
Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Svačina, R.; Sourdille, P.; Kopecký, D.; Bartoš, J. Chromosome pairing in polyploid grasses. Front. Plant Sci. 2020, 11, 1056. [Google Scholar] [CrossRef]
- Manrique-Carpintero, N.C.; Coombs, J.J.; Pham, G.M.; Laimbeer, F.P.E.; Braz, G.T.; Jiang, J.; Veilleux, R.E.; Buell, C.R.; Douches, D.S. Genome reduction in tetraploid potato reveals genetic load, haplotype variation, and loci associated with agronomic traits. Front. Plant Sci. 2018, 9, 944. [Google Scholar] [CrossRef]
- Lian, Q.; Tang, D.; Bai, Z.; Qi, J.; Lu, F.; Huang, S.; Zhang, C. Acquisition of deleterious mutations during potato polyploidization. J. Integr. Plant Biol. 2019, 61, 7–11. [Google Scholar] [CrossRef]
- Zhang, C.; Wang, P.; Tang, D.; Yang, Z.; Lu, F.; Qi, J.; Tawari, N.R.; Shang, Y.; Li, C.; Huang, S. The genetic basis of inbreeding depression in potato. Nat. Genet. 2019, 51, 374–378. [Google Scholar] [CrossRef] [PubMed]
- Labroo, M.R.; Endelman, J.B.; Gemenet, D.C.; Werner, C.R.; Gaynor, R.C.; Covarrubias-Pazaran, G.E. Clonal breeding strategies to harness heterosis insights from stochastic simulation. bioRxiv 2022. [Google Scholar] [CrossRef]
- Charlesworth, D.; Willis, J. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Busbice, T.H.; Wilsie, C.P. Inbreeding depression and heterosis in autotetraploids with application to Medicago sativa L. Euphytica 1966, 15, 52–67. [Google Scholar] [CrossRef]
- Glendinning, D.R. Estimation of inbreeding in potato pedigrees. Potato Res. 1997, 40, 277–284. [Google Scholar] [CrossRef]
- Malecot, G. Les Mathámatiques de L’hérédité; Masson et Cie: Paris, France, 1948. [Google Scholar]
- Kempthorne, O. An Introduction to Genetic Statistics; Iowa State University Press: Ames, Iowa, 1957. [Google Scholar]
- Mendoza, H.A.; Haynes, F.L. Some aspects of breeding and inbreeding in potatoes. Am. Potato J. 1973, 50, 216–222. [Google Scholar] [CrossRef]
- Hagberg, A.; Tedin, O. Inter- and intra-clonal crosses and inbreeding in potatoes. Hereditas 1951, 37, 280–287. [Google Scholar] [CrossRef]
- Mendoza, H.A.; Haynes, F.L. Genetic relationship among potato cultivars grown in the United States. HortScience 1974, 9, 328–330. [Google Scholar] [CrossRef]
- Chen, Z.J. Molecular mechanisms of polyploidy and hybrid vigor. Trends Plant Sci. 2010, 15, 57–71. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, H.A.; Haynes, F.L. Genetic basis of heterosis for yield in the autotetraploid potato. Theor. Appl. Genet. 1974, 45, 21–25. [Google Scholar] [CrossRef]
- Bonierbale, M.W.; Plaisted, R.L.; Tanksley, S.D. A test of the maximum heterozygosity hypothesis using molecular markers in tetraploid potatoes. Theor. Appl. Genet. 1993, 86, 481–491. [Google Scholar] [CrossRef]
- Muthoni, J.; Shimelis, H.; Melis, R. Production of hybrid potatoes: Are heterozygosity and ploidy levels important? Aust. J. Crop Sci. 2019, 13, 687–694. [Google Scholar] [CrossRef]
- Zhang, C.; Yang, Z.; Tang, D.; Zhu, Y.; Wang, P.; Li, D.; Zhu, G.; Xiong, X.; Shang, Y.; Li, C.; et al. Genome design of hybrid potato. Cell 2021, 84, 3873–3883.e12. [Google Scholar] [CrossRef]
- Hirsch, C.N.; Hirsch, C.D.; Felcher, K.; Coombs, J.; Zarka, D.; Van Deynze, A.; De Jong, W.; Veilleux, R.E.; Jansky, S.; Bethke, P.; et al. Retrospective view of North American potato (Solanum tuberosum L.) breeding in the 20th and 21st centuries. G3 2013, 3, 1003–1013. [Google Scholar] [CrossRef] [PubMed]
