
Identification of SNPs Related to Salmonella Resistance in Chickens Using RNA-Seq and Integrated Bioinformatics Approach
 , and
, and
Abstract
1. Introduction
2. Materials and Methods
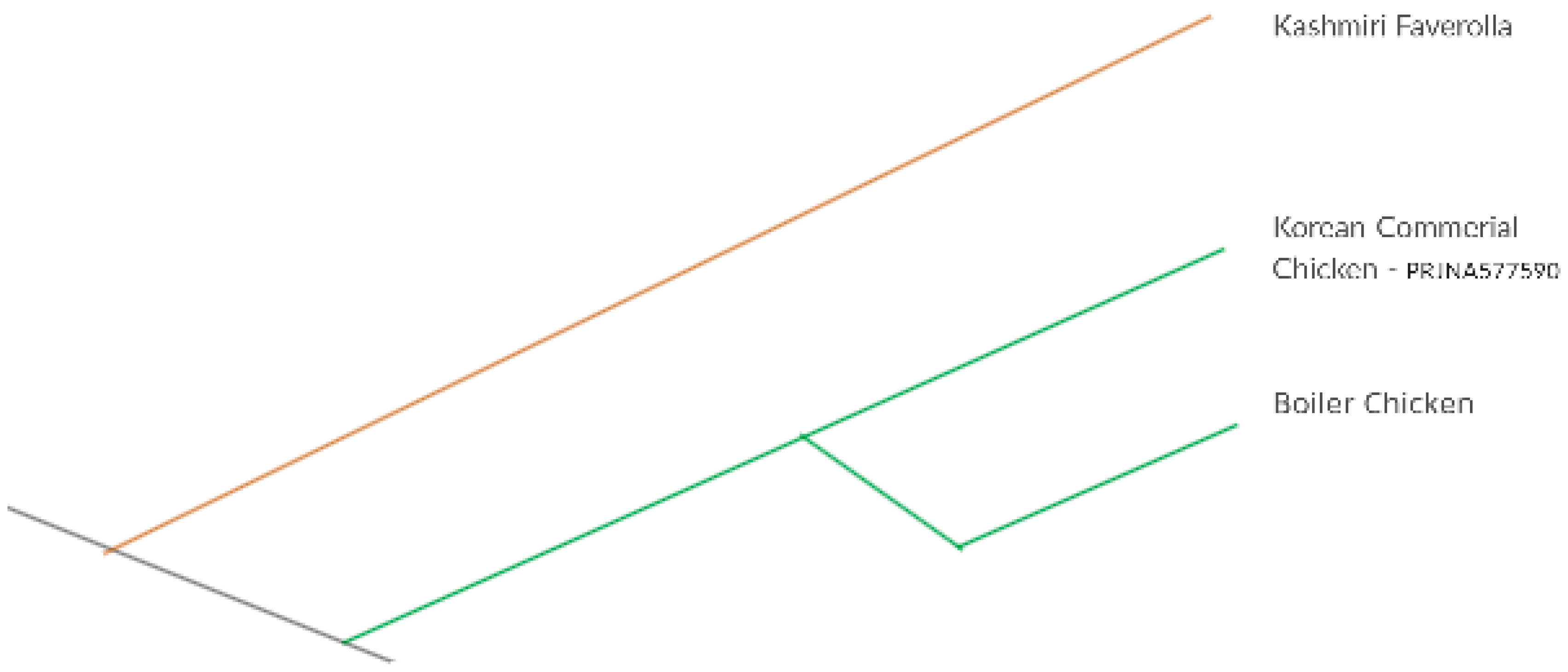
2.1. Experimental Birds and Sample Collection
2.2. Total RNA Isolation, cDNA Library Construction, and Sequencing
2.3. Quality Control, Aligning and Mapping Reads to the Genome
3. Results
3.1. Quality Control, Mapping, and Post Treatment
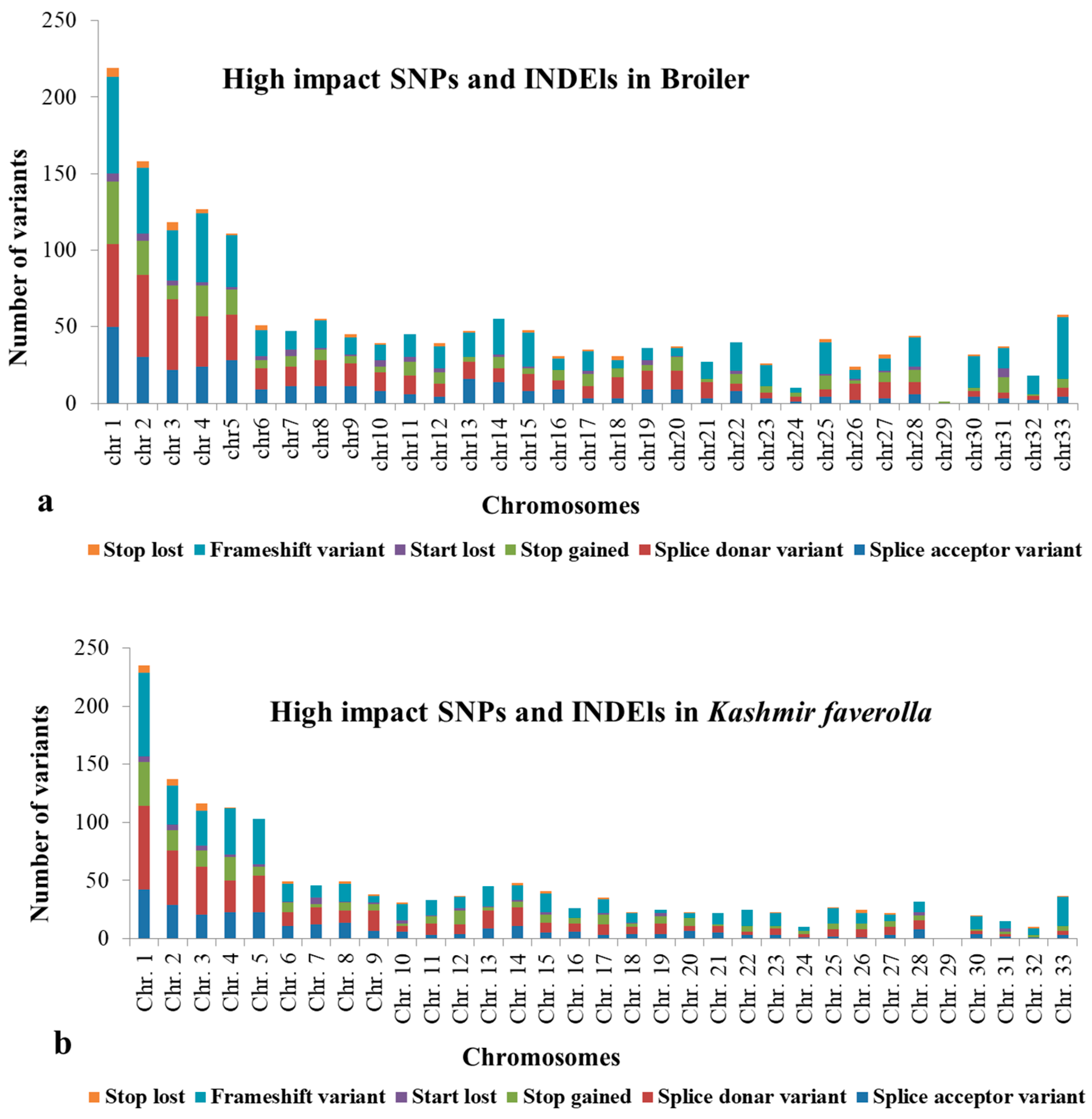
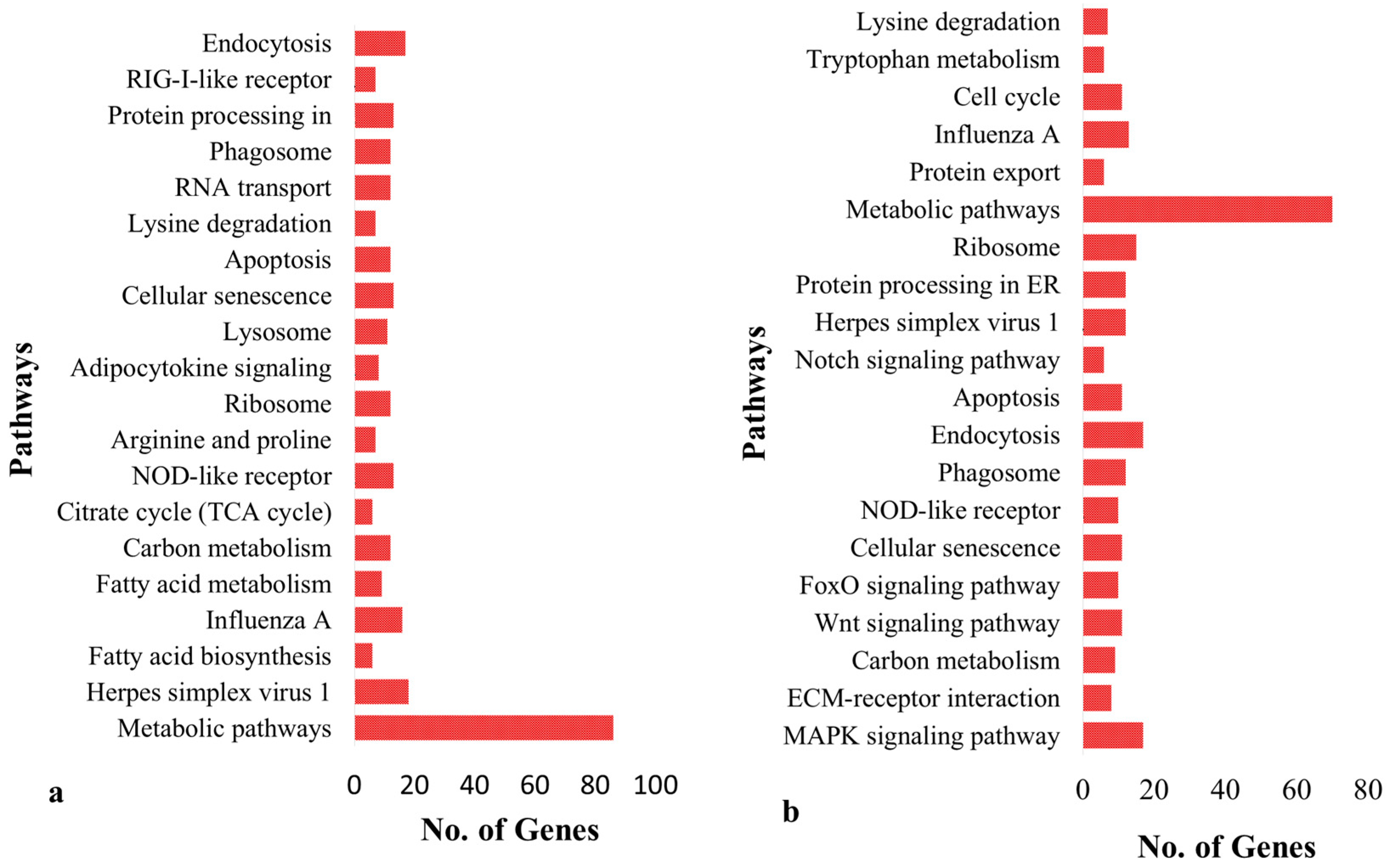
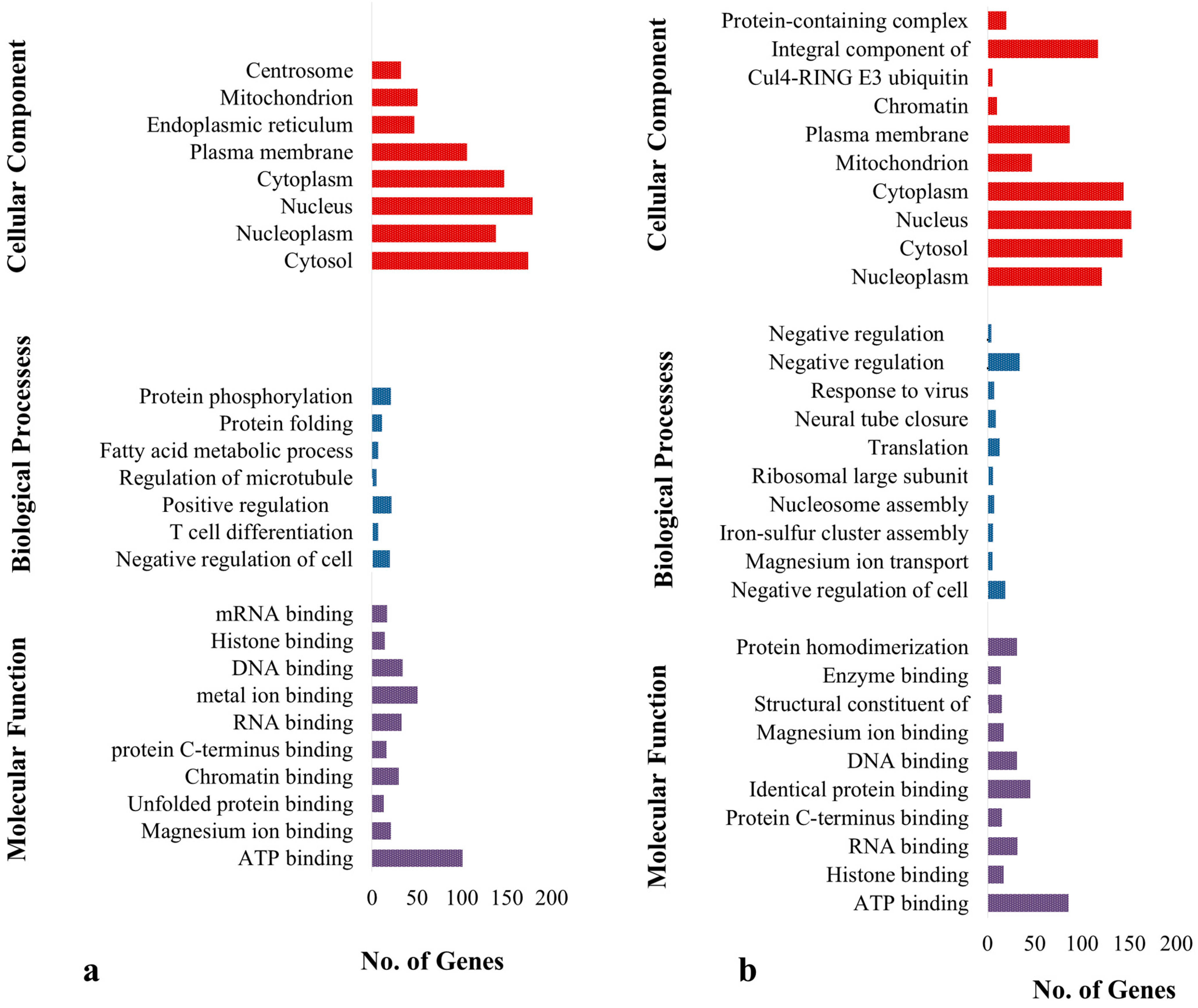
3.2. Analysis of Genes with SNPs and INDELs
3.3. Protein-Protein Interaction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DNA | Deoxyribonucleic acid |
| mRNA | Messenger RNA |
| TCF | Transcription Factor |
| CaMK | CaM-dependent proteins |
| SSRs | Simple sequence repeats |
| INDELs | insertions and deletions |
| BRCA | Breast cancer gene |
| SNP | Single nucleotide polymorphism |
| KEGG | Kyoto encyclopaedia of genes and genomes |
| IL | Interleukin |
| NLK | Nemo-like kinase |
| LRP | Low-density lipoprotein receptor-related protein |
| GO | Gene ontology |
| MLF1P | Myeloid leukemia factor |
References
- Dar, M.A.; Ahmad, S.M.; Bhat, B.A.; Dar, T.A.; ul Haq, Z.; Wani, B.A.; Shabir, N.; Kashoo, Z.A.; Shah, R.A.; Ganai, N.A.; et al. Comparative RNA-Seq analysis reveals insights in Salmonella disease resistance of chicken; and database development as resource for gene expression in poultry. Genomics 2022, 114, 110475. [Google Scholar] [CrossRef]
- Dar, M.A.; Ahmed, R.; Urwat, U.; Ahmad, S.M.; Dar, P.A.; Kushoo, Z.A.; Dar, T.A.; Mumtaz, P.T.; Bhat, S.A.; Amin, U.; et al. Expression kinetics of natural resistance associated macrophage protein (NRAMP) genes in Salmonella typhimurium-infected chicken. BMC Vet. Res. 2018, 14, 180. [Google Scholar] [CrossRef]
- Dar, M.A.; Mumtaz, P.T.; Bhat, S.A.; Taban, Q.; Khan, S.A.; Banday, T.; Ahmad, S.M. Immunopathogenesis of Salmonellosis. In New Insight into Brucella Infection and Foodborne Diseases; IntechOpen: London, UK, 2019. [Google Scholar]
