
Comparative Analysis of the PYL Gene Family in Three Ipomoea Species and the Expression Profiling of IbPYL Genes during Abiotic Stress Response in Sweetpotato



Abstract
1. Introduction
2. Materials and Methods
2.1. The Identification of Putative PYL Genes in Three Ipomoea Species
2.2. Chromosomal Distribution and Genome-Wide Syntenic Analysis of PYL Genes
2.3. The Analysis of Gene Structures, Conserved Motifs, and Protein Properties
2.4. The Phylogenetic Classification of Three Ipomoea PYLs
2.5. RNA Extraction and qRT-PCR Analysis
2.6. Statistical Analysis
3. Results
3.1. The Identification and Chromosomal Location Analysis of PYLs in Three Ipomoea Species
3.2. A Molecular Characterization Analysis of PYLs in Three Ipomoea Species
3.3. A Phylogenetic Analysis of PYL Proteins in Three Ipomoea Species
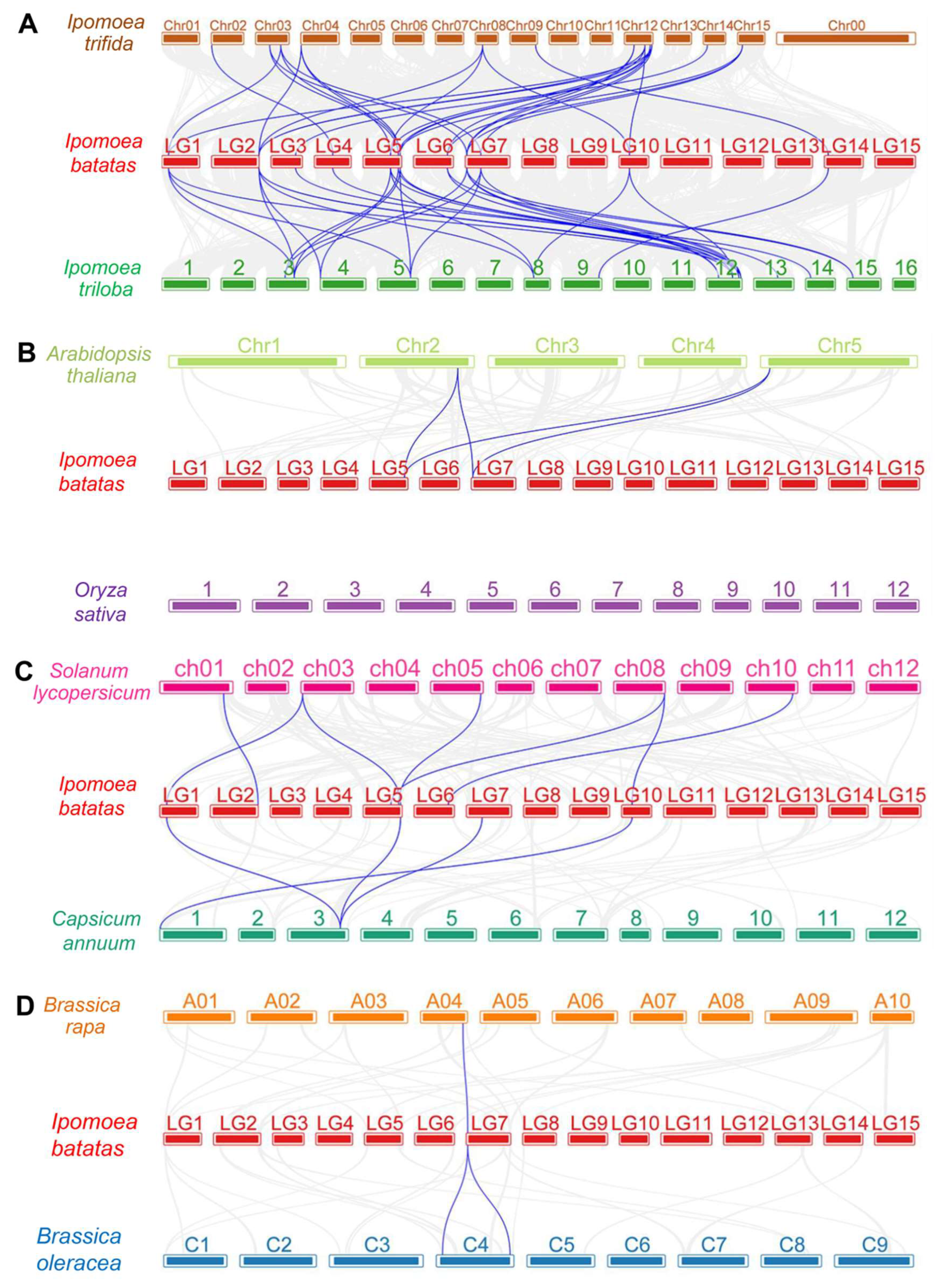
3.4. Gene Duplication and Collinearity Survey of PYL Genes
3.5. Conserved Domain and Gene Organization Analyses of PYL Proteins in Three Ipomoea Species
3.6. Conserved Motif Analyses of PYL Proteins in Three Ipomoea Species
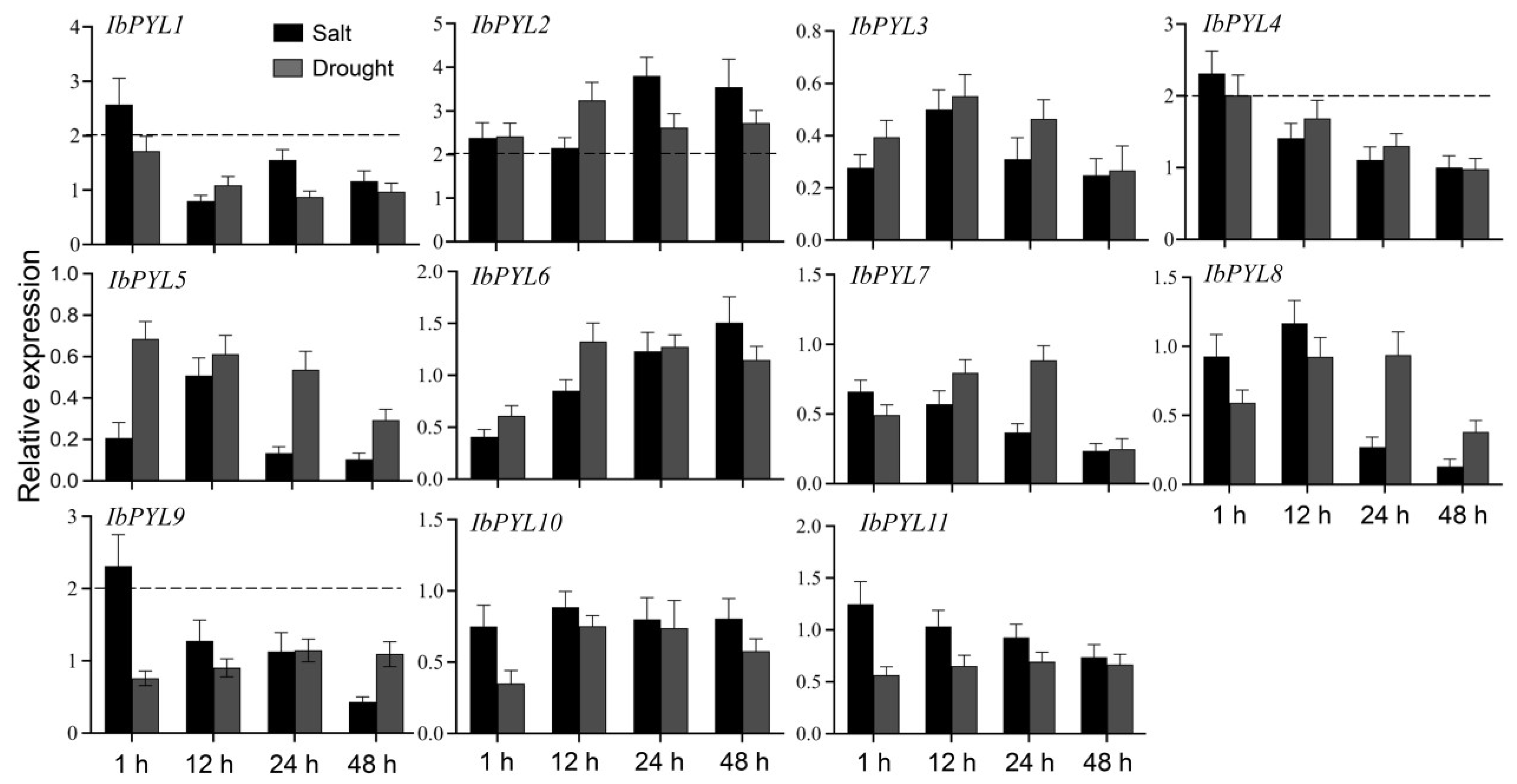
3.7. The Expression Profiles of IbPYLs under Abiotic Stress
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhu, J.K. Abiotic stress signaling and responses in plants. Cell 2016, 167, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Xiong, L.; Shi, H.; Yang, S.; Herrera-Estrella, L.R.; Xu, G.; Chao, D.-Y.; Li, J.; Wang, P.-Y.; Qin, F.; et al. Plant abiotic stress response and nutrient use efficiency. Sci. China-Life Sci. 2020, 63, 635–674. [Google Scholar]
- Zhang, H.; Zhu, J.; Gong, Z.; Zhu, J.-K. Abiotic stress responses in plants. Nat. Rev. Genet. 2022, 23, 104–119. [Google Scholar] [CrossRef] [PubMed]
- Laloum, T.; Martin, G.; Duque, P. Alternative splicing control of abiotic stress responses. Trends Plant Sci. 2018, 23, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Cutler, S.R.; Rodriguez, P.L.; Finkelstein, R.R.; Abrams, S.R. Abscisic acid: Emergence of a core signaling network. Annu. Rev. Plant Biol. 2010, 61, 651–679. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Li, G.J.; Bressan, R.A.; Song, C.P.; Zhu, J.K.; Zhao, Y. Abscisic acid dynamics, signaling, and functions in plants. J. Integr. Plant Biol. 2020, 62, 25–54. [Google Scholar] [CrossRef]
