
Association between HSPA8 Gene Variants and Ischemic Stroke: A Pilot Study Providing Additional Evidence for the Role of Heat Shock Proteins in Disease Pathogenesis

 , , , , , , ,
, , , , , , ,  ,
,  and
and
Abstract
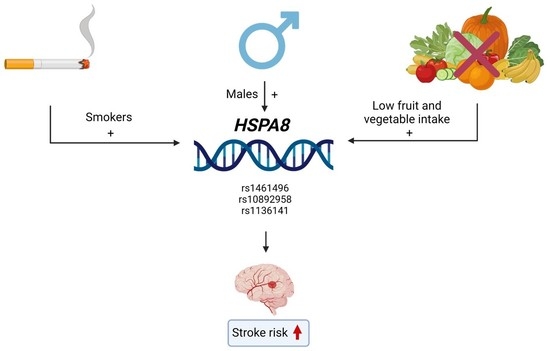
1. Introduction
2. Materials and Methods
2.1. Genetic Analysis
2.2. Statistical and Bioinformatic Analysis
- The expression quantitative trait loci (eQTLs) in the brain, whole blood, and blood vessels have been evaluated using the bioinformatic tool QTLbase (http://www.mulinlab.org/qtlbase/index.html (accessed on 21 February 2023)) [35].
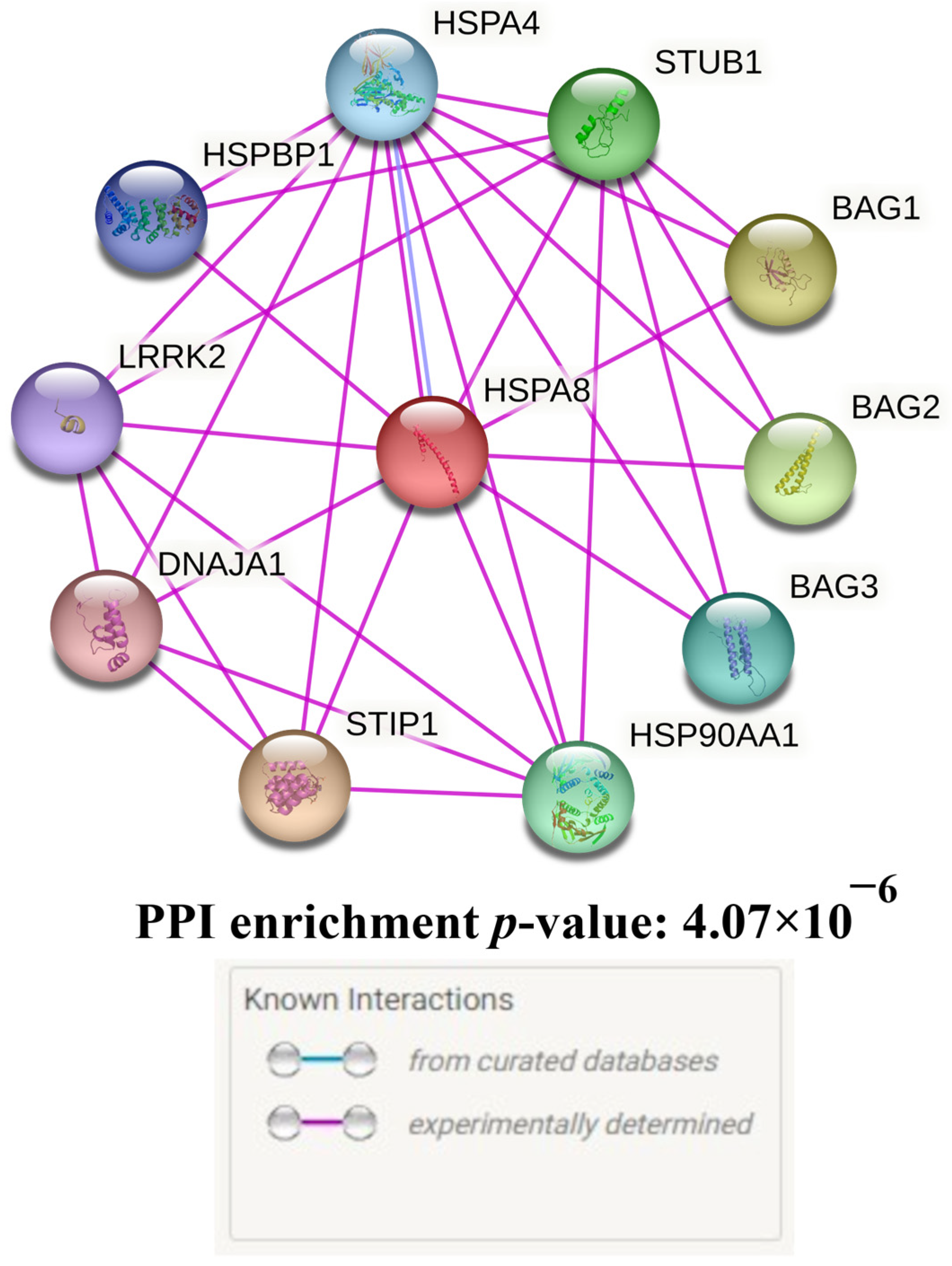
- The STRING database’s bioinformatic tools were utilised to analyse the main functional partners of HSPA8 (https://string-db.org/ (accessed on 21 February 2023)) [36]. Additionally, the STRING database was used to assess biological processes and molecular functions data describing interactions between HSPA8 and its functionally significant partner proteins. For the interpretation of interactions only experimentally confirmed data was used.
- The effect of HSPA8 SNPs on the binding of transcription factors (TFs) to DNA was assessed using the atSNP Function Prediction online tool (http://atsnp.biostat.wisc.edu/search (accessed on 21 February 2023)) [37]. Based on a positional weight matrix-based calculation of the impact of SNPs on how well TFs interact with DNA, certain TFs were added.
- The online Gene Ontology tool was used to conduct the subsequent study of the potential joint involvement of TFs linked with the reference and SNP alleles in overrepresented biological processes that are related to the mechanisms of IS (http://geneontology.org/ (accessed on 21 February 2023)) [38]. As functional groups, we employed biological processes governed by transcription factors connected to HSPA8 SNPs.
