


Distribution and Expression of Pulmonary Ionocyte-Related Factors CFTR, ATP6V0D2, and ATP6V1C2 in the Lungs of Yaks at Different Ages


Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Animals and Materials
2.2. Immunohistochemistry
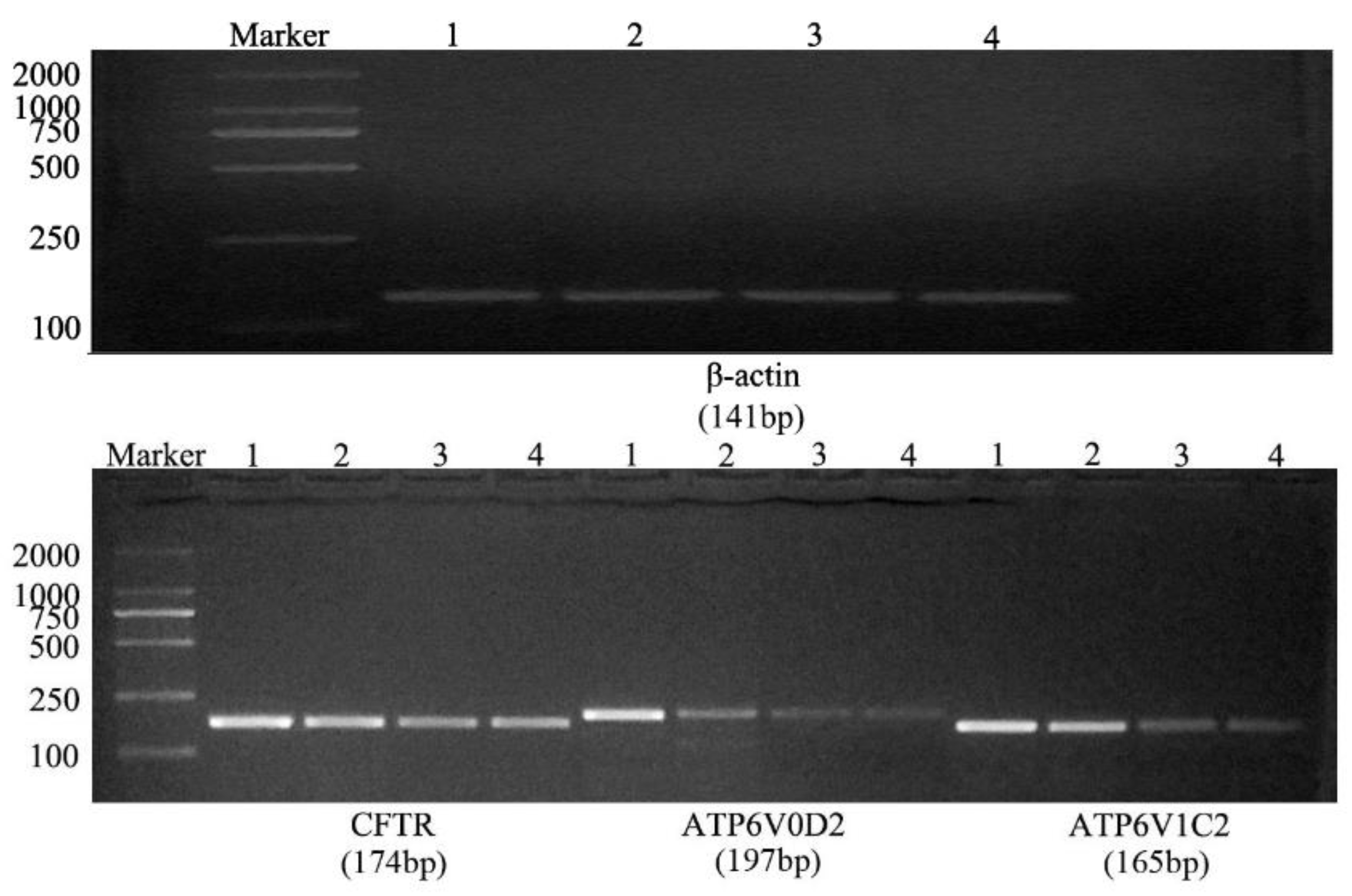
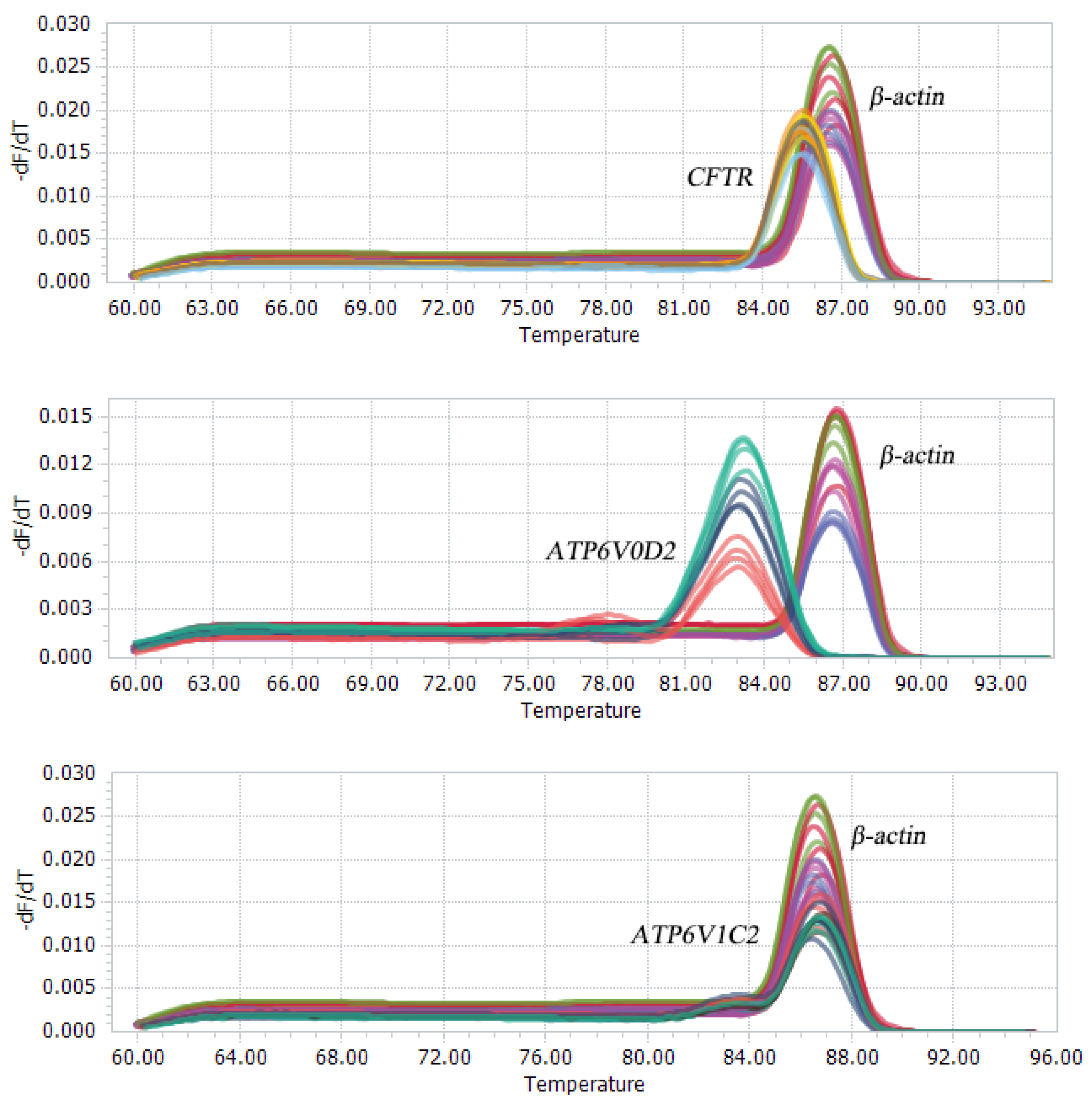
2.3. qRT-PCR
2.4. Western Blotting
3. Results
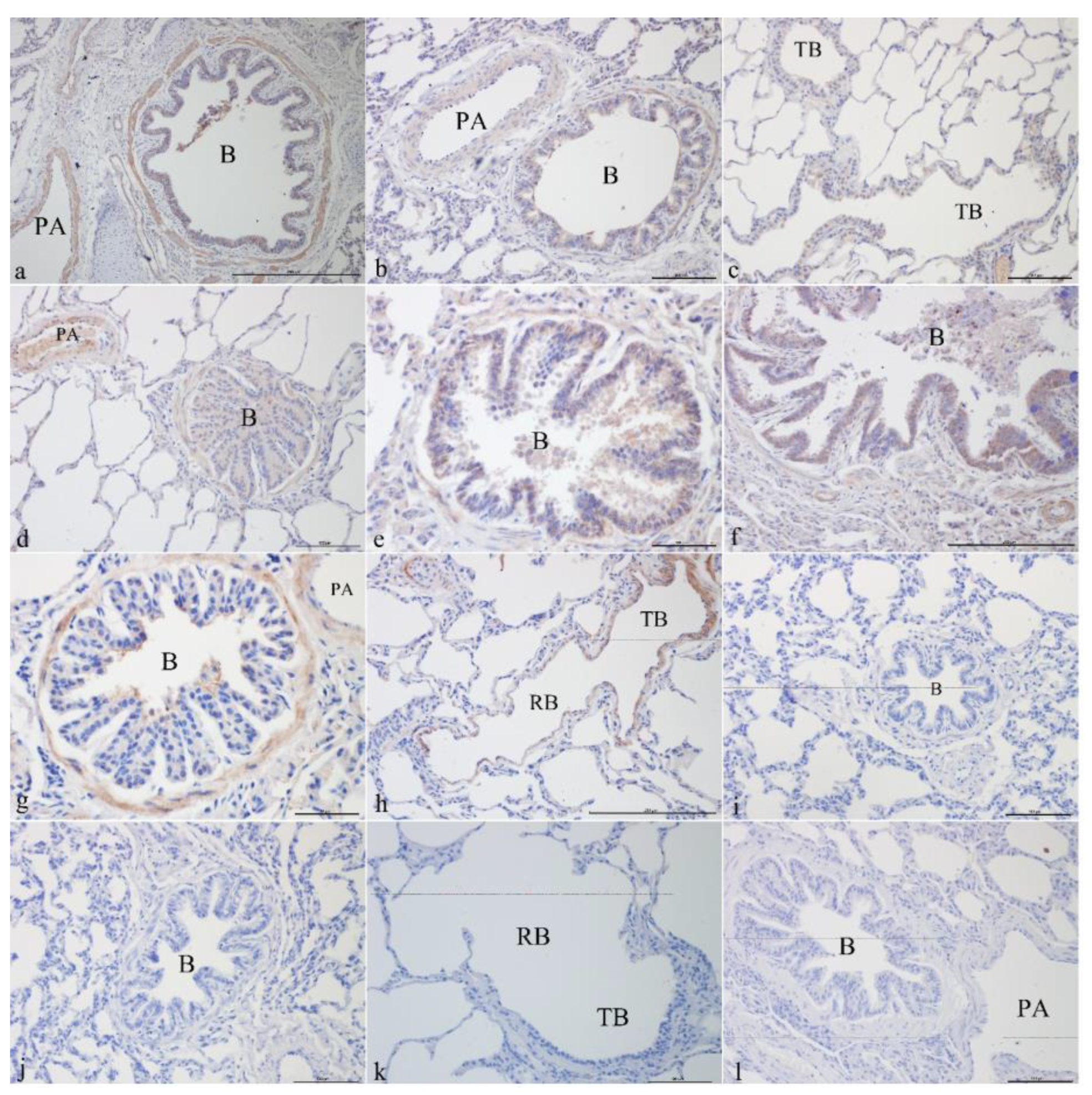
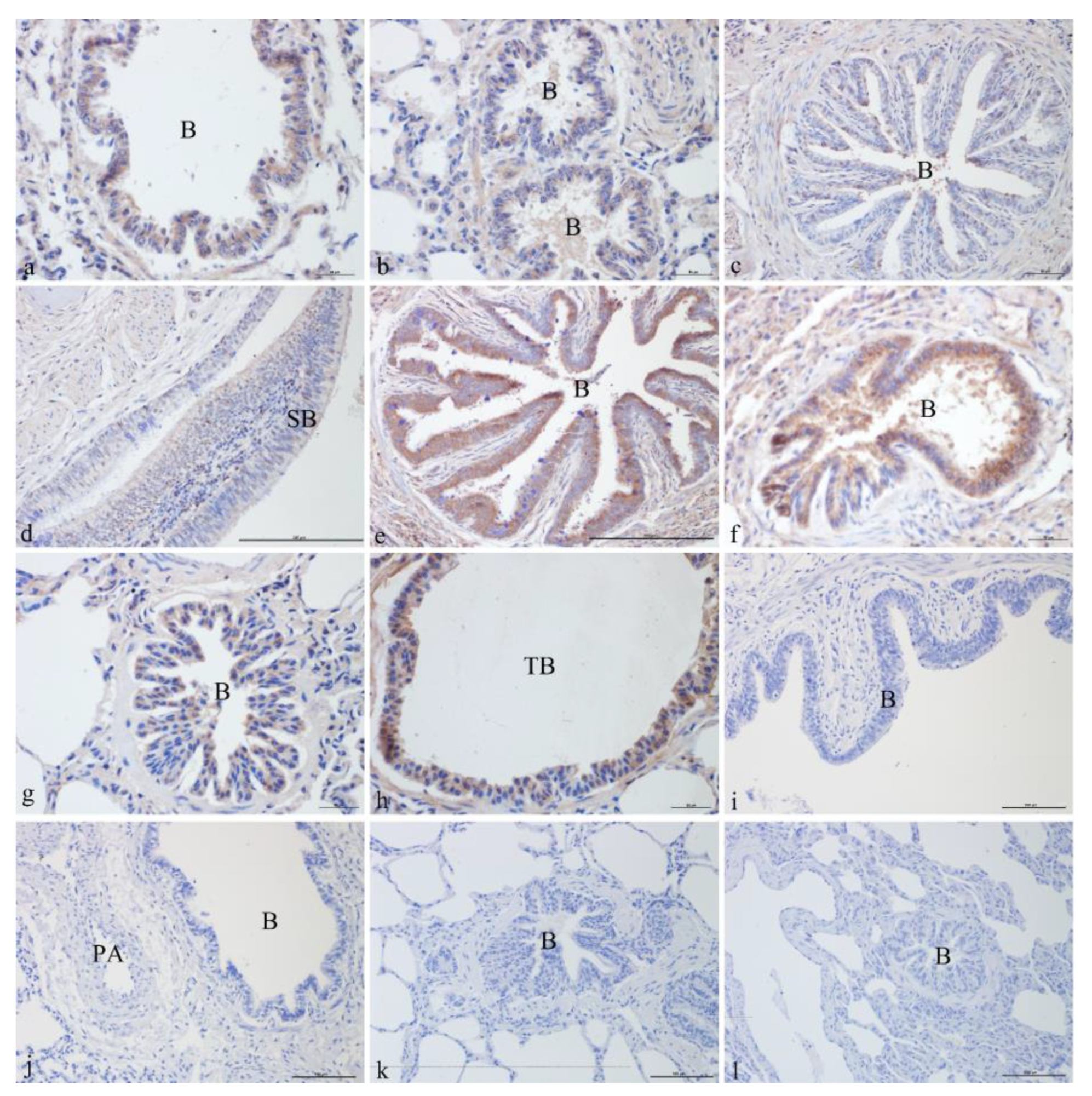
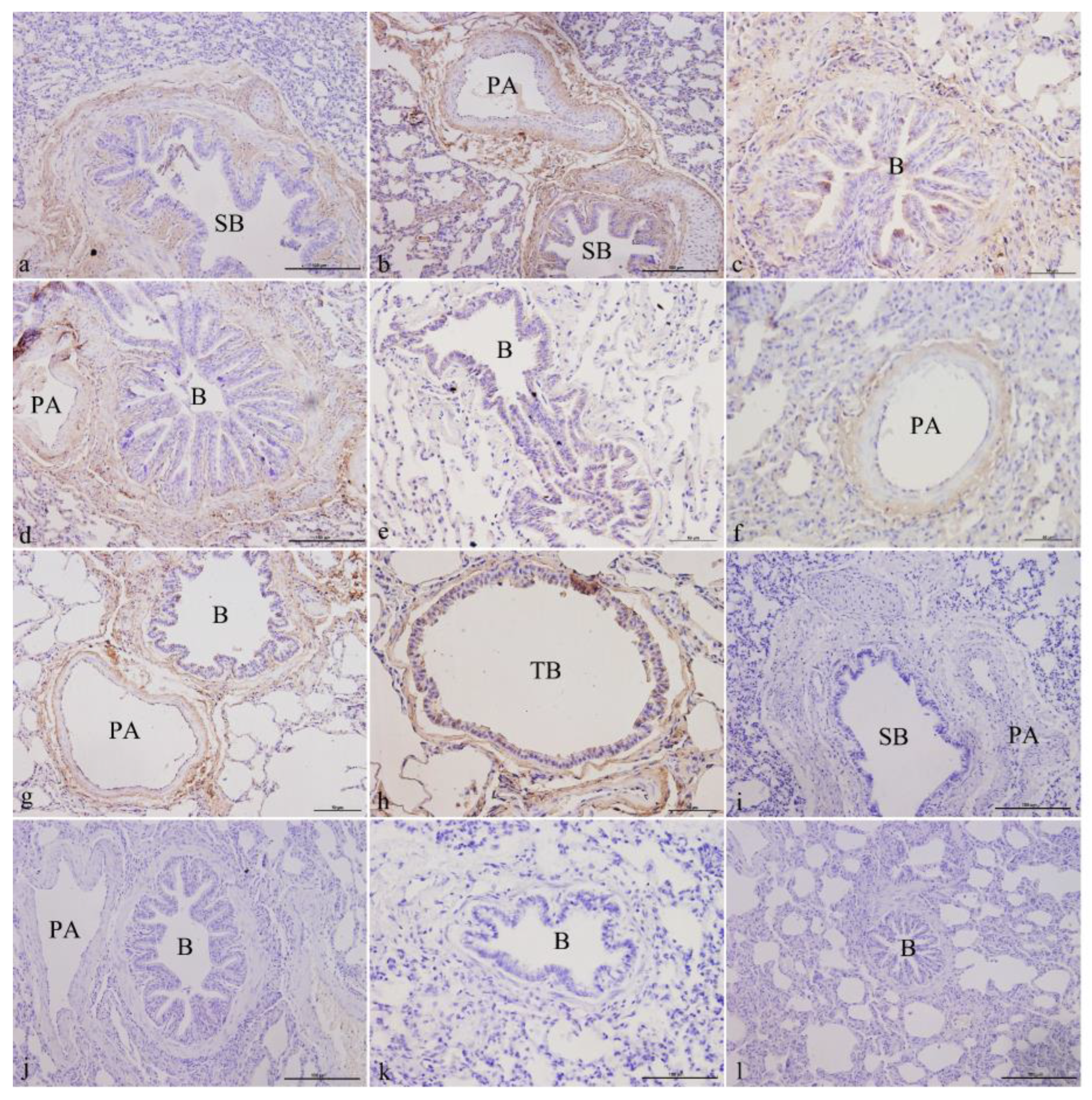
3.1. Immunohistochemical Analysis
3.1.1. CFTR Localization in Yak Lungs at Different Ages
3.1.2. ATP6V0D2 Localization in Yak Lungs at Different Ages
3.1.3. ATP6V1C2 Localization in Yak Lungs at Different Ages
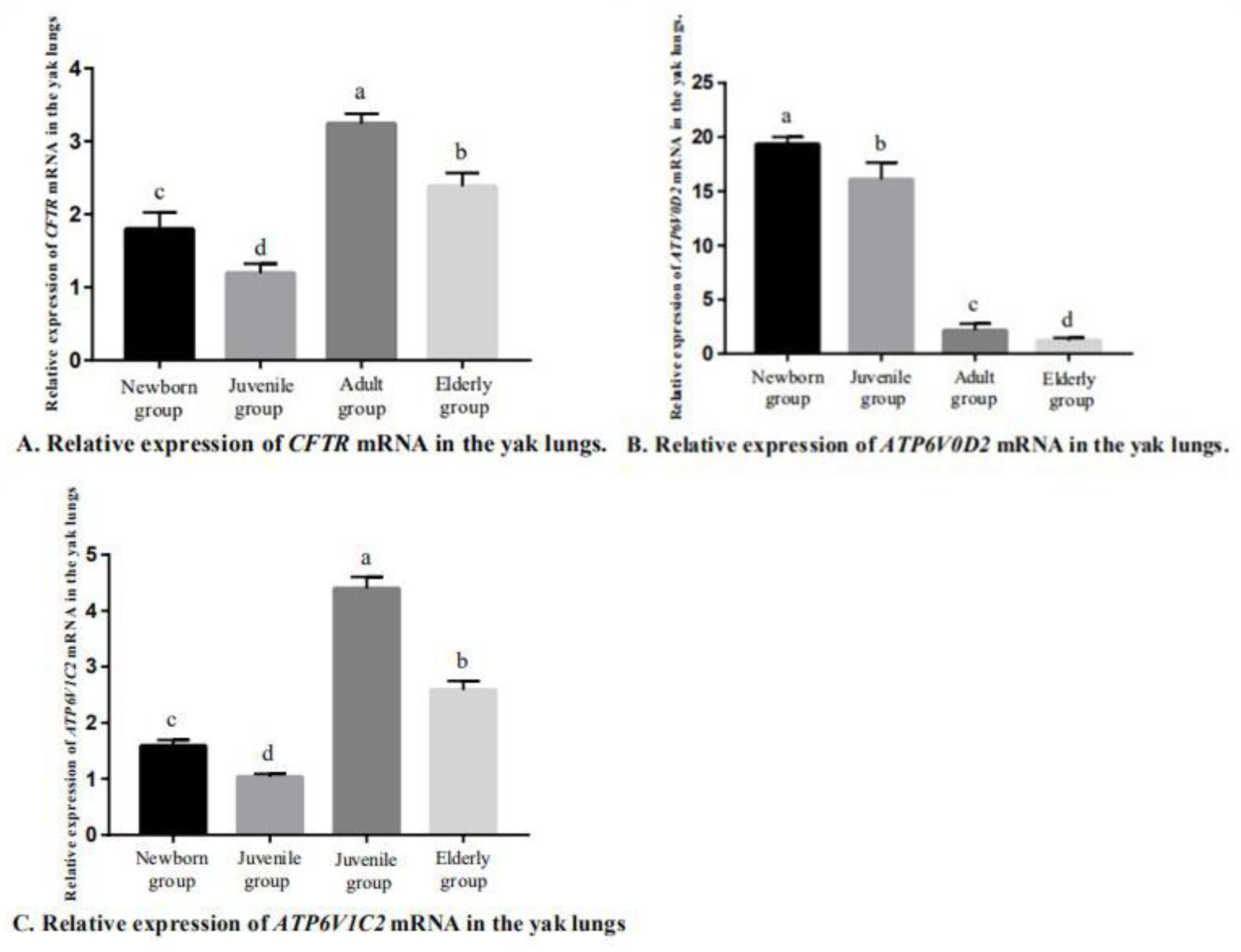
3.2. Fluorescence-Based qRT-PCR
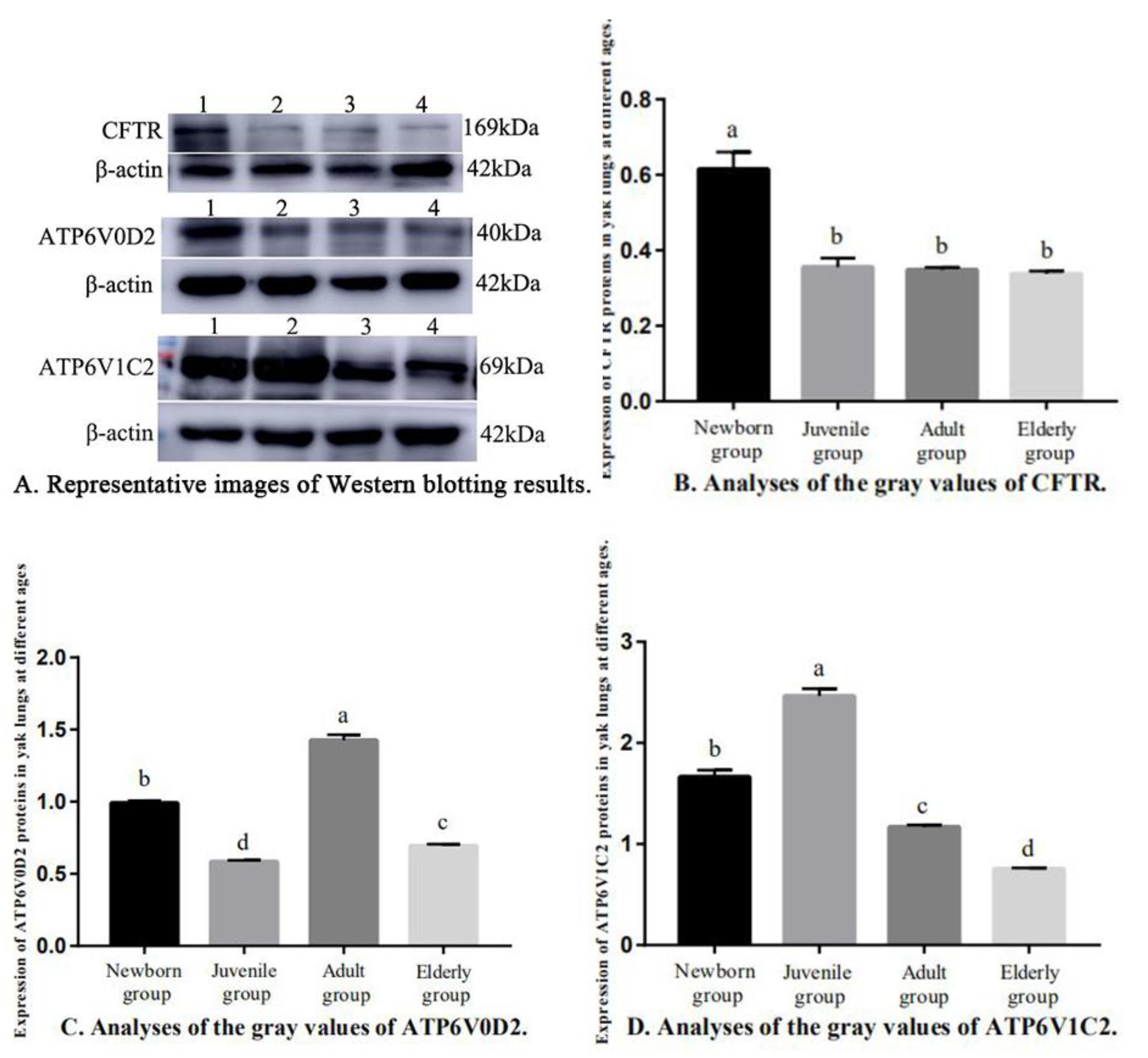
3.3. Western Blotting
4. Discussion
4.1. Pulmonary Ionocytes
4.2. CFTR
4.3. V-ATPase
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Plasschaert, L.W.; Žilionis, R.; Choo-Wing, R.; Savova, V.; Knehr, J.; Roma, G.; Klein, A.M.; Jaffe, A.B. A single-cell atlas of the airway epithelium reveals the CFTR-rich pulmonary ionocyte. Nature 2018, 560, 377–381. [Google Scholar] [CrossRef] [PubMed]