- Ramu, P.; Esuma, W.; Kawuki, R.; Rabbi, I.Y.; Egesi, C.; Bredeson, J.V.; Bart, R.S.; Verma, J.; Buckler, E.S.; Lu, F. Cassava haplotype map highlights fixation of deleterious mutations during clonal propagation. Nat. Genet. 2017, 49, 959–963. [Google Scholar] [CrossRef]
- Karunarathna, N.L. Genomic background selection to reduce the mutation load after random mutagenesis. Sci. Rep. 2021, 11, 19404. [Google Scholar] [CrossRef]
- Momo, S.; Jannink, J.-L. Incorporating selfing to purge deleterious alleles in a cassava genomic selection program. bioRxiv 2020. [Google Scholar] [CrossRef]
- Federer, W. Augmented designs. Hawaii. Plant. Recorder. 1956, 55, 191–208. [Google Scholar] [CrossRef]
- Mann, D.J.; Lammerink, J.P.; Coles, G.D. Predicting potato crisp darkening: Two methods for analysis of glucose. N. Z. J. Crop Hortic. Sci. 1991, 19, 199–201. [Google Scholar] [CrossRef]
- Selga, S.; Reslow, R.; Pérez-Rodríguez, P.; Ortiz, R. The power of genomic estimated breeding values for selection when using a finite population size in genetic improvement of tetraploid potato. G3 2022, 12, jkab362. [Google Scholar] [CrossRef] [PubMed]
- Huaman, Z.; Williams, J.T.; Salhuana, W.; Vincent, L. Descriptors for the Cultivated Potato and the Maintenance and Distribution of Germplasm Collections; International Board for Plant Genetic Resources: Rome, Italy, 1977. [Google Scholar]
- Selga, C.; Koc, A.; Chawade, A.; Ortiz, R. A bioinformatics pipeline to identify a subset of SNPs for genomics-assisted potato breeding. Plants 2021, 10, 30. [Google Scholar] [CrossRef]
- Oliveira, I.B.; Resende, M.F.R., Jr.; Ferrão, L.F.V.; Amadeu, R.R.; Endelman, J.B.; Kirst, M.; Coelho, A.S.G.; Munoz, P.R. Genomic prediction of autotetraploids: Influence of relationship matrices, allele dosage, and continuous genotyping calls in phenotype prediction. G3 2019, 9, 1189–1198. [Google Scholar] [CrossRef]
- Ortiz, R.; Crossa, J.; Reslow, F.; Perez-Rodriguez, P.; Cuevas, J. Genome-based genotype environment prediction enhances potato (Solanum tuberosum L.) improvement using pseudo-diploid and polysomic tetraploid modeling. Front. Plant Sci. 2022, 13, 785196. [Google Scholar] [CrossRef]
- Cotes, T.J.M.; Ñústez, L.C.E. Propuesta para el análisis de diseños aumentados en fitomejoramiento: Un caso en papa. Rev. Latinoam. Papa 2001, 12, 15–34. [Google Scholar] [CrossRef]
- Gardner, C.O.; Eberhart, S.A. Analysis and interpretation of the variety cross diallel and related populations. Biometrics 1966, 22, 439–452. [Google Scholar] [CrossRef]
- Slater, A.T.; Cogan, N.O.; Forster, J.W.; Hayes, B.J.; Daetwyler, H.D. Improving genetic gain with genomic selection in autotetraploid potato. Plant Genome 2016, 9, plantgenome2016.02.0021. [Google Scholar] [CrossRef]
- Amadeu, R.R.; Cellon, C.; Olmstead, J.W.; Garcia, A.A.F.; Resende, M.F.R.; Munoz, P.R. AGHmatrix: R package to construct relationship matrices for autotetraploid and diploid species: A blueberry example. Plant Genome 2016, 9, plantgenome2016.01.0009. [Google Scholar] [CrossRef]
- Pérez, P.; de los Campos, G. Genome-wide regression and prediction with the BGLR statistical package. Genetics 2014, 198, 483–495. [Google Scholar] [CrossRef] [PubMed]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 9 January 2023).