- HaploReg (v4.1), a bioinformatics tool (http://archive.broadinstitute.org/mammals/haploreg/haploreg.php (accessed on 20 February 2023)) was used to evaluate the relationships between HSPA8 SNPs and the following histone modifications that mark promoters and enhancers: acetylation of the lysine residues at positions 27 and 9 of the histone H3 protein, as well as mono-methylation at position 4 of the histone H3 protein (H3K4me1) and tri-methylation at position 4 of the histone H3 protein (H3K4me3). Additionally, this tool has been employed to examine the localization of SNPs in DNase hypersensitive areas, regulatory motif regions, and locations that bind to regulatory proteins [32].
- The interpretation of environment-associated correlates of HSPA8 polymorphism was done using the Comparative Toxicogenomics Database (CTD) resource at http://ctdbase.org (accessed on 24 February 2023) [39]. Based on data gathered from internationally published scientific studies, CTD offers the capability to investigate particular interactions between genes and chemicals in vertebrates and invertebrates. Using this method, bidirectional interactions comprising a single chemical and a single gene or protein were examined.
- The Cerebrovascular Disease Knowledge Portal (CDKP) is available at https://cd.hugeamp.org/ (accessed on 24 February 2023) was employed for a bioinformatic investigation of the relationships between HSPA8 SNPs and stroke-related traits, intermediate phenotypes, and risk factors for IS (such as blood pressure, heart rate, etc.) [40].
3. Results
3.1. Bioinformatic Analysis of the HSPA8 Gene
Protein–Protein Interactions
3.2. HSPA8 SNPs and the Ischemic Stroke Risk: An Analysis of Associations
3.3. Functional Annotation of HSPA8 SNPs
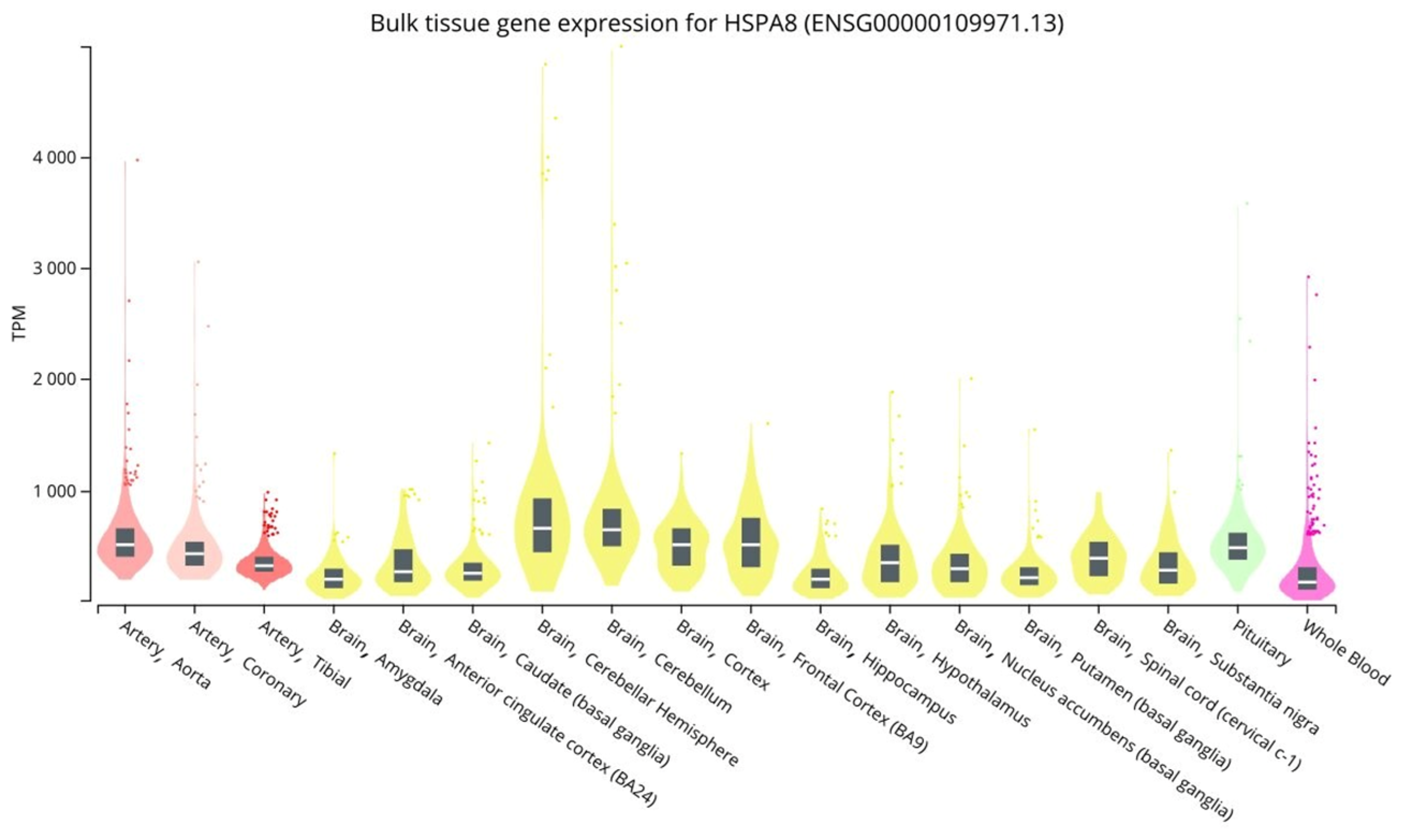
3.3.1. QTL-Effects
3.3.2. Histone Modifications
3.3.3. Analysis of Transcription Factors
3.3.4. Bioinformatic Analysis of the Associations of HSPA8 SNPs with IS-Related Phenotypes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- The Top 10 Causes of Death. Available online: https://www.who.int/news-room/fact-sheets/detail/the-top-10-causes-of-death (accessed on 30 March 2023).