- Jain, P.; Chowdhury, G.; Samajpati, S.; Basak, S.; Ganai, A.; Samanta, S.; Okamoto, K.; Mukhopadhyay, A.K.; Dutta, S. Characterization of non-typhoidal Salmonella isolates from children with acute gastroenteritis, Kolkata, India, during 2000–2016. Braz. J. Microbiol. 2020, 51, 613–627. [Google Scholar] [CrossRef]
- Scharff, R.L. Food Attribution and Economic Cost Estimates for Meat- and Poultry-Related Illnesses. J. Food Prot. 2020, 83, 959–967. [Google Scholar] [CrossRef] [PubMed]
- Dar, M.A.; Urwat, U.; Ahmad, S.M.; Ahmad, R.; Kashoo, Z.A.; Dar, T.A.; Bhat, S.A.; Mumtaz, P.T.; Shabir, N.; Shah, R.A.; et al. Gene expression and antibody response in chicken against Salmonella typhimurium challenge. Poult. Sci. 2019, 98, 2008–2013. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Nie, C.; Liu, Y.; Chen, Y.; Lv, X.; Wang, L.; Zhang, J.; Li, K.; Jia, Y.; Ban, L.; et al. A genome-wide association study explores the genetic determinism of host resistance to Salmonella pullorum infection in chickens. Genet. Sel. Evol. 2019, 51, 51. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Renu, S.; Patil, V.; Schrock, J.; Feliciano-Ruiz, N.; Selvaraj, R.; Renukaradhya, G.J. Immune response to Salmonella enteritidis infection in broilers immunized orally with chitosan-based Salmonella subunit nanoparticle vaccine. Front. Immunol. 2020, 19, 935. [Google Scholar] [CrossRef]
- Rodrigues, G.L.; Panzenhagen, P.; Ferrari, R.G.; Dos Santos, A.; Paschoalin, V.M.; Conte-Junior, C.A. Frequency of antimicrobial resistance genes in Salmonella from Brazil by in silico whole-genome sequencing analysis: An overview of the last four decades. Front. Microbiol. 2020, 11, 1864. [Google Scholar] [CrossRef]
- Manyi-Loh, C.; Mamphweli, S.; Meyer, E.; Okoh, A. Antibiotic use in agriculture and its consequential resistance in environmental sources: Potential public health implications. Molecules 2018, 30, 795. [Google Scholar] [CrossRef]