- Montoro, D.T.; Haber, A.L.; Biton, M.; Vinarsky, V.; Lin, B.; Birket, S.E.; Yuan, F.; Chen, S.; Leung, H.M.; Villoria, J.; et al. A revised airway epithelial hierarchy includes CFTR-expressing ionocytes. Nature 2018, 560, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Quigley, I.K.; Stubbs, J.L.; Kintner, C. Specification of ion transport cells in the Xenopus larval skin. Development 2011, 138, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Esaki, M.; Hoshijima, K.; Nakamura, N.; Munakata, K.; Tanaka, M.; Ookata, K.; Asakawa, K.; Kawakami, K.; Wang, W.; Weinberg, E.S.; et al. Mechanism of development of ionocytes rich in vacuolar-type H(+)-ATPase in the skin of zebrafish larvae. Dev Biol. 2009, 329, 116–129. [Google Scholar] [CrossRef] [PubMed]
- National Health Commission of the People’s Republic of China. Bulletin of the National Health Commission of the People’s Republic of China; National Health Commission of the People’s Republic of China: Beijing, China, 2018; pp. 15–19. (In Chinese) [Google Scholar]
- Kerem, B.; Rommens, J.M.; Buchanan, J.A.; Markiewicz, D.; Cox, T.K.; Chakravarti, A.; Buchwald, M.; Tsui, L.C. Identification of the cystic fibrosis gene: Genetic analysis. Science 1989, 245, 1073–1080. [Google Scholar] [CrossRef]