- Orlacchio, A.; Bernardi, G. Research actuality in the genetics of stroke. Clin. Exp. Hypertens. 2006, 28, 191–197. [Google Scholar] [CrossRef]
- Malik, R.; Chauhan, G.; Traylor, M.; Sargurupremraj, M.; Okada, Y.; Mishra, A.; Rutten-Jacobs, L.; Giese, A.-K.; van der Laan, S.W.; Gretarsdottir, S.; et al. Multiancestry genome-wide association study of 520,000 subjects identifies 32 loci associated with stroke and stroke subtypes. Nat. Genet. 2018, 50, 524–537. [Google Scholar] [CrossRef]
- Debette, S.; Markus, H.S. Stroke Genetics: Discovery, Insight Into Mechanisms, and Clinical Perspectives. Circ. Res. 2022, 130, 1095–1111. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.; Barysheva, E.; Markov, A.; Belykh, A.; Koroleva, I.; Churkin, E.; Polonikov, A.; Ivanov, V.; Nazarenko, M. DNA Hypomethylation of the MPO Gene in Peripheral Blood Leukocytes Is Associated with Cerebral Stroke in the Acute Phase. J. Mol. Neurosci. 2021, 71, 1914–1932. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.Y.; Stetskaya, T.A.; Polonikov, A.V.; Ivanov, V.P. The relationship between polymorphism 640A > G of the CYBA gene with the risk of ischemic stroke in the population of the Central Russia. Zh. Nevrol. Psikhiatr. Im. S. S. Korsakova 2015, 115 Pt 2, 38. [Google Scholar] [CrossRef]
- Polonikov, A.; Bocharova, I.; Azarova, I.; Klyosova, E.; Bykanova, M.; Bushueva, O.; Polonikova, A.; Churnosov, M.; Solodilova, M. The Impact of Genetic Polymorphisms in Glutamate-Cysteine Ligase, a Key Enzyme of Glutathione Biosynthesis, on Ischemic Stroke Risk and Brain Infarct Size. Life 2022, 12, 602. [Google Scholar] [CrossRef]
- Polonikov, A.V.; Ushachev, D.V.; Ivanov, V.P.; Churnosov, M.I.; Freidin, M.B.; Ataman, A.V.; Harbuzova, V.Y.; Bykanova, M.A.; Bushueva, O.Y.; Solodilova, M.A. Altered erythrocyte membrane protein composition mirrors pleiotropic effects of hypertension susceptibility genes and disease pathogenesis. J. Hypertens. 2015, 33, 2265–2277. [Google Scholar] [CrossRef]
- Wang, M.; Brage, S.; Sharp, S.J.; Luo, S.; Yeung, S.L.A.; Kim, Y. Associations of genetic susceptibility and healthy lifestyle with incidence of coronary heart disease and stroke in individuals with hypertension. Eur. J. Prev. Cardiol. 2022, 29, 2101–2110. [Google Scholar] [CrossRef]
- Borén, J.; Chapman, M.J.; Krauss, R.M.; Packard, C.J.; Bentzon, J.F.; Binder, C.J.; Daemen, M.J.; Demer, L.L.; Hegele, R.A.; Nicholls, S.J.; et al. Low-density lipoproteins cause atherosclerotic cardiovascular disease: Pathophysiological, genetic, and therapeutic insights: A consensus statement from the European Atherosclerosis Society Consensus Panel. Eur. Heart J. 2020, 41, 2313–2330. [Google Scholar] [CrossRef]
- Sorokin, A.; Kotani, K.; Bushueva, O.; Taniguchi, N.; Lazarenko, V. The Cardio-Ankle Vascular Index and Ankle-Brachial Index in Young Russians. J. Atheroscler. Thromb. 2015, 22, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Nimjee, S.M.; Akhter, A.S.; Zakeri, A.; Herson, P.S. Sex differences in thrombosis as it affects acute ischemic stroke. Neurobiol. Dis. 2022, 165, 105647. [Google Scholar] [CrossRef] [PubMed]
- Sun, X.; Zhao, G.; Hua, C.; Ju, W.-N.; Jin, H. Molecular chaperones and hypoxic-ischemic encephalopathy. Neural Regen. Res. 2017, 12, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kim, J.W.; Yenari, M.A. Heat shock protein signaling in brain ischemia and injury. Neurosci. Lett. 2020, 715, 134642. [Google Scholar] [CrossRef]
- Doeppner, T.R.; Doehring, M.; Kaltwasser, B.; Majid, A.; Lin, F.; Bähr, M.; Kilic, E.; Hermann, D.M. Ischemic Post-Conditioning Induces Post-Stroke Neuroprotection via Hsp70-Mediated Proteasome Inhibition and Facilitates Neural Progenitor Cell Transplantation. Mol. Neurobiol. 2017, 54, 6061–6073. [Google Scholar] [CrossRef] [PubMed]
- Dukay, B.; Csoboz, B.; Tóth, M.E. Heat-Shock Proteins in Neuroinflammation. Front. Pharmacol. 2019, 10, 920. [Google Scholar] [CrossRef]
- Zhan, X.; Ander, B.P.; Liao, I.H.; Hansen, J.E.; Kim, C.; Clements, D.; Weisbart, R.H.; Nishimura, R.N.; Sharp, F.R. Recombinant Fv-Hsp70 Protein Mediates Neuroprotection After Focal Cerebral Ischemia in Rats. Stroke 2010, 41, 538–543. [Google Scholar] [CrossRef]
- Weiss, Y.G.; Maloyan, A.; Tazelaar, J.; Raj, N.; Deutschman, C.S. Adenoviral transfer of HSP-70 into pulmonary epithelium ameliorates experimental acute respiratory distress syndrome. J. Clin. Investig. 2002, 110, 801–806. [Google Scholar] [CrossRef]
- Stricher, F.; Macri, C.; Ruff, M.; Muller, S. HSPA8/HSC70 chaperone protein: Structure, Function, and Chemical Targeting. Autophagy 2013, 9, 1937–1954. [Google Scholar] [CrossRef]
- Yamamoto, Y.-H.; Kimura, T.; Momohara, S.; Takeuchi, M.; Tani, T.; Kimata, Y.; Kadokura, H.; Kohno, K. A Novel ER J-protein DNAJB12 Accelerates ER-associated Degradation of Membrane Proteins Including CFTR. Cell Struct. Funct. 2010, 35, 107–116. [Google Scholar] [CrossRef]