- Hoopes, G.M.; Zarka, D.; Feke, A.; Acheson, K.; Hamilton, J.P.; Douches, D.; Buell, C.R.; Farré, E.M. Keeping time in the dark: Potato diel and circadian rhythmic gene expression reveals tissue-specific circadian clocks. Plant Direct 2022, 6, e425. [Google Scholar] [CrossRef] [PubMed]
- Niu, Y.; Li, G.; Jiang, Y.; Duan, S.; Liu, J.; Xu, J.; Jin, L. Genes related to circadian rhythm are involved in regulating tuberization time in potato. Hortic. Plant J. 2022, 8, 369–380. [Google Scholar] [CrossRef]
- Zhou, T.; Song, B.; Liu, T.; Shen, Y.; Dong, L.; Jing, S.; Xie, C.; Liu, J. Phytochrome F plays critical roles in potato photoperiodic tuberization. Plant J. 2019, 98, 42–54. [Google Scholar] [CrossRef]
- Rodríguez-Falcón, M.; Bou, J.; Prat, S. Seasonal control of tuberization in potato: Conserved elements with the flowering response. Annu. Rev. Plant Biol. 2006, 57, 151–180. [Google Scholar] [CrossRef] [PubMed]
- Golmirzaie, A.; Ortiz, R.; Atlin, G.; Iwanaga, M. Inbreeding and true seed in tetrasomic potato. I. Selfing and open pollination in Andean landraces (Solanum tuberosum Gp. Andigena). Theor. Appl. Genet. 1998, 97, 1125–1128. [Google Scholar] [CrossRef]
- Golmirzaie, A.; Bretschneider, K.; Ortiz, R. Inbreeding and true seed in tetrasomic potato. II. Selfing and sib-mating in heterogeneous hybrid populations of Solanum tuberosum. Theor. Appl. Genet. 1998, 97, 1129–1132. [Google Scholar] [CrossRef]
- Krantz, F.A. Potato breeding methods. In Technical Bulletin; University of Minnesota Department of Agriculture: St Paul, MN, USA, 1924; Volume 25. [Google Scholar]
- Krantz, F.A.; Hutchins, A.E. Technical Bulletin; University of Minnesota Department of Agriculture: St Paul, MN, USA, 1929; Volume 58. [Google Scholar]
- Mendiburu, A.O.; Peloquin, S.J. The significance of 2n gametes in potato breeding. Theor. Appl. Genet. 1977, 49, 53–61. [Google Scholar] [CrossRef]
- Batista, L.G.; Mello, V.H.; Souza, A.P.; Margarido, G.R.A. Genomic prediction with allele dosage information in highly polyploid species. Theor. Appl. Genet. 2022, 135, 723–739. [Google Scholar] [CrossRef]
- Amadeu, R.R.; Ferrão, L.F.V.; Oliveira, I.d.B.; Benevenuto, J.; Endelman, J.B.; Munoz, P.R. Impact of dominance effects on autotetraploid genomic prediction. Crop Sci. 2020, 60, 656–665. [Google Scholar] [CrossRef]
- Ortiz, R.; Reslow, F.; Crossa, J.; Cuevas, J. Heritable variation, genetic and phenotypic correlations for tuber traits and host plant resistance to late blight for potato breeding in Scandinavian testing sites. Agriculture 2021, 11, 1287. [Google Scholar] [CrossRef]