- Gentzsch, M.; Boyles, S.E.; Cheluvaraju, C.; Chaudhry, I.G.; Quinney, N.L.; Cho, C.; Dang, H.; Liu, X.; Schlegel, R.; Randell, S.H. Pharmacological Rescue of Conditionally Reprogrammed Cystic Fibrosis Bronchial Epithelial Cells. Am. J. Respir. Cell Mol. Biol. 2017, 56, 568–574. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Zhang, Z.; Csanády, L.; Gadsby, D.C.; Chen, J. Molecular Structure of the Human CFTR Ion Channel. Cell 2017, 169, 85–95.e8. [Google Scholar] [CrossRef]
- Cutting, G.R. Cystic fibrosis genetics: From molecular understanding to clinical application. Nat. Rev. Genet. 2015, 16, 45–56. [Google Scholar] [CrossRef]
- Feng, N.H.; Lin, H.I.; Wang, J.S.; Chou, S.T.; Ma, H.K.; Rooney, S.A.; Lu, J.F. Differential expression of a V-type ATPase C subunit gene, Atp6v1c2, during culture of rat lung type II pneumocytes. J. Biomed. Sci. 2005, 12, 899–911. [Google Scholar] [CrossRef]
- Sun-Wada, G.H.; Murata, Y.; Namba, M.; Yamamoto, A.; Wada, Y.; Futai, M. Mouse proton pump ATPase C subunit isoforms (C2-a and C2-b) specifically expressed in kidney and lung. J. Biol. Chem. 2003, 278, 44843–44851. [Google Scholar] [CrossRef]