- Fujii, H.; Chinnusamy, V.; Rodrigues, A.; Rubio, S.; Antoni, R.; Park, S.Y.; Cutler, S.R.; Sheen, J.; Rodriguez, P.L.; Zhu, J.K. In vitro reconstitution of an abscisic acid signalling pathway. Nature 2009, 462, 660–664. [Google Scholar] [CrossRef]
- Umezawa, T.; Sugiyama, N.; Mizoguchi, M.; Hayashi, S.; Myouga, F.; Yamaguchi-Shinozaki, K.; Ishihama, Y.; Hirayama, T.; Shinozaki, K. Type 2C protein phosphatases directly regulate abscisic acid-activated protein kinases in Arabidopsis. Proc. Natl. Acad. Sci. USA 2009, 106, 17588–17593. [Google Scholar] [CrossRef]
- Park, S.-Y.; Fung, P.; Nishimura, N.; Jensen, D.R.; Fujii, H.; Zhao, Y.; Lumba, S.; Santiago, J.; Rodrigues, A.; Chow, T.-f.F. Abscisic acid inhibits type 2C protein phosphatases via the PYR/PYL family of START proteins. Science 2009, 324, 1068–1071. [Google Scholar] [CrossRef]
- Santiago, J.; Dupeux, F.; Betz, K.; Antoni, R.; Gonzalez-Guzman, M.; Rodriguez, L.; Márquez, J.A.; Rodriguez, P.L. Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs. Plant Sci. 2012, 182, 3–11. [Google Scholar] [CrossRef]
- Di, F.; Jian, H.; Wang, T.; Chen, X.; Ding, Y.; Du, H.; Lu, K.; Li, J.; Liu, L. Genome-wide analysis of the PYL gene family and identification of PYL genes that respond to abiotic stress in Brassica napus. Genes 2018, 9, 156. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.K.; Santosh Kumar, V.V.; Verma, R.K.; Yadav, P.; Saroha, A.; Wankhede, D.P.; Chaudhary, B.; Chinnusamy, V. Genome-wide identification and characterization of ABA receptor PYL gene family in rice. BMC Genom. 2020, 21, 676. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Lu, T.; Miao, W.; Sun, L.; Tian, M.; Wang, J.; Hao, F. Genome-wide identification of ABA receptor PYL family and expression analysis of PYLs in response to ABA and osmotic stress in Gossypium. PeerJ 2017, 5, e4126. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Luo, S.; Liu, Z.; Wan, Z.; Gao, X.; Qiao, Y.; Yu, J.; Zhang, G. Genome-wide identification and expression analysis of the cucumber PYL gene family. PeerJ 2022, 10, e12786. