef] [PubMed]
- VarGoats Consortium; Nijman, I.J.; Rosen, B.D.; Bardou, P.; Faraut, T.; Cumer, T.; Daly, K.G.; Zheng, Z.; Cai, Y.; Asadollahpour, H.; et al. Geographical Contrasts of Y-chromosomal Haplogroups from Wild and Domestic Goats Reveal Ancient Migrations and Recent Introgressions. Mol. Ecol. 2022, 31, 4364–4380. [Google Scholar] [CrossRef] [PubMed]
- Di Lorenzo, P.; Lancioni, H.; Ceccobelli, S.; Curcio, L.; Panella, F.; Lasagna, E. Uniparental Genetic Systems: A Male and a Female Perspective in the Domestic Cattle Origin and Evolution. Electron. J. Biotechnol. 2016, 23, 69–78. [Google Scholar] [CrossRef]
- Di Lorenzo, P.; Lancioni, H.; Ceccobelli, S.; Colli, L.; Cardinali, I.; Karsli, T.; Capodiferro, M.R.; Sahin, E.; Ferretti, L.; Ajmone Marsan, P.; et al. Mitochondrial DNA Variants of Podolian Cattle Breeds Testify for a Dual Maternal Origin. PLoS ONE 2018, 13, e0192567. [Google Scholar] [CrossRef]
- Lancioni, H.; Di Lorenzo, P.; Cardinali, I.; Ceccobelli, S.; Capodiferro, M.R.; Fichera, A.; Grugni, V.; Semino, O.; Ferretti, L.; Gruppetta, A.; et al. Survey of Uniparental Genetic Markers in the Maltese Cattle Breed Reveals a Significant Founder Effect but Does Not Indicate Local Domestication. Anim. Genet. 2016, 47, 267–269. [Google Scholar] [CrossRef]
- Mauki, D.H.; Adeola, A.C.; Ng’ang’a, S.I.; Tijjani, A.; Akanbi, I.M.; Sanke, O.J.; Abdussamad, A.M.; Olaogun, S.C.; Ibrahim, J.; Dawuda, P.M.; et al. Genetic Variation of Nigerian Cattle Inferred from Maternal and Paternal Genetic Markers. PeerJ 2021, 9, e10607. [Google Scholar] [CrossRef]
- Wallner, B.; Brem, G.; Müller, M.; Achmann, R. Fixed Nucleotide Differences on the Y Chromosome Indicate Clear Divergence between Equus przewalskii and Equus caballus: Fixed Nucleotide Difference on the Y Chromosome. Anim. Genet. 2003, 34, 453–456. [Google Scholar] [CrossRef]
- Wallner, B. Isolation of Y Chromosome-Specific Microsatellites in the Horse and Cross-Species Amplification in the Genus Equus. J. Hered. 2004, 95, 158–164. [Google Scholar] [CrossRef]
- Ling, Y.; Ma, Y.; Guan, W.; Cheng, Y.; Wang, Y.; Han, J.; Jin, D.; Mang, L.; Mahmut, H. Identification of Y Chromosome Genetic Variations in Chinese Indigenous Horse Breeds. J. Hered. 2010, 101, 639–643. [Google Scholar] [CrossRef] [PubMed]
- Wallner, B.; Vogl, C.; Shukla, P.; Burgstaller, J.P.; Druml, T.; Brem, G. Identification of Genetic Variation on the Horse Y Chromosome and the Tracing of Male Founder Lineages in Modern Breeds. PLoS ONE 2013, 8, e60015. [Google Scholar] [CrossRef] [PubMed]
- Brandariz-Fontes, C.; Leonard, J.A.; Vega-Pla, J.L.; Backström, N.; Lindgren, G.; Lippold, S.; Rico, C. Y-Chromosome Analysis in Retuertas Horses. PLoS ONE 2013, 8, e64985. [Google Scholar] [CrossRef] [PubMed]
- Kreutzmann, N.; Brem, G.; Wallner, B. The Domestic Horse Harbours Y-Chromosomal Microsatellite Polymorphism Only on Two Widely Distributed Male Lineages. Anim. Genet. 2014, 45, 460. [Google Scholar] [CrossRef]
- Han, H.; Zhang, Q.; Gao, K.; Yue, X.; Zhang, T.; Dang, R.; Lan, X.; Chen, H.; Lei, C. Y-Single Nucleotide Polymorphisms Diversity in Chinese Indigenous Horse. Asian-Aust. J. Anim. Sci. 2015, 28, 1066–1074. [Google Scholar] [CrossRef]
- Wallner, B.; Palmieri, N.; Vogl, C.; Rigler, D.; Bozlak, E.; Druml, T.; Jagannathan, V.; Leeb, T.; Fries, R.; Tetens, J.; et al. Y Chromosome Uncovers the Recent Oriental Origin of Modern Stallions. Curr. Biol. 2017, 27, 2029–2035. [Google Scholar] [CrossRef]