- Shen, L.; Hu, P.; Zhang, Y.; Ji, Z.; Shan, X.; Ni, L.; Ning, N.; Wang, J.; Tian, H.; Shui, G.; et al. Serine metabolism antagonizes antiviral innate immunity by preventing ATP6V0d2-mediated YAP lysosomal degradation. Cell Metab. 2021, 33, 971–987.e6. [Google Scholar] [CrossRef]
- Xia, Y.; Liu, N.; Xie, X.; Bi, G.; Ba, H.; Li, L.; Zhang, J.; Deng, X.; Yao, Y.; Tang, Z.; et al. The macrophage-specific V-ATPase subunit ATP6V0D2 restricts inflammasome activation and bacterial infection by facilitating autophagosome-lysosome fusion. Autophagy 2019, 15, 960–975. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Feng, X.; Jiang, S. Study on the structure of adaptability of yak lung in plateau. Sci. Agric. Sin. 2006, 39, 2107–2113. (In Chinese) [Google Scholar]
- Zhang, R. Yak of China; Gansu Science and Technology Press: Lanzhou, China, 1989. (In Chinese) [Google Scholar]
- He, J.; Yu, S.; Cui, Y. Characteristics of lung structure in different age plateau yak. Acta Vet. Zootech. Sin. 2009, 40, 748–755. (In Chinese) [Google Scholar]
- Ivanis, G.; Braun, M.; Perry, S.F. Renal expression and localization of SLC9A3 sodium/hydrogen exchanger and its possible role in acid-base regulation in freshwater rainbow trout (Oncorhynchus mykiss). Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, R971–R978. [Google Scholar] [CrossRef] [PubMed]