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; Wei, X.; Gao, R.; Huo, F.; Nie, X.; Tong, W.; Song, W. Genome-wide identification of PYL gene family in wheat: Evolution, expression and 3D structure analysis. Genomics 2021, 113, 854–866. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Chan, Z.; Gao, J.; Xing, L.; Cao, M.; Yu, C.; Hu, Y.; You, J.; Shi, H.; Zhu, Y. ABA receptor PYL9 promotes drought resistance and leaf senescence. Proc. Natl. Acad. Sci. USA 2016, 113, 1949–1954. [Google Scholar] [CrossRef]
- Zhao, Y.; Chan, Z.; Xing, L.; Liu, X.; Hou, Y.-J.; Chinnusamy, V.; Wang, P.; Duan, C.; Zhu, J.-K. The unique mode of action of a divergent member of the ABA-receptor protein family in ABA and stress signaling. Cell Res. 2013, 23, 1380–1395. [Google Scholar] [CrossRef]
- Miao, C.; Xiao, L.; Hua, K.; Zou, C.; Zhao, Y.; Bressan, R.A.; Zhu, J.-K. Mutations in a subfamily of abscisic acid receptor genes promote rice growth and productivity. Proc. Natl. Acad. Sci. USA 2018, 115, 6058–6063. [Google Scholar] [CrossRef]
- Sun, H.; Mei, J.; Zhao, W.; Hou, W.; Zhang, Y.; Xu, T.; Wu, S.; Zhang, L. Phylogenetic analysis of the SQUAMOSA promoter-binding protein-like genes in four Ipomoea species and expression profiling of the IbSPLs during storage root development in sweet potato (Ipomoea batatas). Front. Plant Sci. 2022, 12, 3291. [Google Scholar] [CrossRef]
- Ahn, Y.O.; Kim, S.H.; Kim, C.Y.; Lee, J.-S.; Kwak, S.-S.; Lee, H.-S. Exogenous sucrose utilization and starch biosynthesis among sweetpotato cultivars. Carbohydr. Res. 2010, 345, 55–60. [Google Scholar] [CrossRef]
- Liu, Q. Improvement for agronomically important traits by gene engineering in sweetpotato. Breed. Sci. 2017, 67, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Moeinzadeh, M.H.; Kuhl, H.; Helmuth, J.; Xiao, P.; Haas, S.; Liu, G.; Zheng, J.; Sun, Z.; Fan, W. Haplotype-resolved sweet potato genome traces back its hexaploidization history. Nat. Plants 2017, 3, 696. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Lau, K.H.; Cao, Q.; Hamilton, J.P.; Sun, H.; Zhou, C.; Eserman, L.; Gemenet, D.C.; Olukolu, B.A.; Wang, H.; et al. Genome sequences of two diploid wild relatives of cultivated sweetpotato reveal targets for genetic improvement. Nat. Commun. 2018, 9, 4580. [Google Scholar] [CrossRef] [PubMed]