- Wang, M.; Qazi, I.H.; Wang, L.; Zhou, G.; Han, H. Salmonella virulence and immune escape. Microorganisms 2020, 8, 407. [Google Scholar] [CrossRef]
- Haraga, A.; Ohlson, M.B.; Miller, S.I. Salmonellae interplay with host cells. Nat. Rev. Microbiol. 2008, 6, 53–66. [Google Scholar] [CrossRef]
- Hansen-Wester, I.; Hensel, M. Salmonella pathogenicity islands encoding type III secretion systems. Microbes Infect. 2001, 3, 549–559. [Google Scholar] [CrossRef] [PubMed]
- Alpuche-Aranda, C.M.; Racoosin, E.L.; Swanson, J.A.; Miller, S.I. Salmonella stimulate macrophage macropinocytosis and persist within spacious phagosomes. J. Exp. Med. 1994, 179, 601–608. [Google Scholar] [CrossRef] [PubMed]
- Carrol, M.E.W.; Jackett, P.S.; Aber, V.R.; Lowrie, D.B. Phagolysosome formation, cyclic adenosine 3′: 5′-monophosphate and the fate of Salmonella typhimurium within mouse peritoneal macrophages. Microbiology 1979, 110, 421–429. [Google Scholar] [CrossRef] [PubMed]
- Buchmeier, N.A.; Heffron, F.R.E.D. Inhibition of macrophage phagosome-lysosome fusion by Salmonella typhimurium. Infect. Immun. 1991, 59, 2232–2238. [Google Scholar] [CrossRef] [PubMed]
- Rautenschlein, S.; Cheng, H.H.; Lamont, S.J. Host factors for disease resistance. Dis. Poult. 2020, 13, 79–108. [Google Scholar]
- Pal, A.; Chakravarty, A.K. Disease resistance for different livestock species. Genet. Breed. Dis. Resist. Livest. 2020, 2020, 271–296. [Google Scholar]
- Wani, H.; Darzi, M.M.; Kamil, S.A.; Wani, S.A.; Munshi, Z.H.; Shakoor, A.; Raja, T.A.; Shoukat, S.; Kashani, B.; Shah, A. Histological and histochemical studies on the reproductive tract of Kashmir faverolla chicken. J. Etnomology Zool. Stud. 2017, 5, 2256–2262. [Google Scholar]
- Iqbal, S.; Pampori, Z.A. Production potential and qualitative traits of indigenous chicken of Kashmir. Livest. Res. Rural. Dev. 2008, 20, 14. [Google Scholar]