- Qi, M.; Liu, D.M.; Ji, W.; Wang, H.L. ATP6V0D2, a subunit associated with proton transport, serves an oncogenic role in esophagus cancer and is correlated with epithelial-mesenchymal transition. Esophagus 2020, 17, 456–467. [Google Scholar] [CrossRef]
- Smith, A.N.; Borthwick, K.J.; Karet, F.E. Molecular cloning and characterization of novel tissue-specific isoforms of the human vacuolar H(+)-ATPase C, G and d subunits, and their evaluation in autosomal recessive distal renal tubular acidosis. Gene 2002, 297, 169–177. [Google Scholar] [CrossRef] [PubMed]
- Guilbault, C.; De Sanctis, J.B.; Wojewodka, G.; Saeed, Z.; Lachance, C.; Skinner, T.A.; Vilela, R.M.; Kubow, S.; Lands, L.C.; Hajduch, M.; et al. Fenretinide corrects newly found ceramide deficiency in cystic fibrosis. Am. J. Respir. Cell Mol. Biol. 2008, 38, 47–56. [Google Scholar] [CrossRef] [PubMed]
- McCarron, A.; Donnelley, M.; Parsons, D. Airway disease phenotypes in animal models of cystic fibrosis. Respir. Res. 2018, 19, 54. [Google Scholar] [CrossRef]
- Rogers, C.S.; Stoltz, D.A.; Meyerholz, D.K.; Ostedgaard, L.S.; Rokhlina, T.; Taft, P.J.; Rogan, M.P.; Pezzulo, A.A.; Karp, P.H.; Itani, O.A.; et al. Disruption of the CFTR gene produces a model of cystic fibrosis in newborn pigs. Science 2008, 321, 1837–1841. [Google Scholar] [CrossRef]