- Xue, L.; Wei, Z.; Zhai, H.; Xing, S.; Wang, Y.; He, S.; Gao, S.; Zhao, N.; Zhang, H.; Liu, Q. The IbPYL8-IbbHLH66-IbbHLH118 complex mediates the abscisic acid-dependent drought response in sweet potato. New Phytol. 2022, 236, 2151–2171. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-h.; Jin, H.; Marler, B.; Guo, H.; et al. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Krzywinski, M.; Schein, J.; Birol, I.; Connors, J.; Gascoyne, R.; Horsman, D.; Jones, S.J.; Marra, M.A. Circos: An information aesthetic for comparative genomics. Genome Res. 2009, 19, 1639–1645. [Google Scholar] [CrossRef]
- Guo, F.; Liu, S.; Zhang, C.; Dong, T.; Meng, X.; Zhu, M. Genome-wide systematic survey and analysis of NAC transcription factor family and their response to abiotic stress in sweetpotato. Sci. Hortic. 2022, 299, 111048. [Google Scholar] [CrossRef]
- Bailey, T.L.; Johnson, J.; Grant, C.E.; Noble, W.S. The MEME Suite. Nucleic Acids Res. 2015, 43, W39–W49. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Meng, X.; Li, G.; Yu, J.; Cai, J.; Dong, T.; Sun, J.; Xu, T.; Li, Z.; Pan, S.; Ma, D.; et al. Isolation, expression analysis, and function evaluation of 12 novel stress-responsive genes of NAC transcription factors in sweetpotato. Crop Sci. 2018, 58, 1328–1341. [Google Scholar] [CrossRef]
- Park, S.C.; Kim, Y.H.; Chang, Y.J.; Park, S.; Jeong, J.C.; Lee, H.S.; Kwak, S.S. Stable internal reference genes for the normalization of real-time PCR in different sweetpotato cultivars subjected to abiotic stress conditions. PLoS ONE 2012, 7, e51502. [Google Scholar] [CrossRef] [PubMed]
- Zhu, M.; Chen, G.; Dong, T.; Wang, L.; Zhang, J.; Zhao, Z.; Hu, Z. SlDEAD31, a putative DEAD-box RNA helicase gene, regulates salt and drought tolerance and stress-related genes in tomato. PLoS ONE 2015, 10, e0133849. [Google Scholar] [CrossRef] [PubMed]
- Dupeux, F.; Santiago, J.; Betz, K.; Twycross, J.; Park, S.Y.; Rodriguez, L.; Gonzalez-Guzman, M.; Jensen, M.R.; Krasnogor, N.; Blackledge, M. A thermodynamic switch modulates abscisic acid receptor sensitivity. EMBO J. 2011, 30, 4171–4184. [Google Scholar] [CrossRef] [PubMed]