- Grove, D.E.; Fan, C.-Y.; Ren, H.Y.; Cyr, D.M. The endoplasmic reticulum–associated Hsp40 DNAJB12 and Hsc70 cooperate to facilitate RMA1 E3–dependent degradation of nascent CFTRΔF508. Mol. Biol. Cell 2011, 22, 301–314. [Google Scholar] [CrossRef]
- Sopha, P.; Kadokura, H.; Yamamoto, Y.-H.; Takeuchi, M.; Saito, M.; Tsuru, A.; Kohno, K. A Novel Mammalian ER-located J-protein, DNAJB14, Can Accelerate ERAD of Misfolded Membrane Proteins. Cell Struct. Funct. 2012, 37, 177–187. [Google Scholar] [CrossRef]
- Young, J.C.; Hoogenraad, N.J.; Hartl, F. Molecular Chaperones Hsp90 and Hsp70 Deliver Preproteins to the Mitochondrial Import Receptor Tom70. Cell 2003, 112, 41–50. [Google Scholar] [CrossRef]
- Matsumura, Y.; Sakai, J.; Skach, W.R. Endoplasmic Reticulum Protein Quality Control Is Determined by Cooperative Interactions between Hsp/c70 Protein and the CHIP E3 Ligase. J. Biol. Chem. 2013, 288, 31069–31079. [Google Scholar] [CrossRef] [PubMed]
- Vialykh, E.K.; A Solidolova, M.; Bushueva, O.; Bulgakova, I.V.; Polonikov, A. Catalase gene polymorphism is associated with increased risk of cerebral stroke in hypertensive patients. Zh. Nevrol. Psikhiatr. im. S. S. Korsakova 2012, 112, 3–7. [Google Scholar] [PubMed]
- Kobzeva, K.A.; Shilenok, I.V.; Belykh, A.E.; Gurtovoy, D.E.; Bobyleva, L.A.; Krapiva, A.B.; Stetskaya, T.A.; Bykanova, M.A.; Mezhenskaya, A.A.; Lysikova, E.A.; et al. C9orf16 (BBLN) gene, encoding a member of Hero proteins, is a novel marker in ischemic stroke risk. Res. RESULTS Biomed. 2022, 8, 278. [Google Scholar] [CrossRef]
- Who, J.; Consultation, F.E. Diet, nutrition and the prevention of chronic diseases. World Health Organ Tech. Rep. Ser. 2003, 916, 1–149. [Google Scholar]
- Bushueva, O.; Solodilova, M.; Ivanov, V.; Polonikov, A. Gender-specific protective effect of the −463G > A polymorphism of myeloperoxidase gene against the risk of essential hypertension in Russians. J. Am. Soc. Hypertens. 2015, 9, 902–906. [Google Scholar] [CrossRef]
- Bushueva, O.Y. Single nucleotide polymorphisms in genes encoding xenobiotic metabolizing enzymes are associated with predisposition to arterial hypertension. Res. Results Biomed. 2020, 6, 447–456. [Google Scholar] [CrossRef]
- Xu, Z.; Taylor, J.A. SNPinfo: Integrating GWAS and candidate gene information into functional SNP selection for genetic association studies. Nucleic Acids Res. 2009, 37, W600–W605. [Google Scholar] [CrossRef]
- Dong, S.; Boyle, A.P. Predicting functional variants in enhancer and promoter elements using RegulomeDB. Hum. Mutat. 2019, 40, 1292–1298. [Google Scholar] [CrossRef]
- Ward, L.D.; Kellis, M. HaploReg: A resource for exploring chromatin states, conservation, and regulatory motif alterations within sets of genetically linked variants. Nucleic Acids Res. 2012, 40, D930–D934. [Google Scholar] [CrossRef] [PubMed]
- Koressaar, T.; Remm, M. Enhancements and modifications of primer design program Primer3. Bioinformatics 2007, 23, 1289–1291. [Google Scholar] [CrossRef] [PubMed]
- Solé, X.; Guinó, E.; Valls, J.; Iniesta, R.; Moreno, V. SNPStats: A web tool for the analysis of association studies. Bioinformatics 2006, 22, 1928–1929. [Google Scholar] [CrossRef]
- Zheng, Z.; Huang, D.; Wang, J.; Zhao, K.; Zhou, Y.; Guo, Z.; Zhai, S.; Xu, H.; Cui, H.; Yao, H.; et al. QTLbase: An integrative resource for quantitative trait loci across multiple human molecular phenotypes. Nucleic Acids Res. 2020, 48, D983–D991. [Google Scholar] [CrossRef]
- Von Mering, C.; Jensen, L.J.; Snel, B.; Hooper, S.D.; Krupp, M.; Foglierini, M.; Jouffre, N.; Huynen, M.A.; Bork, P. STRING: Known and predicted protein-protein associations, integrated and transferred across organisms. Nucleic Acids Res. 2005, 33, D433–D437. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.; Hudson, R.; Harrison, C.; Craven, M.; Keleş, S. atSNP Search: A web resource for statistically evaluating influence of human genetic variation on transcription factor binding. Bioinformatics 2019, 35, 2657–2659. [Google Scholar] [CrossRef]
- The Gene Ontology Consortium. The Gene Ontology Resource: 20 years and still GOing strong. Nucleic Acids Res. 2019, 47, D330–D338. [Google Scholar] [CrossRef] [PubMed]
- Davis, A.P.; Grondin, C.J.; Johnson, R.J.; Sciaky, D.; Wiegers, J.; Wiegers, T.C.; Mattingly, C.J. Comparative Toxicogenomics Database (CTD): Update 2021. Nucleic Acids Res. 2021, 49, D1138–D1143. [Google Scholar] [CrossRef]
- Crawford, K.M.; Gallego-Fabrega, C.; Kourkoulis, C.; Miyares, L.; Marini, S.; Flannick, J.; Burtt, N.P.; Von Grotthuss, M.; Alexander, B.; Costanzo, M.C.; et al. Cerebrovascular Disease Knowledge Portal. Stroke 2018, 49, 470–475. [Google Scholar] [CrossRef]
- Rashedan, M. Characterization of the Localization of Heat-Shock Proteins, HSPA8 and HSPA5, in Mammalian Cells; California State University: Fullerton, CA, USA, 2013. [Google Scholar]