- Engelhardt, J.F.; Yankaskas, J.R.; Ernst, S.A.; Yang, Y.; Marino, C.R.; Boucher, R.C.; Cohn, J.A.; Wilson, J.M. Submucosal glands are the predominant site of CFTR expression in the human bronchus. Nat. Genet. 1992, 2, 240–248. [Google Scholar] [CrossRef]
- Trezise, A.E.; Chambers, J.A.; Wardle, C.J.; Gould, S.; Harris, A. Expression of the cystic fibrosis gene in human foetal tissues. Hum. Mol. Genet. 1993, 2, 213–218. [Google Scholar] [CrossRef]
- Jiang, Q.; Engelhardt, J.F. Cellular heterogeneity of CFTR expression and function in the lung: Implications for gene therapy of cystic fibrosis. Eur. J. Hum. Genet. 1998, 6, 12–31. [Google Scholar] [CrossRef]
- Cotter, K.; Stransky, L.; McGuire, C.; Forgac, M. Recent Insights into the Structure, Regulation, and Function of the V-ATPases. Trends Biochem. Sci. 2015, 40, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Luo, J.; Kuang, D.; Xu, S.; Duan, Y.; Xia, Y.; Wei, Z.; Xie, X.; Yin, B.; Chen, F.; et al. Lactate inhibits ATP6V0d2 expression in tumor-associated macrophages to promote HIF-2α-mediated tumor progression. J. Clin. Invest. 2019, 129, 631–646. [Google Scholar] [CrossRef] [PubMed]