- Ying-Xian, C.; Zhi-Chao, X.; Xin-Lian, C.; Li-Ping, N.; Li-Wei, W.; Yu, W.; Jing-Yuan, S.; Hui, Y. Genome-wide identification of abscisic acid (ABA) receptor pyrabactin resistance 1-like protein (PYL) family members and expression analysis of PYL genes in response to different concentrations of ABA stress in Glycyrrhiza uralensis. Chin. J. Nat. Med. 2020, 18, 606–611. [Google Scholar]
- Hou, H.; Lv, L.; Huo, H.; Dai, H.; Zhang, Y. Genome-wide identification of the ABA receptors genes and their response to abiotic stress in apple. Plants 2020, 9, 1028. [Google Scholar] [CrossRef]
- Xu, P.; Zhang, X.; Su, H.; Liu, X.; Wang, Y.; Hong, G. Genome-wide analysis of PYL-PP2C-SnRK2s family in Camellia sinensis. Bioengineered 2020, 11, 103–115. [Google Scholar] [CrossRef]
- Liu, S.; Zhang, C.; Guo, F.; Sun, Q.; Yu, J.; Dong, T.; Wang, X.; Song, W.; Li, Z.; Meng, X. A systematical genome-wide analysis and screening of WRKY transcription factor family engaged in abiotic stress response in sweetpotato. BMC Plant Biol. 2022, 22, 616. [Google Scholar] [CrossRef]
- Nian, L.; Zhang, X.; Yi, X.; Liu, X.; Ain, N.u.; Yang, Y.; Li, X.; Haider, F.U.; Zhu, X. Genome-wide identification of ABA receptor PYL/RCAR gene family and their response to cold stress in Medicago sativa L. Physiol. Mol. Biol. Plants 2021, 27, 1979–1995. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Zhou, J.; An, F.; Sun, Y.; Guo, R.; Pan, L.; Wan, T.; Hao, Y.; Cai, Y. Genome-wide identification of the ABA receptor PYL gene family and expression analysis in Prunus avium L. Sci. Hortic. 2023, 313, 111919. [Google Scholar] [CrossRef]
- Zhang, C.; Dong, T.; Yu, J.; Hong, H.; Liu, S.; Guo, F.; Ma, H.; Zhang, J.; Zhu, M.; Meng, X. Genome-wide survey and expression analysis of Dof transcription factor family in sweetpotato shed light on their promising functions in stress tolerance. Front. Plant Sci. 2023, 14, 1140727. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Zhou, W.; Chen, F.; Luo, X.; Yang, W. Abscisic acid and gibberellins antagonistically mediate plant development and abiotic stress responses. Front. Plant Sci. 2018, 9, 416. [Google Scholar] [CrossRef]