- Sirtori, R.; Riva, C.; Ferrarese, C.; Sala, G. HSPA8 knock-down induces the accumulation of neurodegenerative disorder-associated proteins. Neurosci. Lett. 2020, 736, 135272. [Google Scholar] [CrossRef]
- Bonam, S.R.; Ruff, M.; Muller, S. HSPA8/HSC70 in Immune Disorders: A Molecular Rheostat that Adjusts Chaperone-Mediated Autophagy Substrates. Cells 2019, 8, 849. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, A.; López-Cancio, E.; Pich, S.; Penalba, A.; Giralt, D.; García-Berrocoso, T.; Ferrer-Costa, C.; Gasull, T.; Hernández-Pérez, M.; Millan, M.; et al. Blood Biomarkers for the Early Diagnosis of Stroke: The Stroke-Chip Study. Stroke 2017, 48, 2419–2425. [Google Scholar] [CrossRef]
- Stankowski, J.N.; Zeiger, S.L.; Cohen, E.L.; DeFranco, D.B.; Cai, J.; McLaughlin, B. C-Terminus of Heat Shock Cognate 70 Interacting Protein Increases Following Stroke and Impairs Survival Against Acute Oxidative Stress. Antioxid. Redox Signal. 2011, 14, 1787–1801. [Google Scholar] [CrossRef] [PubMed]
- Dybdahl, B.; A Slørdahl, S.; Waage, A.; Kierulf, P.; Espevik, T.; Sundan, A. Myocardial ischaemia and the inflammatory response: Release of heat shock protein 70 after myocardial infarction. Heart 2005, 91, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Timofeeva, A.; Goryunova, L.; Khaspekov, G.; Kovalevskii, D.; Scamrov, A.; Bulkina, O.; Karpov, Y.; Talitskii, K.; Buza, V.; Britareva, V.; et al. Altered Gene Expression Pattern in Peripheral Blood Leukocytes from Patients with Arterial Hypertension. Ann. N. Y. Acad. Sci. 2006, 1091, 319–335. [Google Scholar] [CrossRef]
- Yu, Q.; Ju, P.; Kou, W.; Zhai, M.; Zeng, Y.; Maimaitiaili, N.; Shi, Y.; Xu, X.; Zhao, Y.; Jian, W.; et al. Macrophage-Specific NLRC5 Protects from Cardiac Remodeling Through Interaction With HSPA8. JACC Basic Transl. Sci. 2023, 8, 479–496. [Google Scholar] [CrossRef]
- Xu, H.; Jiang, J.; Chen, W.; Li, W.; Chen, Z. Vascular Macrophages in Atherosclerosis. J. Immunol. Res. 2019, 2019, 4354786. [Google Scholar] [CrossRef]
- Gerges, M.; Gerges, C.; Publig, M.; Skoro-Sajer, N.; Bonderman, D.; Frey, M.; Schwarzinger, I.; Lechner, K.; Seidl, V.; Alimohammadi, A.; et al. Chronic inflammation after splenectomy is a risk factor for increased thrombotic cardiovascular events. Eur. Heart J. 2017, 38 (Suppl. 1), A5706. [Google Scholar] [CrossRef]
- Guo, Y.-L.; Bai, R.; Chen, C.X.-J.; Liu, D.-Q.; Liu, Y.; Zhang, C.-Y.; Zen, K. Role of Junctional Adhesion Molecule-Like Protein in Mediating Monocyte Transendothelial Migration. Arter. Thromb. Vasc. Biol. 2009, 29, 75–83. [Google Scholar] [CrossRef]
- Liu, Y.; Jiang, S.; Yang, P.-Y.; Zhang, Y.-F.; Li, T.-J.; Rui, Y.-C. EF1A1/HSC70 Cooperatively Suppress Brain Endothelial Cell Apoptosis via Regulating JNK Activity. CNS Neurosci. Ther. 2016, 22, 836–844. [Google Scholar] [CrossRef]
- Chen, S.; Brown, I.R. Translocation of constitutively expressed heat shock protein Hsc70 to synapse-enriched areas of the cerebral cortex after hyperthermic stress. J. Neurosci. Res. 2007, 85, 402–409. [Google Scholar] [CrossRef]
- McLaughlin, B.; Hartnett, K.A.; Erhardt, J.A.; Legos, J.J.; White, R.F.; Barone, F.C.; Aizenman, E. Caspase 3 activation is essential for neuroprotection in preconditioning. Proc. Natl. Acad. Sci. USA 2003, 100, 715–720. [Google Scholar] [CrossRef] [PubMed]
- Garrido, C.; Schmitt, E.; Candé, C.; Vahsen, N.; Parcellier, A.; Kroemer, G. HSP27 and HSP70: Potentially oncogenic apoptosis inhibitors. Cell Cycle 2003, 2, 578–583. [Google Scholar] [CrossRef]
- Giffard, R.G.; Yenari, M.A. Many Mechanisms for Hsp70 Protection from Cerebral Ischemia. J. Neurosurg. Anesthesiol. 2004, 16, 53–61. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Zhang, L.; Kuang, Y.; Venkataramani, V.; Jin, F.; Hein, K.; Zafeiriou, M.P.; Lenz, C.; Moebius, W.; Kilic, E.; et al. Extracellular Vesicles Derived from Neural Progenitor Cells––A Preclinical Evaluation for Stroke Treatment in Mice. Transl. Stroke Res. 2021, 12, 185–203. [Google Scholar] [CrossRef]
- Chong, K.-Y.; Lai, C.-C.; Lille, S.; Chang, C.; Su, C.-Y. Stable Overexpression of the Constitutive Form of Heat Shock Protein 70 Confers Oxidative Protection. J. Mol. Cell. Cardiol. 1998, 30, 599–608. [Google Scholar] [CrossRef]
- Su, C.-Y.; Chong, K.-Y.; E Owen, O.; Dillmann, W.H.; Chang, C.; Lai, C.-C. Constitutive and Inducible hsp70s are Involved in Oxidative Resistance Evoked by Heat Shock or Ethanol. J. Mol. Cell. Cardiol. 1998, 30, 587–598. [Google Scholar] [CrossRef]
- Canton, M.; Menabo, R.; Carpi, A.; Di Lisa, F. P253Oxidative stress causes the release of a specific subset of proteins from viable cardiac myocytes. Cardiovasc. Res. 2014, 103 (Suppl. 1), S45. [Google Scholar] [CrossRef]