- Felkel, S.; Vogl, C.; Rigler, D.; Jagannathan, V.; Leeb, T.; Fries, R.; Neuditschko, M.; Rieder, S.; Velie, B.; Lindgren, G.; et al. Asian Horses Deepen the MSY Phylogeny. Anim. Genet. 2018, 49, 90–93. [Google Scholar] [CrossRef]
- Janečka, J.E.; Davis, B.W.; Ghosh, S.; Paria, N.; Das, P.J.; Orlando, L.; Schubert, M.; Nielsen, M.K.; Stout, T.A.E.; Brashear, W.; et al. Horse Y Chromosome Assembly Displays Unique Evolutionary Features and Putative Stallion Fertility Genes. Nat. Commun. 2018, 9, 2945. [Google Scholar] [CrossRef]
- Kakoi, H.; Kikuchi, M.; Tozaki, T.; Hirota, K.I.; Nagata, S.I.; Hobo, S.; Takasu, M. Distribution of Y Chromosomal Haplotypes in Japanese Native Horse Populations. J. Equine Sci. 2018, 29, 39–42. [Google Scholar] [CrossRef]
- Felkel, S.; Vogl, C.; Rigler, D.; Dobretsberger, V.; Chowdhary, B.P.; Distl, O.; Fries, R.; Jagannathan, V.; Janečka, J.E.; Leeb, T.; et al. The Horse Y Chromosome as an Informative Marker for Tracing Sire Lines. Sci. Rep. 2019, 9, 6095. [Google Scholar] [CrossRef]
- Han, H.; Wallner, B.; Rigler, D.; MacHugh, D.E.; Manglai, D.; Hill, E.W. Chinese Mongolian Horses May Retain Early Domestic Male Genetic Lineages yet to Be Discovered. Anim. Genet. 2019, 50, 399–402. [Google Scholar] [CrossRef] [PubMed]
- Sild, E.; Värv, S.; Kaart, T.; Kantanen, J.; Popov, R.; Viinalass, H. Maternal and Paternal Genetic Variation in Estonian Local Horse Breeds in the Context of Geographically Adjacent and Distant Eurasian Breeds. Anim. Genet. 2019, 50, 757–760. [Google Scholar] [CrossRef]
- Giontella, A.; Cardinali, I.; Pieramati, C.; Cherchi, R.; Biggio, G.P.; Achilli, A.; Silvestrelli, M.; Lancioni, H. A Genetic Window on Sardinian Native Horse Breeds through Uniparental Molecular Systems. Animals 2020, 10, 1544. [Google Scholar] [CrossRef] [PubMed]
- Giontella, A.; Cardinali, I.; Lancioni, H.; Giovannini, S.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Mitochondrial DNA Survey Reveals the Lack of Accuracy in Maremmano Horse Studbook Records. Animals 2020, 10, 839. [Google Scholar] [CrossRef] [PubMed]
- Lancioni, H.; Cardinali, I.; Giontella, A.; Antognoni, M.T.; Miglio, A. Mitochondrial DNA Variation in the Italian Heavy Draught Horse. PeerJ 2020, 8, e8996. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Yang, Y.; Pan, Q.; Sun, Y.; Ma, H.; Liu, Y.; Wang, M.; Zhao, C.; Wu, C. Ancient Patrilineal Lines and Relatively High ECAY Diversity Preserved in Indigenous Horses Revealed With Novel Y-Chromosome Markers. Front. Genet. 2020, 11, 467. [Google Scholar] [CrossRef]
- Nguyen, T.B.; Paul, R.C.; Okuda, Y.; Le, T.N.A.; Pham, P.T.K.; Kaissar, K.J.; Kazhmurat, A.; Bibigul, S.; Bakhtin, M.; Kazymbet, P.; et al. Genetic Characterization of Kushum Horses in Kazakhstan Based on Haplotypes of MtDNA and Y Chromosome, and Genes Associated with Important Traits of the Horses. J. Equine Sci. 2020, 31, 35–43. [Google Scholar] [CrossRef]
- Liu, S.; Fu, C.; Yang, Y.; Zhang, Y.; Ma, H.; Xiong, Z.; Ling, Y.; Zhao, C. Current Genetic Conservation of Chinese Indigenous Horses Revealed with Y-Chromosomal and Mitochondrial DNA Polymorphisms. G3 GenesGenomesGenetics 2021, 11, jkab008. [Google Scholar] [CrossRef]
- Cardinali, I.; Giontella, A.; Tommasi, A.; Silvestrelli, M.; Lancioni, H. Unlocking Horse Y Chromosome Diversity. Genes 2022, 13, 2272. [Google Scholar] [CrossRef]
- Gáspárdy, A.; Wagenhoffer, Z.; Fürlinger, D.; Halmágyi, M.; Bodó, I.; Lancioni, H.; Maróti-Agóts, Á. Matrilineal Composition of the Reconstructed Stock of the Szekler Horse Breed. Agriculture 2023, 13, 456. [Google Scholar] [CrossRef]