- Verma, V.; Ravindran, P.; Kumar, P.P. Plant hormone-mediated regulation of stress responses. BMC Plant Biol. 2016, 16, 86. [Google Scholar] [CrossRef]
- Kim, H.; Lee, K.; Hwang, H.; Bhatnagar, N.; Kim, D.-Y.; Yoon, I.S.; Byun, M.-O.; Kim, S.T.; Jung, K.-H.; Kim, B.-G. Overexpression of PYL5 in rice enhances drought tolerance, inhibits growth, and modulates gene expression. J. Exp. Bot. 2014, 65, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Verma, R.K.; Santosh Kumar, V.V.; Yadav, S.K.; Pushkar, S.; Rao, M.V.; Chinnusamy, V. Overexpression of ABA receptor PYL10 gene confers drought and cold tolerance to indica rice. Front. Plant Sci. 2019, 10, 1488. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, L.; Wei, N.; Liu, Z.-H.; Hu, S.; Li, X.-B. Overexpression of cotton PYL genes in Arabidopsis enhances the transgenic plant tolerance to drought stress. Plant Physiol. Biochem. 2017, 115, 229–238. [Google Scholar] [CrossRef]
- Yang, J.; Wang, M.; Zhou, S.; Xu, B.; Chen, P.; Ma, F.; Mao, K. The ABA receptor gene MdPYL9 confers tolerance to drought stress in transgenic apple (Malus domestica). Environ. Exp. Bot. 2022, 194, 104695. [Google Scholar] [CrossRef]
- Chan, Z. Expression profiling of ABA pathway transcripts indicates crosstalk between abiotic and biotic stress responses in Arabidopsis. Genomics 2012, 100, 110–115. [Google Scholar] [CrossRef]
- Santiago, J.; Rodrigues, A.; Saez, A.; Rubio, S.; Antoni, R.; Dupeux, F.; Park, S.Y.; Márquez, J.A.; Cutler, S.R.; Rodriguez, P.L. Modulation of drought resistance by the abscisic acid receptor PYL5 through inhibition of clade A PP2Cs. Plant J. 2009, 60, 575–588. [Google Scholar] [CrossRef]
- Hao, Q.; Yin, P.; Li, W.; Wang, L.; Yan, C.; Lin, Z.; Wu, J.Z.; Wang, J.; Yan, S.F.; Yan, N. The molecular basis of ABA-independent inhibition of PP2Cs by a subclass of PYL proteins. Mol. Cell 2011, 42, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Mou, W.; Li, D.; Luo, Z.; Mao, L.; Ying, T. Transcriptomic analysis reveals possible influences of ABA on secondary metabolism of pigments, flavonoids and antioxidants in tomato fruit during ripening. PLoS ONE 2015, 10, e0129598. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Amino Acids | MW (Da) | PI | No. of Phosphorylation Site | |||