- Pietrement, C.; Sun-Wada, G.H.; Silva, N.D.; McKee, M.; Marshansky, V.; Brown, D.; Futai, M.; Breton, S. Distinct expression patterns of different subunit isoforms of the V-ATPase in the rat epididymis. Biol. Reprod. 2006, 74, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.N.; Francis, R.W.; Sorrell, S.L.; Karet, F.E. The d subunit plays a central role in human vacuolar H(+)-ATPases. J. Bioenerg. Biomembr. 2008, 40, 371–380. [Google Scholar] [CrossRef]
- Yang, J.; Guo, F.; Yuan, L.; Lv, G.; Gong, J.; Chen, J. Elevated expression of the V-ATPase D2 subunit triggers increased energy metabolite levels in KrasG12D -driven cancer cells. J. Cell. Biochem. 2019, 120, 11690–11701. [Google Scholar] [CrossRef]
- Nishi, T.; Kawasaki-Nishi, S.; Forgac, M. Expression and function of the mouse V-ATPase d subunit isoforms. J. Biol. Chem. 2003, 278, 46396–46402. [Google Scholar] [CrossRef]
- Yang, K.; Yu, S.; He, J.; Cui, Y. Morphological study on fetal lung development in yak. Acta Theriol. Sin. 2012, 32, 346–355. (In Chinese) [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Sequence | Annealing (°C) | Length (bp) |
|---|---|---|---|
| CFTR | F: AAGTTGCAGATGAGGTCGGA R: GAGCACTGGGTTCATCAAGC | 57.5 | 174 |