- Endelman, J.B.; Carley, C.A.S.; Bethke, P.C.; Coombs, J.J.; Clough, M.E.; da Silva, W.L.; De Jong, W.S.; Douches, D.S.; Frederick, C.M.; Haynes, K.G.; et al. Genetic variance partitioning and genome-wide prediction with allele dosage information in autotetraploid potato. Genetics 2018, 209, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Cuevas, J.; Reslow, F.; Crossa, J.; Ortiz, R. Modeling genotype × environment interaction for single and multi-trait genomic prediction in potato (Solanum tuberosum L.). G3 2022, 13, jkac322. [Google Scholar] [CrossRef] [PubMed]
- Zingaretti, L.M.; Gezan, S.A.; Ferrão, L.F.V.; Osorio, L.F.; Monfort, A.; Muñoz, P.R.; Whitaker, V.M.; Pérez-Enciso, M. Exploring deep learning for complex trait genomic prediction in polyploid outcrossing species. Front. Plant Sci. 2020, 11, 25. [Google Scholar] [CrossRef]
- Atlin, G. Inbreeding in TPS progenies: Implications for breeding and seed production strategy. In Proceedings of the Present and Future Strategies for Potato Breeding and Improvement: Report of the XXVI Planning Conference, Lima, Peru, 12–14 December 1983; Centro Internacional de la Papa: Lima, Peru, 1985; pp. 71–85. [Google Scholar]


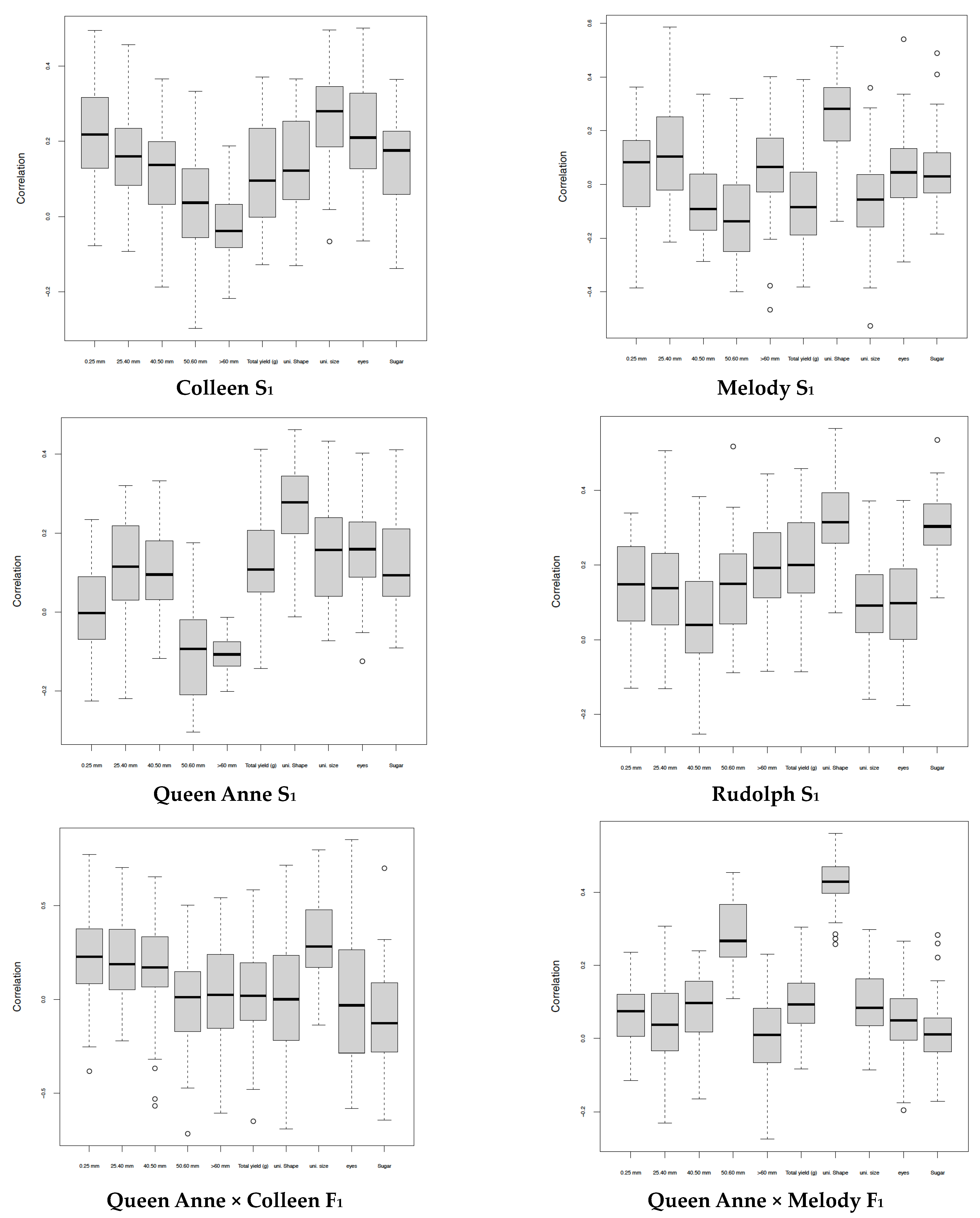{kind=link}
| Cultivar or Offspring | Tuber Weight (4-Plant Plot, g) | Tuber Uniformity Z | Tuber Eye Depth Y | Tuber Flesh Reducing Sugar | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| <25 mm | 25–40 mm | 40–50 mm | 50–60 mm | >60 mm | Total | Shape | Size | |||
| Cultivars (S0) | ||||||||||
| Colleen | 34 | 280 | 528 | 1118 | 810 | 2771 | 5.3 | 5.3 | 5.0 | 0.00 |
| Melody | 23 | 452 | 844 | 1349 | 637 | 3304 | 5.3 | 6.6 | 5.0 | 0.00 |
| Queen Anne | 43 | 513 | 1010 | 721 | 161 | 2247 | 6.1 | 5.6 | 6.0 | 0.22 |
| First inbred generation (S1) | ||||||||||
| Colleen | 39 | 219 | 240 | 176 | 51 | 725 | 5.2 | 5.2 | 4.6 | 0.32 |
| Melody | 66 | 291 | 298 | 103 | 59 | 781 | 5.4 | 4.9 | 4.9 | 0.22 |
| Queen Anne | 60 | 249 | 139 | 67 | 11 | 497 | 6.7 | 5.4 | 5.4 | 0.47 |
| Rudolph | 26 | 133 | 248 | 284 | 214 | 889 | 5.0 | 4.6 | 4.0 | 0.92 |
| Hybrid offspring (F1) | ||||||||||
| Queen Anne × Colleen | 80 | 257 | 378 | 312 | 136 | 1162 | 6.0 | 4.6 | 4.7 | 0.69 |
| Queen Anne × Melody | 53 | 318 | 497 | 552 | 169 | 1546 | 6.1 | 4.7 | 4.9 | 0.42 |
| LSD0.05 | 27 | 104 | 145 | 189 | 136 | 378 | 0.9 | 0.5 | 0.4 | 0.41 |