|---|---|---|---|---|---|---|---|---|
| Ser Site | Tyr Site | Thr Site | Total | |||||
| Ipomoea batatas | ||||||||
| IbPYL1 | g928.t1 | 208 | 23,607.73 | 6.17 | 9 | 3 | 6 | 18 |
| IbPYL2 | g9406.t1 | 166 | 18,635.23 | 5.5 | 7 | 3 | 5 | 15 |
| IbPYL3 | g14758.t1 | 217 | 24,651.82 | 9.88 | 17 | 1 | 11 | 29 |
| IbPYL4 | g19566.t1 | 235 | 26,974.6 | 5.36 | 9 | 3 | 5 | 17 |
| IbPYL5 | g20407.t1 | 222 | 24,209.3 | 6.72 | 13 | 0 | 8 | 21 |
| IbPYL6 | g20643.t1 | 185 | 20,584.35 | 5.8 | 12 | 2 | 4 | 18 |
| IbPYL7 | g24506.t1 | 224 | 24,246.15 | 5.5 | 14 | 1 | 11 | 26 |
| IbPYL8 | g25480.t1 | 225 | 24,211.46 | 9.01 | 22 | 3 | 12 | 37 |
| IbPYL9 | g26930.t1 | 185 | 20,834.65 | 6.17 | 12 | 3 | 4 | 19 |
| IbPYL10 | g39321.t1 | 244 | 28,064.36 | 6.85 | 9 | 3 | 4 | 16 |
| IbPYL11 | g55788.t1 | 231 | 25,788.44 | 5.38 | 15 | 1 | 14 | 30 |
| IbPYL12L | g12068.t1 | 123 | 13,787.9 | 5.72 | 4 | 1 | 4 | 9 |
| IbPYL13L | g55782.t1 | 191 | 21,202.85 | 8.74 | 8 | 1 | 9 | 18 |
| Ipomoea trifida | ||||||||
| ItfPYL1 | itf02g00240.t1 | 216 | 23,545.22 | 6.19 | 15 | 1 | 11 | 27 |
| ItfPYL2 | itf03g14430.t1 | 226 | 24,533.66 | 6.58 | 10 | 0 | 8 | 18 |
| ItfPYL3 | itf03g25530.t1 | 205 | 23,146.29 | 6.39 | 12 | 3 | 5 | 20 |
| ItfPYL4 | itf04g00430.t1 | 194 | 21,674.61 | 5.23 | 7 | 4 | 7 | 18 |
| ItfPYL5 | itf05g19440.t1 | 589 | 66,277.93 | 5.96 | 36 | 9 | 20 | 65 |
| ItfPYL6 | itf06g09740.t1 | 189 | 21,208.09 | 5.86 | 13 | 1 | 9 | 23 |
| ItfPYL7 | itf08g08150.t1 | 190 | 21,766.77 | 6.07 | 8 | 2 | 4 | 14 |
| ItfPYL8 | itf09g25130.t1 | 176 | 19,641.92 | 6.07 | 8 | 1 | 12 | 21 |
| ItfPYL9 | itf12g11260.t1 | 205 | 22,234.04 | 8.26 | 18 | 0 | 10 | 28 |
| ItfPYL10 | itf12g17820.t1 | 190 | 21,696.61 | 6.3 | 9 | 2 | 4 | 15 |
| ItfPYL11 | itf12g23670.t1 | 222 | 24,274.38 | 6.55 | 12 | 0 | 7 | 19 |
| ItfPYL12 | itf12g25980.t1 | 185 | 20,568.35 | 5.8 | 10 | 2 | 4 | 16 |
| ItfPYL13 | itf15g06620.t1 | 222 | 23,971.92 | 5.8 | 13 | 1 | 11 | 25 |
| ItfPYL14 | itf15g20740.t1 | 206 | 22,708.42 | 6.35 | 9 | 2 | 9 | 20 |
| Ipomoea triloba | ||||||||
| ItbPYL1 | itb03g15060.t1 | 222 | 24,141.25 | 6.62 | 9 | 0 | 7 | 16 |