- He, J.; Zhao, X.; Laroche, A.; Lu, Z.X.; Liu, H.; Li, Z. Genotyping-by-sequencing (GBS), an ultimate marker-assisted selection (MAS) tool to accelerate plant breeding. Front. Plant Sci. 2014, 30, 484. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Nawaz, M.A.; Shahid, M.Q.; Doğan, Y.; Comertpay, G.; Yıldız, M.; Hatipoğlu, R.; Ahmad, F.; Alsaleh, A.; Labhane, N.; et al. DNA molecular markers in plant breeding: Current status and recent advancements in genomic selection and genome editing. Biotechnol. Biotechnol. Equip. 2018, 32, 26185. [Google Scholar] [CrossRef]
- Jehl, F.; Degalez, F.; Bernard, M.; Lecerf, F.; Lagoutte, L.; Désert, C.; Coulée, M.; Bouchez, O.; Leroux, S.; Abasht, B.; et al. RNA-Seq Data for Reliable SNP Detection and Genotype Calling: Interest for Coding Variant Characterization and Cis-Regulation Analysis by Allele-Specific Expression in Livestock Species. Front. Genet. 2021, 12, 655707. [Google Scholar] [CrossRef]
- Ahmad, S.M.; Bhat, S.S.; Shafi, S.; Dar, M.A.; Saleem, A.; Haq, Z.; Farooq, N.; Nazir, J.; Bhat, B. Identification of key transcription factors and their functional role involved in Salmonella typhimurium infection in chicken using integrated transcriptome analysis and bioinformatics approach. BMC Genom. 2023, 24, 214. [Google Scholar] [CrossRef] [PubMed]
- Andrews, S. FastQC: A Quality Control Tool for High Throughput Sequence Data 2010; BibSonomy, Babraham Bioinformatics: Cambridgeshire, UK, 2018; Volume 17. [Google Scholar]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Poplin, R.; Chang, P.C.; Alexander, D.; Schwartz, S.; Colthurst, T.; Ku, A.; Newburger, D.; Dijamco, J.; Nguyen, N.; Afshar PTGross, S.S. A universal SNP and small-indel variant caller using deep neural networks. Nat. Biotechnol. 2018, 36, 983–987. [Google Scholar] [CrossRef] [PubMed]
- Toolkit, P. Broad Institute, GitHub Repository; Broad Institute: Cambridge, MA, USA, 2019. [Google Scholar]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R. The sequence alignment/map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef]
- Kanehisa, M.; Furumichi, M.; Sato, Y.; Ishiguro-Watanabe, M.; Tanabe, M. KEGG: Integrating viruses and cellular organisms. Nucleic Acids Res. 2021, 49, D545–D551. [Google Scholar] [CrossRef]