- Allende, M.; Molina, E.; Guruceaga, E.; Tamayo, I.; González-Porras, J.R.; Gonzalez-López, T.J.; Toledo, E.; Rabal, O.; Ugarte, A.; Roldán, V.; et al. Hsp70 protects from stroke in atrial fibrillation patients by preventing thrombosis without increased bleeding risk. Cardiovasc. Res. 2016, 110, 309–318. [Google Scholar] [CrossRef]
- Silander, K.; Alanne, M.; Kristiansson, K.; Saarela, O.; Ripatti, S.; Auro, K.; Karvanen, J.; Kulathinal, S.; Niemelä, M.; Ellonen, P.; et al. Gender Differences in Genetic Risk Profiles for Cardiovascular Disease. PLoS ONE 2008, 3, e3615. [Google Scholar] [CrossRef]
- Bushueva, O.; Solodilova, M.; Churnosov, M.; Ivanov, V.; Polonikov, A. The Flavin-Containing Monooxygenase 3 Gene and Essential Hypertension: The Joint Effect of Polymorphism E158K and Cigarette Smoking on Disease Susceptibility. Int. J. Hypertens. 2014, 2014, 712169. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.Y.; Bulgakova, I.V.; Ivanov, V.P.; Polonikov, A.V. Association of Flavin Monooxygenase Gene E158K Polymorphism with Chronic Heart Disease Risk. Bull. Exp. Biol. Med. 2015, 159, 776–778. [Google Scholar] [CrossRef]
- Bushueva, O.; A Stetskaya, T.; Korogodina, G.V.; Ivanov, V.P.; Polonikov, A. Gender-specific differences of endothelial nitric oxide synthase e298d polymorphism and the risk of stroke. Clin. Med. (Russ. J.) 2015, 93, 34–40. [Google Scholar]
- Stetskaia, T.A.; Bushueva, O.I.; Bulgakova, I.V.; Vialykh, E.K.; Shuteeva, T.V.; Biriukov, A.E.; Ivanov, V.P.; Polonikov, A.V. Association of T174M polymorphism of the angiotensinogen gene with the higher risk of cerebral stroke in women. Ter. Arkhiv 2014, 86, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Jennen, D.G.J.; Magkoufopoulou, C.; Ketelslegers, H.B.; van Herwijnen, M.H.M.; Kleinjans, J.C.S.; van Delft, J.H.M. Comparison of HepG2 and HepaRG by Whole-Genome Gene Expression Analysis for the Purpose of Chemical Hazard Identification. Toxicol. Sci. 2010, 115, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Scafoglio, C.; Ambrosino, C.; Cicatiello, L.; Altucci, L.; Ardovino, M.; Bontempo, P.; Medici, N.; Molinari, A.M.; Nebbioso, A.; Facchiano, A.; et al. Comparative gene expression profiling reveals partially overlapping but distinct genomic actions of different antiestrogens in human breast cancer cells. J. Cell. Biochem. 2006, 98, 1163–1184. [Google Scholar] [CrossRef]
- Wang, D.-Y.; Fulthorpe, R.; Liss, S.N.; Edwards, E.A. Identification of Estrogen-Responsive Genes by Complementary Deoxyribonucleic Acid Microarray and Characterization of a Novel Early Estrogen-Induced Gene: EEIG1. Mol. Endocrinol. 2004, 18, 402–411. [Google Scholar] [CrossRef]
- Krebs, C.J.; Jarvis, E.D.; Pfaff, D.W. The 70-kDa heat shock cognate protein (Hsc73) gene is enhanced by ovarian hormones in the ventromedial hypothalamus. Proc. Natl. Acad. Sci. USA 1999, 96, 1686–1691. [Google Scholar] [CrossRef]
- Kwekel, J.C.; Burgoon, L.D.; Burt, J.W.; Harkema, J.R.; Zacharewski, T.R. A cross-species analysis of the rodent uterotrophic program: Elucidation of conserved responses and targets of estrogen signaling. Physiol. Genom. 2005, 23, 327–342. [Google Scholar] [CrossRef]
- De Oliveira, A.A.; Faustino, J.; Nunes, K.P. Transcriptomic Data Analysis Reveals Sex-Related Differences in the Interplay Between Toll-Like Receptor 4 and Heat-Shock Protein 70 in the Aorta of Type 2 Diabetic Donors. Hypertension 2019, 74 (Suppl. 1), AP190. [Google Scholar] [CrossRef]
- Polonikov, A.V.; Samgina, T.A.; Nazarenko, P.M.; Bushueva, O.Y.; Ivanov, V.P. Alcohol Consumption and Cigarette Smoking are Important Modifiers of the Association Between Acute Pancreatitis and the PRSS1-PRSS2 Locus in Men. Pancreas 2017, 46, 230–236. [Google Scholar] [CrossRef] [PubMed]
- Bushueva, O.Y.; Ivanov, V.P.; Ryzhaeva, V.N.; Ponomarenko, I.V.; I Churnosov, M.; Polonikov, A.V. Association of the -844G>A polymorphism in the catalase gene with the increased risk of essential hypertension in smokers. Ter. Arkhiv 2016, 88, 50–54. [Google Scholar] [CrossRef] [PubMed]
- Scicchitano, P.; Cortese, F.; Gesualdo, M.; De Palo, M.; Massari, F.; Giordano, P.; Ciccone, M.M. The role of endothelial dysfunction and oxidative stress in cerebrovascular diseases. Free. Radic. Res. 2019, 53, 579–595. [Google Scholar] [CrossRef]
- Bacchetti, T.; Turco, I.; Urbano, A.; Morresi, C.; Ferretti, G. Relationship of fruit and vegetable intake to dietary antioxidant capacity and markers of oxidative stress: A sex-related study. Nutrition 2019, 61, 164–172. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, A.V.; Kotani, K.; Bushueva, O.Y.; Polonikov, A.V. Antioxidant-related gene polymorphisms associated with the cardio-ankle vascular index in young Russians. Cardiol. Young 2016, 26, 677–682. [Google Scholar] [CrossRef]
- Sorokin, A.V.; Kotani, K.; Bushueva, O.Y. Association of matrix metalloproteinase 3 and γ-glutamyltransferase 1 gene polymorphisms with the cardio-ankle vascular index in young Russians. Cardiol. Young 2016, 26, 1238–1240. [Google Scholar] [CrossRef]