| ItbPYL2 | itb03g23630.t1 | 185 | 20,820.62 | 6.16 | 12 | 3 | 4 | 19 |
| ItbPYL3 | itb04g00250.t1 | 188 | 20,930.69 | 5.34 | 7 | 4 | 7 | 18 |
| ItbPYL4 | itb05g20040.t1 | 208 | 23,575.67 | 6.17 | 9 | 3 | 6 | 18 |
| ItbPYL5 | itb06g07990.t1 | 189 | 21,208.09 | 5.86 | 13 | 1 | 9 | 23 |
| ItbPYL6 | itb08g08610.t1 | 190 | 21,766.77 | 6.07 | 8 | 2 | 4 | 14 |
| ItbPYL7 | itb09g28750.t1 | 231 | 25,797.45 | 5.46 | 15 | 1 | 14 | 30 |
| ItbPYL8 | itb12g11410.t1 | 211 | 22,806.61 | 6.15 | 18 | 0 | 12 | 30 |
| ItbPYL9 | itb12g18500.t1 | 190 | 21,696.61 | 6.3 | 9 | 2 | 4 | 15 |
| ItbPYL10 | itb12g24010.t1 | 222 | 24,176.31 | 6.72 | 14 | 0 | 7 | 21 |
| ItbPYL11.1 | itb12g26370.t1 | 185 | 20,616.41 | 5.8 | 12 | 2 | 4 | 18 |
| ItbPYL11.2 | itb12g26370.t2 | 185 | 20,616.41 | 5.8 | 12 | 2 | 4 | 18 |
| ItbPYL12 | itb13g12810.t1 | 216 | 23,516.19 | 6.36 | 15 | 0 | 11 | 26 |
| ItbPYL13 | itb15g06930.t1 | 224 | 24,172.11 | 5.64 | 13 | 1 | 11 | 25 |
| ItbPYL14 | itb15g21060.t1 | 210 | 23,078.74 | 6.55 | 9 | 2 | 9 | 20 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, L.; Song, W.; Xin, G.; Zhu, M.; Meng, X. Comparative Analysis of the PYL Gene Family in Three Ipomoea Species and the Expression Profiling of IbPYL Genes during Abiotic Stress Response in Sweetpotato. Genes 2023, 14, 1471. https://doi.org/10.3390/genes14071471
Zhang L, Song W, Xin G, Zhu M, Meng X. Comparative Analysis of the PYL Gene Family in Three Ipomoea Species and the Expression Profiling of IbPYL Genes during Abiotic Stress Response in Sweetpotato. Genes. 2023; 14(7):1471. https://doi.org/10.3390/genes14071471
Chicago/Turabian StyleZhang, Lei, Weihan Song, Guosheng Xin, Mingku Zhu, and Xiaoqing Meng. 2023. "Comparative Analysis of the PYL Gene Family in Three Ipomoea Species and the Expression Profiling of IbPYL Genes during Abiotic Stress Response in Sweetpotato" Genes 14, no. 7: 1471. https://doi.org/10.3390/genes14071471
APA StyleZhang, L., Song, W., Xin, G., Zhu, M., & Meng, X. (2023). Comparative Analysis of the PYL Gene Family in Three Ipomoea Species and the Expression Profiling of IbPYL Genes during Abiotic Stress Response in Sweetpotato. Genes, 14(7), 1471. https://doi.org/10.3390/genes14071471