- Cardinali, I.; Lancioni, H.; Giontella, A.; Capodiferro, M.R.; Capomaccio, S.; Buttazzoni, L.; Biggio, G.P.; Cherchi, R.; Albertini, E.; Olivieri, A.; et al. An Overview of Ten Italian Horse Breeds through Mitochondrial DNA. PLoS ONE 2016, 11, e0153004. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, E.P.; Dooley, J.J.; Splan, R.K.; Bradley, D.G. Microsatellite Diversity, Pedigree Relatedness and the Contributions of Founder Lineages to Thoroughbred Horses. Anim. Genet. 2001, 32, 360–364. [Google Scholar] [CrossRef]
- Raudsepp, T.; Finno, C.J.; Bellone, R.R.; Petersen, J.L. Ten Years of the Horse Reference Genome: Insights into Equine Biology, Domestication and Population Dynamics in the Post-genome Era. Anim. Genet. 2019, 50, 569–597. [Google Scholar] [CrossRef] [PubMed]
- Radovic, L.; Remer, V.; Krcal, C.; Rigler, D.; Brem, G.; Rayane, A.; Driss, K.; Benamar, M.; Machmoum, M.; Piro, M.; et al. Y Chromosome Haplotypes Enlighten Origin, Influence, and Breeding History of North African Barb Horses. Animals 2022, 12, 2579. [Google Scholar] [CrossRef] [PubMed]
- Remer, V.; Bozlak, E.; Felkel, S.; Radovic, L.; Rigler, D.; Grilz-Seger, G.; Stefaniuk-Szmukier, M.; Bugno-Poniewierska, M.; Brooks, S.; Miller, D.C.; et al. Y-Chromosomal Insights into Breeding History and Sire Line Genealogies of Arabian Horses. Genes 2022, 13, 229. [Google Scholar] [CrossRef]
- Sharif, M.B.; Fitak, R.R.; Wallner, B.; Orozco-terWengel, P.; Frewin, S.; Fremaux, M.; Mohandesan, E. Reconstruction of the Major Maternal and Paternal Lineages in the Feral New Zealand Kaimanawa Horses. Animals 2022, 12, 3508. [Google Scholar] [CrossRef] [PubMed]
- Bozlak, E.; Radovic, L.; Remer, V.; Rigler, D.; Allen, L.; Brem, G.; Stalder, G.; Castaneda, C.; Cothran, G.; Raudsepp, T.; et al. Refining the Evolutionary Tree of the Horse Y Chromosome. Sci. Rep. 2023, 13, 8954. [Google Scholar] [CrossRef]
- Cherchi, R. l’Anglo Arabo: La Versatilità, il Cuore, la Classe di un Cavallo Europeo; C. Delfino: Sassari, Italy, 2020; ISBN 978-88-9361-185-5. [Google Scholar]
- Bigi, D.; Zanon, A. Atlante delle Razze Autoctone: Bovini, Equini, Ovicaprini, Suini Allevati in Italia, 2nd ed.; Edagricole: Milano, Italy, 2020; ISBN 978-88-506-5561-8. [Google Scholar]
- Giontella, A.; Pieramati, C.; Silvestrelli, M.; Sarti, F.M. Analysis of Founders and Performance Test Effects on an Autochthonous Horse Population through Pedigree Analysis: Structure, Genetic Variability and Inbreeding. Animal 2019, 13, 15–24. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giontella, A.; Cardinali, I.; Sarti, F.M.; Silvestrelli, M.; Lancioni, H. Y-Chromosome Haplotype Report among Eight Italian Horse Breeds. Genes 2023, 14, 1602. https://doi.org/10.3390/genes14081602
Giontella A, Cardinali I, Sarti FM, Silvestrelli M, Lancioni H. Y-Chromosome Haplotype Report among Eight Italian Horse Breeds. Genes. 2023; 14(8):1602. https://doi.org/10.3390/genes14081602
Chicago/Turabian StyleGiontella, Andrea, Irene Cardinali, Francesca Maria Sarti, Maurizio Silvestrelli, and Hovirag Lancioni. 2023. "Y-Chromosome Haplotype Report among Eight Italian Horse Breeds" Genes 14, no. 8: 1602. https://doi.org/10.3390/genes14081602
APA StyleGiontella, A., Cardinali, I., Sarti, F. M., Silvestrelli, M., & Lancioni, H. (2023). Y-Chromosome Haplotype Report among Eight Italian Horse Breeds. Genes, 14(8), 1602. https://doi.org/10.3390/genes14081602