- Biswas, A.; Zhou, D.; Fiches, G.N.; Wu, Z.; Liu, X.; Ma, Q.; Zhao, W.; Zhu, J.; Santoso, N.G. Inhibition of polo-like kinase 1 (PLK1) facilitates reactivation of γ-herpesviruses and their elimination. PLoS Pathog. 2021, 23, e1009764. [Google Scholar] [CrossRef]
- Morand, S.; Stanbery, L.; Walter, A.; Rocconi, R.P.; Nemunaitis, J. BRCA1/2 Mutation Status Impact on Autophagy and Immune Response: Unheralded Target. JNCI Cancer Spectr. 2020, 4, pkaa077. [Google Scholar] [CrossRef]
- Irawan, C.; Atmakusumah, D.; Siregar, N.C.; Tean, T.B.; Kong, L.W.; Kiat, O.C.; Mansyur, M. Expression of biomarkers CXCR4, IL11-RA, TFF1, MLF1P in advanced breast cancer patients with bone metastatic: A diagnostic study. Acta Med. Indones 2016, 48, 261–268. [Google Scholar]
- Chambers, M.C.; Schneider, D.S. Balancing resistance and infection tolerance through metabolic means. Proc. Natl. Acad. Sci. USA 2012, 109, 13886–13887. [Google Scholar] [CrossRef]
- Chang, C.S.; Chen, C.F.; Berthouly-Salazar, C.; Chazara, O.; Lee, Y.P.; Chang, C.M.; Chang, K.H.; Bed’Hom, B.; Tixier-Boichard, M. A global analysis of molecular markers and phenotypic traits in local chicken breeds in Taiwan. Anim. Genet. 2012, 43, 172–182. [Google Scholar] [CrossRef]
- Khanyile, K.S.; Dzomba, E.F.; Muchadeyi, F.C. Population genetic structure, linkage disequilibrium and effective population size of conserved and extensively raised village chicken populations of Southern Africa. Front. Genet. 2015, 3, 13. [Google Scholar] [CrossRef]
- Girmay, G.; Pal, M.; Dessie, T.; Sissay, T.; Wubete, A. Evaluating the relative resistance of different poultry breeds to Salmonella typhimurium. Afr. J. Agric. Res. 2015, 10, 2928–2939. [Google Scholar]
- Wang, F.; Zhang, J.; Zhu, B.; Wang, J.; Wang, Q.; Zheng, M.; Wen, J.; Li, Q.; Zhao, G. Transcriptome analysis of the cecal tonsil of Jingxing yellow chickens revealed the mechanism of differential resistance to Salmonella. Genes 2019, 10, 979. [Google Scholar] [CrossRef]