| ATP6V0D2 | F: ACCCCTAGCTCCGTTCTTTC R: TGATGATAAAAGCACGCCGG | 57.5 | 197 |
| ATP6V1C2 | F: CCAAATATCCCGCCAAGCAG R: TCCGAGTGAAGAGGTTTCCC | 57.5 | 165 |
| β-actin | F: GCAATGAGCGGTTCC R: CCGTGTTGGCGTAGAG | 60 | 141 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, J.; Wei, Y.; Cui, Y.; Zhang, Q. Distribution and Expression of Pulmonary Ionocyte-Related Factors CFTR, ATP6V0D2, and ATP6V1C2 in the Lungs of Yaks at Different Ages. Genes 2023, 14, 597. https://doi.org/10.3390/genes14030597
He J, Wei Y, Cui Y, Zhang Q. Distribution and Expression of Pulmonary Ionocyte-Related Factors CFTR, ATP6V0D2, and ATP6V1C2 in the Lungs of Yaks at Different Ages. Genes. 2023; 14(3):597. https://doi.org/10.3390/genes14030597
Chicago/Turabian StyleHe, Junfeng, Yating Wei, Yan Cui, and Qian Zhang. 2023. "Distribution and Expression of Pulmonary Ionocyte-Related Factors CFTR, ATP6V0D2, and ATP6V1C2 in the Lungs of Yaks at Different Ages" Genes 14, no. 3: 597. https://doi.org/10.3390/genes14030597
APA StyleHe, J., Wei, Y., Cui, Y., & Zhang, Q. (2023). Distribution and Expression of Pulmonary Ionocyte-Related Factors CFTR, ATP6V0D2, and ATP6V1C2 in the Lungs of Yaks at Different Ages. Genes, 14(3), 597. https://doi.org/10.3390/genes14030597