| Statistical significance (P > Fc) of contrasts | ||||||||||
| S0 vs. S1 | 0.1086 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.8898 | <0.0001 | <0.0001 | 0.0031 |
| S0 vs. F1 | 0.0008 | 0.0008 | <0.0001 | <0.0001 | <0.0001 | <0.0001 | 0.0228 | <0.0001 | 0.0002 | 0.0012 |
| S1 vs. F1 | 0.0002 | 0.0014 | <0.0001 | <0.0001 | 0.0059 | <0.0001 | <0.0001 | <0.0001 | 0.1618 | 0.3104 |
| Training set | Testing sets | |||||
|---|---|---|---|---|---|---|
| ‘Queen Anne’ S1 | ‘Colleen’ S1 | ‘Melody’ S1 | ‘Queen Anne’ × ‘Colleen’ F1 | ‘Queen Anne’ × ‘Melody’ F1 | ‘Rudolph’ S1 | |
| ‘Queen Anne’ S1 | ||||||
| ‘Colleen’ S1 | ||||||
| ‘Melody’ S1 | ||||||
| ‘Queen Anne’ × ‘Colleen’ F1 | ||||||
| ‘Queen Anne’ × ‘Melody’ F1 | ||||||
| ‘Rudolph’ S1 | ||||||
| Training and Validating Offspring | Tuber Weight (4-Plant Plot, g) | Tuber Uniformity | Tuber Eye Depth | Tuber Flesh Reducing Sugar | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| <25 mm | 25–40 mm | 40–50 mm | 50–60 mm | >60 mm | Total | Shape | Size | |||
| Full-sibs S1 inbred offspring | ||||||||||
| A. Colleen | 0.227 | 0.145 | 0.120 | 0.032 | −0.022 | 0.105 | 0.136 | 0.266 | 0.217 | 0.142 |
| B. Melody | 0.050 | 0.119 | −0.048 | −0.120 | 0.061 | −0.070 | 0.246 | −0.055 | 0.045 | 0.056 |
| C. Queen Anne | −0.007 | 0.143 | 0.122 | −0.196 | −0.053 | −0.143 | 0.109 | 0.186 | −0.125 | 0.218 |
| D. Rudolph | 0.137 | 0.136 | 0.061 | 0.138 | 0.202 | 0.206 | 0.316 | 0.097 | 0.090 | 0.297 |
| Full-sibs F1 hybrid offspring | ||||||||||
| Y. Queen Anne × Colleen | 0.218 | 0.213 | 0.157 | 0.002 | 0.026 | 0.036 | 0.009 | 0.320 | −0.001 | −0.096 |
| Z. Queen Anne × Melody | 0.070 | 0.049 | 0.088 | 0.287 | 0.011 | 0.094 | 0.420 | 0.095 | 0.048 | 0.021 |
| Inbred full-sibs S1 (training population)—half-sib F1 hybrids (breeding population) | ||||||||||
| A.–Y. | 0.024 | 0.083 | 0.232 | 0.100 | 0.111 | 0.311 | 0.444 | 0.104 | 0.165 | 0.245 |
| B.–Z. | −0.088 | −0.013 | 0.140 | 0.059 | 0.108 | 0.100 | 0.217 | −0.085 | 0.060 | 0.014 |
| C.–Y. | 0.221 | −0.062 | −0.100 | −0.152 | −0.393 | −0.224 | 0.106 | 0.310 | 0.235 | 0.379 |
| C.–Z. | 0.159 | 0.076 | 0.156 | 0.208 | 0.100 | 0.264 | 0.135 | 0.026 | 0.030 | 0.112 |
| Half-sib F1 hybrids (one as a training population and the other as a breeding population) | ||||||||||
| Y.–Z. | 0.100 | −0.110 | 0.141 | −0.148 | −0.093 | −0.057 | −0.042 | −0.084 | 0.018 | −0.023 |
| Z.–Y. | 0.147 | −0.153 | 0.354 | −0.266 | −0.206 | −0.011 | 0.166 | −0.226 | 0.104 | −0.275 |