- Lacharme-Lora, L.; Owen, S.V.; Blundell, R.; Canals, R.; Wenner, N.; Perez-Sepulveda, B.; Fong, W.Y.; Gilory, R.; Wigley, P.; Hinton, J.C. The use of chicken and insect infection models to assess the virulence of African Salmonella typhimurium ST313. PLoS Negl. Trop. Dis. 2019, 13, e0007540. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Tong, C.; Ma, S.; Zhou, L.; Zhao, L.; Zhao, X. Involvement of microRNAs in probiotics-induced reduction of the caecal inflammation by Salmonella typhimurium. Front. Immunol. 2017, 8, 704. [Google Scholar] [CrossRef] [PubMed]
- Li, S.Z.; Shu, Q.P.; Song, Y.; Zhang, H.H.; Liu, Y.; Jin, B.X.; Liuyu, T.Z.; Li, C.; Huang, X.C.; Du, R.L.; et al. Phosphorylation of MAVS/VISA by Nemo-like kinase (NLK) for degradation regulates the antiviral innate immune response. Nat. Commun. 2019, 10, 3233. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, J.K.; Kling, J.C.; Tran, T.T.; Blumenthal, A. Functions of the WNT signaling network in shaping host responses to infection. Front. Immunol. 2019, 10, 2521. [Google Scholar] [CrossRef] [PubMed]
- Monson, M.S.; Van Goor, A.G.; Ashwell, C.M.; Persia, M.E.; Rothschild, M.F.; Schmidt, C.J. Immunomodulatory effects of heat stress and lipopolysaccharide on the bursal transcriptome in two distinct chicken lines. BMC Genom. 2018, 19, 643. [Google Scholar] [CrossRef] [PubMed]
- Haseeb, M.; Pirzada, R.H.; Ain, Q.U.; Choi, S. Wnt Signaling in the Regulation of Immune Cell and Cancer Therapeutics. Cells 2019, 8, 1380. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Wang, W.; Wang, X. Roles of transcriptional factor 7 in production of inflammatory factors for lung diseases. J. Transl. Med. 2015, 13, 273. [Google Scholar] [CrossRef]
- Swafford, D.; Shanmugam, A.; Ranganathan, P.; Manoharan, I.; Hussein, M.S.; Patel, N.; Sifuentes, H.; Koni, P.A.; Prasad, P.D.; Thangaraju, M.; et al. The Wnt–β-Catenin–IL-10 Signaling Axis in Intestinal APCs Protects Mice from Colitis-Associated Colon Cancer in Response to Gut Microbiota. J. Immunol. 2020, 205, 2265–2275. [Google Scholar] [CrossRef]