- Polonikov, A.; Vialykh, E.; Vasil’eva, O.; Bulgakova, I.; Bushueva, O.; Illig, T.; Solodilova, M. Genetic Variation in Glutathione S-Transferase Genes and Risk of Nonfatal Cerebral Stroke in Patients Suffering from Essential Hypertension. J. Mol. Neurosci. 2012, 47, 511–513. [Google Scholar] [CrossRef]
- Vasil’eva Yu, I.; Bushueva, O.; Zhabin, S.; Ivanov, S.V.; Polonikov, A. Smoking as a trigger factor in the development of diabetic angiopathy of lower extremities in men with methylenetetrahydrofolatereductase 677tt genotype. Clin. Med. (Russ. J.) 2015, 93, 45–49. [Google Scholar]
- Iskandar, A.R.; Mathis, C.; Schlage, W.K.; Frentzel, S.; Leroy, P.; Xiang, Y.; Sewer, A.; Majeed, S.; Ortega-Torres, L.; Johne, S.; et al. A systems toxicology approach for comparative assessment: Biological impact of an aerosol from a candidate modified-risk tobacco product and cigarette smoke on human organotypic bronchial epithelial cultures. Toxicol. Vitr. 2017, 39, 29–51. [Google Scholar] [CrossRef]
- Sheridan, J. Partnership between the Aryl Hydrocarbon Receptor (AHR) and RELB Regulates Cigarette Smoke-Induced Cyclooxygenase-2 (COX-2) Expression; McGill University: Montréal, QC, Canada, 2014. [Google Scholar]
- Bin-Jumah, M.N.; Nadeem, M.S.; Gilani, S.J.; Mubeen, B.; Ullah, I.; Alzarea, S.I.; Ghoneim, M.M.; Alshehri, S.; Al-Abbasi, F.A.; Kazmi, I. Lycopene: A Natural Arsenal in the War against Oxidative Stress and Cardiovascular Diseases. Antioxidants 2022, 11, 232. [Google Scholar] [CrossRef]
- Ciumărnean, L.; Milaciu, M.V.; Runcan, O.; Vesa, S.C.; Răchișan, A.L.; Negrean, V.; Perné, M.-G.; Donca, V.I.; Alexescu, T.-G.; Para, I.; et al. The effects of favonoids in cardiovascular diseases. Molecules 2020, 25, 4320. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.B.; A Niaz, M.; Agarwal, P.; Begom, R.; Rastogi, S.S. Effect of Antioxidant-Rich Foods on Plasma Ascorbic Acid, Cardiac Enzyme, and Lipid Peroxide Levels in Patients Hospitalized with Acute Myocardial Infarction. J. Am. Diet. Assoc. 1995, 95, 775–780. [Google Scholar] [CrossRef] [PubMed]
- Ishige, K.; Schubert, D.; Sagara, Y. Flavonoids protect neuronal cells from oxidative stress by three distinct mechanisms. Free. Radic. Biol. Med. 2001, 30, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Spada, P.D.; Dani, C.; Bortolini, G.V.; Funchal, C.; Henriques, J.A.; Salvador, M. Frozen Fruit Pulp of Euterpe oleraceae Mart. (Acai) Prevents Hydrogen Peroxide-Induced Damage in the Cerebral Cortex, Cerebellum, and Hippocampus of Rats. J. Med. Food 2009, 12, 1084–1088. [Google Scholar] [CrossRef] [PubMed]
- Scalbert, A.; Williamson, G. Dietary Intake and Bioavailability of Polyphenols. J. Nutr. 2000, 130 (Suppl. 8), 2073S–2085S. [Google Scholar] [CrossRef]
- Chadwick, W.; Zhou, Y.; Park, S.-S.; Wang, L.; Mitchell, N.; Stone, M.D.; Becker, K.G.; Martin, B.; Maudsley, S. Minimal Peroxide Exposure of Neuronal Cells Induces Multifaceted Adaptive Responses. PLoS ONE 2010, 5, e14352. [Google Scholar] [CrossRef] [PubMed]
- Law, C.-H.; Li, J.-M.; Chou, H.-C.; Chen, Y.-H.; Chan, H.-L. Hyaluronic acid-dependent protection in H9C2 cardiomyocytes: A cell model of heart ischemia–reperfusion injury and treatment. Toxicology 2013, 303, 54–71. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Baseline and Clinical Characteristics | IS Patients (N = 888) | Controls (N = 1251) | p-Value | |
|---|---|---|---|---|
| Age, Me [Q1; Q3] | 62 [55; 69] | 58 [53; 66] | <0.001 | |
| Gender | Males, N (%) | 481 (54.2%) | 577 (46.1%) | <0.001 |
| Females, N (%) | 407 (45.8%) | 674 (53.9%) | ||
| Smoking | Yes, N (%) | 425 (47.9%) | 331 (26.5%) | <0.001 |
| No, N (%) | 463 (52.1%) | 920 (73.5%) | ||
| Hypodynamia | Yes, N (%) | 332 (39.34%) | ND | |
| No, N (%) | 512 (60.66%) | |||
| Low fruit/vegetable consumption | Yes, N (%) | 449 (53.20%) | ND | |
| No, N (%) | 395 (46.80%) | |||
| Type 2 diabetes mellitus | Yes, N (%) | 103 (11,6%) | - | |
| No, N (%) | 740 (83,3%) | - | ||
| ND, N (%) | 45 (5,1%) | - | ||
| Body mass index, Me [Q1; Q3] | 23 [22; 26] (N = 567) | - | ||
| Family history of cerebrovascular diseases | Yes, N (%) | 296 (35.20%) | ND | |
| No, N (%) | 545 (64.80%) | ND | ||
| Age at onset of stroke, Me [Q1; Q3] | 61 [54; 69] (N = 862) | - | ||
| Number of strokes including event in question | 1, N (%) | 766 (88.86%) | - | |
| 2, N (%) | 85 (9.86%) | - | ||
| 3, N (%) | 11 (1.28%) | - | ||
| Stroke localization | Right/left middle cerebral artery basin, N (%) | 720 (83.82%) | - | |
| Vertebrobasilar basin, N (%) | 139 (16.18%) | - | ||
| Area of lesion in stroke, mm2, Me [Q1; Q3] | 105.00 [28; 468] (N = 841) | - | ||
| Total cholesterol, mmol/L, Me [Q1; Q3] | 5.2 [4.4; 5.8] (N = 583) | ND | ||
| Triglycerides, mmol/L, Me [Q1; Q3] | 1.3 [1.1; 1.8] (N = 577) | ND | ||
| Glucose level, mmol/L, Me [Q1; Q3] | 4.7 [4.3; 5.5] (N = 849) | ND | ||