| Inbred S1 (training population)—non-related F1 (breeding population) | ||||||||||
| A.–Z. | 0.096 | 0.114 | −0.078 | −0.109 | 0.056 | −0.021 | 0.066 | −0.072 | 0.076 | 0.158 |
| B.–Y. | −0.170 | 0.090 | 0.079 | 0.171 | −0.151 | 0.282 | 0.105 | −0.008 | −0.082 | −0.098 |
| D.–Y. | 0.214 | 0.164 | 0.173 | 0.116 | −0.037 | 0.001 | −0.040 | −0.136 | 0.130 | 0.097 |
| D.–Z. | 0.106 | 0.136 | 0.160 | −0.029 | 0.003 | 0.123 | 0.225 | 0.064 | 0.024 | 0.012 |
| Among inbred S1 offspring (one as a training population and the other as a breeding population) | ||||||||||
| A.–B. | −0.227 | −0.074 | −0.015 | −0.132 | 0.202 | −0.058 | 0.171 | −0.118 | 0.028 | −0.281 |
| A.–C. | 0.042 | −0.139 | −0.096 | 0.131 | 0.077 | 0.093 | −0.013 | 0.035 | 0.193 | −0.077 |
| B.–C. | −0.083 | −0.050 | 0.083 | −0.052 | 0.076 | 0.099 | −0.098 | 0.166 | −0.134 | 0.185 |
| B.–A. | −0.173 | −0.013 | 0.034 | −0.091 | −0.014 | 0.004 | 0.140 | −0.055 | 0.093 | −0.127 |
| C.–A. | 0.081 | −0.111 | −0.125 | 0.053 | −0.002 | 0.054 | 0.018 | 0.116 | 0.165 | −0.083 |
| C.–B. | −0.185 | −0.164 | 0.044 | 0.108 | −0.058 | −0.042 | −0.076 | 0.067 | −0.028 | 0.193 |
| D.–A. | 0.154 | 0.148 | −0.033 | 0.048 | 0.110 | 0.118 | 0.230 | −0.050 | 0.094 | −0.053 |
| D.–B. | −0.097 | −0.151 | −0.013 | −0.008 | 0.024 | 0.014 | 0.002 | −0.066 | −0.295 | 0.146 |
| D.–C. | −0.080 | −0.066 | 0.019 | −0.069 | 0.174 | 0.030 | 0.006 | −0.179 | 0.015 | 0.136 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ortiz, R.; Reslow, F.; Vetukuri, R.; García-Gil, M.R.; Pérez-Rodríguez, P.; Crossa, J. Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes. Genes 2023, 14, 1302. https://doi.org/10.3390/genes14061302
Ortiz R, Reslow F, Vetukuri R, García-Gil MR, Pérez-Rodríguez P, Crossa J. Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes. Genes. 2023; 14(6):1302. https://doi.org/10.3390/genes14061302
Chicago/Turabian StyleOrtiz, Rodomiro, Fredrik Reslow, Ramesh Vetukuri, M. Rosario García-Gil, Paulino Pérez-Rodríguez, and José Crossa. 2023. "Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes" Genes 14, no. 6: 1302. https://doi.org/10.3390/genes14061302
APA StyleOrtiz, R., Reslow, F., Vetukuri, R., García-Gil, M. R., Pérez-Rodríguez, P., & Crossa, J. (2023). Inbreeding Effects on the Performance and Genomic Prediction for Polysomic Tetraploid Potato Offspring Grown at High Nordic Latitudes. Genes, 14(6), 1302. https://doi.org/10.3390/genes14061302