- Beckendorf, J.; van den Hoogenhof, M.M.G.; Backs, J. Physiological and unappreciated roles of CaMKII in the heart. Basic Res. Cardiol. 2018, 113, 29. [Google Scholar] [CrossRef]
- Jati, S.; Sengupta, S.; Sen, M. Wnt5A-Mediated Actin Organization Regulates Host Response to Bacterial Pathogens and Non-Pathogens. Front. Immunol. 2021, 11, 628191. [Google Scholar] [CrossRef]
- Thompson, A.; Fulde, M.; Tedin, K. The metabolic pathways utilized by Salmonella typhimurium during infection of host cells. Environ. Microbiol. Rep. 2018, 10, 140–154. [Google Scholar] [CrossRef]
- Calenge, F.; Kaiser, P.; Vignal, A. Genetic control of resistance to salmonellosis and to Salmonella carrier-state in fowl: A review. Genet. Sel. Evol. 2010, 42, 11. [Google Scholar] [CrossRef]
- Izakovicova, H.L. Interleukin-18 in asthma and other allergies. Clin. Exp. Allergy 2003, 33, 1023–1025. [Google Scholar] [CrossRef]
- Imboden, M.; Nicod, L.; Nieters, A.; Glaus, E.; Matyas, G.; Bircher, A.J.; Ackermann-Liebrich, U.; Berger, W.; Probst-Hensch, N.M.; APALDIA Team. The common G-allele of interleukin-18 single-nucleotide polymorphism is a genetic risk factor for atopic asthma. The SAPALDIA Cohort Study. Clin. Exp. Allergy 2006, 36, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Bakr, N.M.; Awad, A.; Moustafa, E. Association of genetic variants in the interleukin-18 gene promoter with risk of hepatocellular carcinoma and metastasis in patients with hepatitis C virus infection. IUBMB Life 2018, 70, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Schabath, M.B.; Giuliano, A.R.; Thompson, Z.J.; Amankwah, E.K.; Gray, J.E.; Fenstermacher, D.A.; Jonathan, K.A.; Beg, A.A.; Haura, E.B. TNFRSF10B polymorphisms and haplotypes associated with increased risk of death in non-small cell lung cancer. Carcinogenesis 2013, 34, 2525–2530. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Chromosome | Reference Nucleotide | Mutated Nucleotide | SNP Nature | Impact |