| Prothrombin time, seconds, Me [Q1; Q3] | 10.79 [10.14; 11.70] (N = 839) | ND | ||
| International normalized ratio, Me [Q1; Q3] | 1 [0.94; 1.09] (N = 573) | ND | ||
| Activated partial thromboplastin time, seconds, Me [Q1; Q3] | 32.7 [29; 37] (N = 576) | ND | ||
| Genetic Variant | Effect Allele | Other Allele | N | OR [95% CI] 1 | p2 (Pbonf) |
|---|---|---|---|---|---|
| Entire group | |||||
| rs1461496 | A | G | 2132 | 1.00 [0.88–1.15] | 0.95 |
| rs10892958 | G | G | 2138 | 1.16 [0.99–1.35] | 0.06 |
| rs1136141 | A | G | 2024 | 1.09 [0.90–1.30] | 0.38 |
| Males | |||||
| rs1461496 | A | G | 1065 | 1.01 [0.85; 1.21] | 0.9 |
| rs10892958 | G | G | 1057 | 1.30 [1.05; 1.61] | 0.01 |
| rs1136141 | A | G | 999 | 1.08 [0.84; 1.40] | 0.55 |
| Females | |||||
| rs1461496 | A | G | 1076 | 1.05 [0.87; 1.26] | 0.63 |
| rs10892958 | G | G | 1081 | 1.08 [0.87; 1.33] | 0.49 |
| rs1136141 | A | G | 1025 | 1.16 [0.91; 1.47] | 0.24 |
| Nonsmokers (f−) | |||||
| rs1461496 | A | G | 1379 | 1.10 [0.93; 1.31] | 0.24 |
| rs10892958 | G | G | 1383 | 1.07 [0.88; 1.30] | 0.51 |
| rs1136141 | A | G | 1306 | 0.87 [0.68; 1.10] | 0.23 |
| Smokers (f+) | |||||
| rs1461496 | A | G | 753 | 0.86 [0.70; 1.07] | 0.18 |
| rs10892958 | G | G | 755 | 1.37 [1.07; 1.77] | 0.01 |
| rs1136141 | A | G | 718 | 1.68 [1.23; 2.28] | 7.0 × 10−4 |
| Normal fruit and vegetable intake (f−) | |||||
| rs1461496 | A | G | 1639 | 1.02 [0.86; 1.20] | 0.86 (bonf1.0) |
| rs10892958 | G | G | 1645 | 1.03 [0.84; 1.25] | 0.78 (bonf1.0) |
| rs1136141 | A | G | 1559 | 0.91 [0.72; 1.16] | 0.45 (bonf 0.9) |
| Low fruit and vegetable intake (f+) | |||||
| rs1461496 | A | G | 1694 | 1.07 [0.91; 1.25] | 0.42 (bonf0.84) |
| rs10892958 | G | G | 1699 | 1.36 [1.14; 1.63] | 9.0 × 10−4 (bonf0.002) |
| rs1136141 | A | G | 1608 | 1.29 [1.05; 1.60] | 0.02 (bonf0.04) |
| SNP | Trait | Effect Allele | Tissue | Effect Size (Beta) | PVAL | FDR |
|---|---|---|---|---|---|---|
| rs10892958 | HSPA8 | G | Brain-Hippocampus | −0.44 | 1.9 × 10−7 | 5.8 × 10−5 |
| rs1136141 | HSPA8 | A | Brain-Hippocampus | −0.12 | 3.8 × 10−5 | 0.006 |
| CLMP | A | Brain-Hippocampus | 0.13 | 4.8 × 10−5 | 0.008 |
| SNP (Ref/Alt Allele) | Tissues Marks | Brain | Blood | ||||||
|---|---|---|---|---|---|---|---|---|---|
| (1) | (2) | (3) | (4) | (5) | (6) | (7) | (8) | ||
| rs10892958 (C/G) | H3K4me1 | No | No | No | E | No | No | E | E |
| H3K4me3 | P | P | P | P | P | P | P | P | |
| H3K27ac | E | E | E | E | E | E | E | E | |
| H3K9ac | No | P | P | P | P | P | P | P | |
| DNase | No | No | No | No | No | No | No | DNase | |
| rs1136141 (G/A) | H3K4me1 | No | No | No | No | No | No | No | E |
| H3K4me3 | P | P | P | P | P | P | P | P | |
| H3K27ac | E | E | E | E | E | E | E | E | |
| H3K9ac | No | P | P | P | P | P | P | P | |
| DNase | No | No | No | No | No | No | No | DNase | |
| No. | SNP | Phenotype | p-Value | Beta (OR) | Sample Size |
|---|---|---|---|---|---|
| 1. | rs1136141 (G/A) | Systolic blood pressure | 0.008 | Beta▲0.0056 | 1,325,890 |
| 2. | Heart rate | 0.01 | Beta▲0.007 | 484,178 | |
| 3. | Peripheral artery disease in ever-smokers | 0.04 | OR▲1.0639 | 28,235 | |
| 4. | TOAST, other-determined | 0.03 | OR▲2.4103 | 9277 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kobzeva, K.A.; Soldatova, M.O.; Stetskaya, T.A.; Soldatov, V.O.; Deykin, A.V.; Freidin, M.B.; Bykanova, M.A.; Churnosov, M.I.; Polonikov, A.V.; Bushueva, O.Y. Association between HSPA8 Gene Variants and Ischemic Stroke: A Pilot Study Providing Additional Evidence for the Role of Heat Shock Proteins in Disease Pathogenesis. Genes 2023, 14, 1171. https://doi.org/10.3390/genes14061171
Kobzeva KA, Soldatova MO, Stetskaya TA, Soldatov VO, Deykin AV, Freidin MB, Bykanova MA, Churnosov MI, Polonikov AV, Bushueva OY. Association between HSPA8 Gene Variants and Ischemic Stroke: A Pilot Study Providing Additional Evidence for the Role of Heat Shock Proteins in Disease Pathogenesis. Genes. 2023; 14(6):1171. https://doi.org/10.3390/genes14061171
Chicago/Turabian StyleKobzeva, Ksenia A., Maria O. Soldatova, Tatiana A. Stetskaya, Vladislav O. Soldatov, Alexey V. Deykin, Maxim B. Freidin, Marina A. Bykanova, Mikhail I. Churnosov, Alexey V. Polonikov, and Olga Y. Bushueva. 2023. "Association between HSPA8 Gene Variants and Ischemic Stroke: A Pilot Study Providing Additional Evidence for the Role of Heat Shock Proteins in Disease Pathogenesis" Genes 14, no. 6: 1171. https://doi.org/10.3390/genes14061171
APA StyleKobzeva, K. A., Soldatova, M. O., Stetskaya, T. A., Soldatov, V. O., Deykin, A. V., Freidin, M. B., Bykanova, M. A., Churnosov, M. I., Polonikov, A. V., & Bushueva, O. Y. (2023). Association between HSPA8 Gene Variants and Ischemic Stroke: A Pilot Study Providing Additional Evidence for the Role of Heat Shock Proteins in Disease Pathogenesis. Genes, 14(6), 1171. https://doi.org/10.3390/genes14061171