|---|---|---|---|---|---|
| MADPRT1 | 1 | C | T | Stop gained | HIGH |
| PPARD | 26 | A | T | Splice donor variant and intron variant | HIGH |
| IL18 | 24 | G | A | Splice donor variant and intron variant | HIGH |
| IL18R1 | 1 | TCC | TCCC | frameshift variant | HIGH |
| TNFRSF10B | 22 | A | C | Splice acceptor variant and intron variant | HIGH |
| IL1R1 | 1 | T | A | Splice donor variant and Intron variant | HIGH |
| TNFAIP1 | 19 | G | A | Stop gained | HIGH |
| MMP28 | 19 | G | T | Splice donor variant and intron variant | HIGH |
| SLC9A9 | 9 | T | A | Splice donor variant and intron variant | HIGH |
| SLC5A10 | 14 | A | G | Stop lost and splice region variant | HIGH |
| SLC13A2 | 19 | T | C | Splice donor variant and intron variant | HIGH |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dar, M.A.; Bhat, B.; Nazir, J.; Saleem, A.; Manzoor, T.; Khan, M.; Haq, Z.; Bhat, S.S.; Ahmad, S.M. Identification of SNPs Related to Salmonella Resistance in Chickens Using RNA-Seq and Integrated Bioinformatics Approach. Genes 2023, 14, 1283. https://doi.org/10.3390/genes14061283
Dar MA, Bhat B, Nazir J, Saleem A, Manzoor T, Khan M, Haq Z, Bhat SS, Ahmad SM. Identification of SNPs Related to Salmonella Resistance in Chickens Using RNA-Seq and Integrated Bioinformatics Approach. Genes. 2023; 14(6):1283. https://doi.org/10.3390/genes14061283
Chicago/Turabian StyleDar, Mashooq Ahmad, Basharat Bhat, Junaid Nazir, Afnan Saleem, Tasaduq Manzoor, Mahak Khan, Zulfqarul Haq, Sahar Saleem Bhat, and Syed Mudasir Ahmad. 2023. "Identification of SNPs Related to Salmonella Resistance in Chickens Using RNA-Seq and Integrated Bioinformatics Approach" Genes 14, no. 6: 1283. https://doi.org/10.3390/genes14061283
APA StyleDar, M. A., Bhat, B., Nazir, J., Saleem, A., Manzoor, T., Khan, M., Haq, Z., Bhat, S. S., & Ahmad, S. M. (2023). Identification of SNPs Related to Salmonella Resistance in Chickens Using RNA-Seq and Integrated Bioinformatics Approach. Genes, 14(6), 1283. https://doi.org/10.3390/genes14061283