
Alternative Evolutionary Pathways in Paspalum Involving Allotetraploidy, Sexuality, and Varied Mating Systems


 ,
,  , , ,
, , , 


Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Collections
2.2. Ploidy Level Estimation
2.3. Reproductive Mode
2.4. Reproductive Pathway Efficiency
2.5. Mating System and Seed Fertility
2.6. Statistical Analysis
3. Results
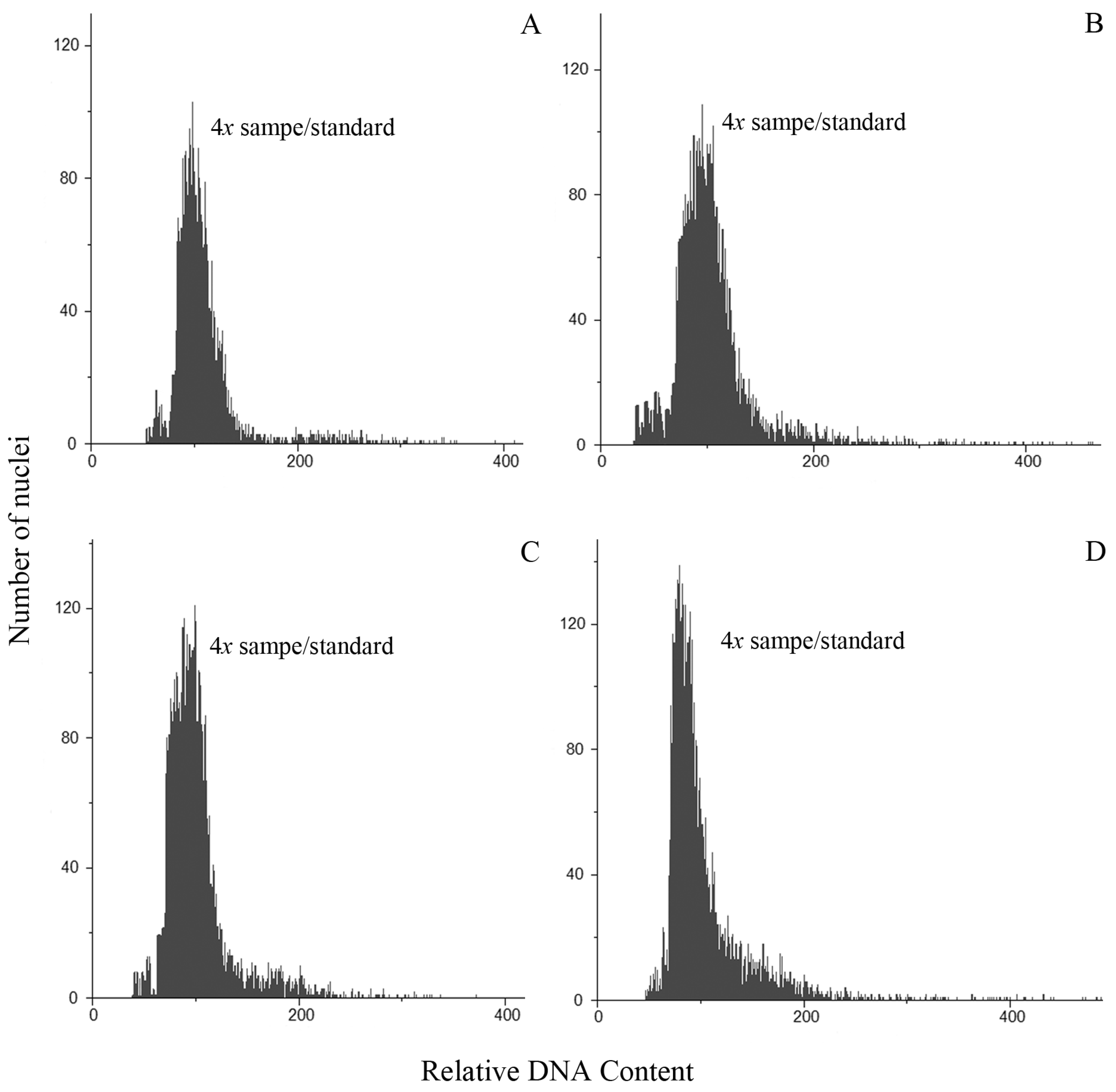
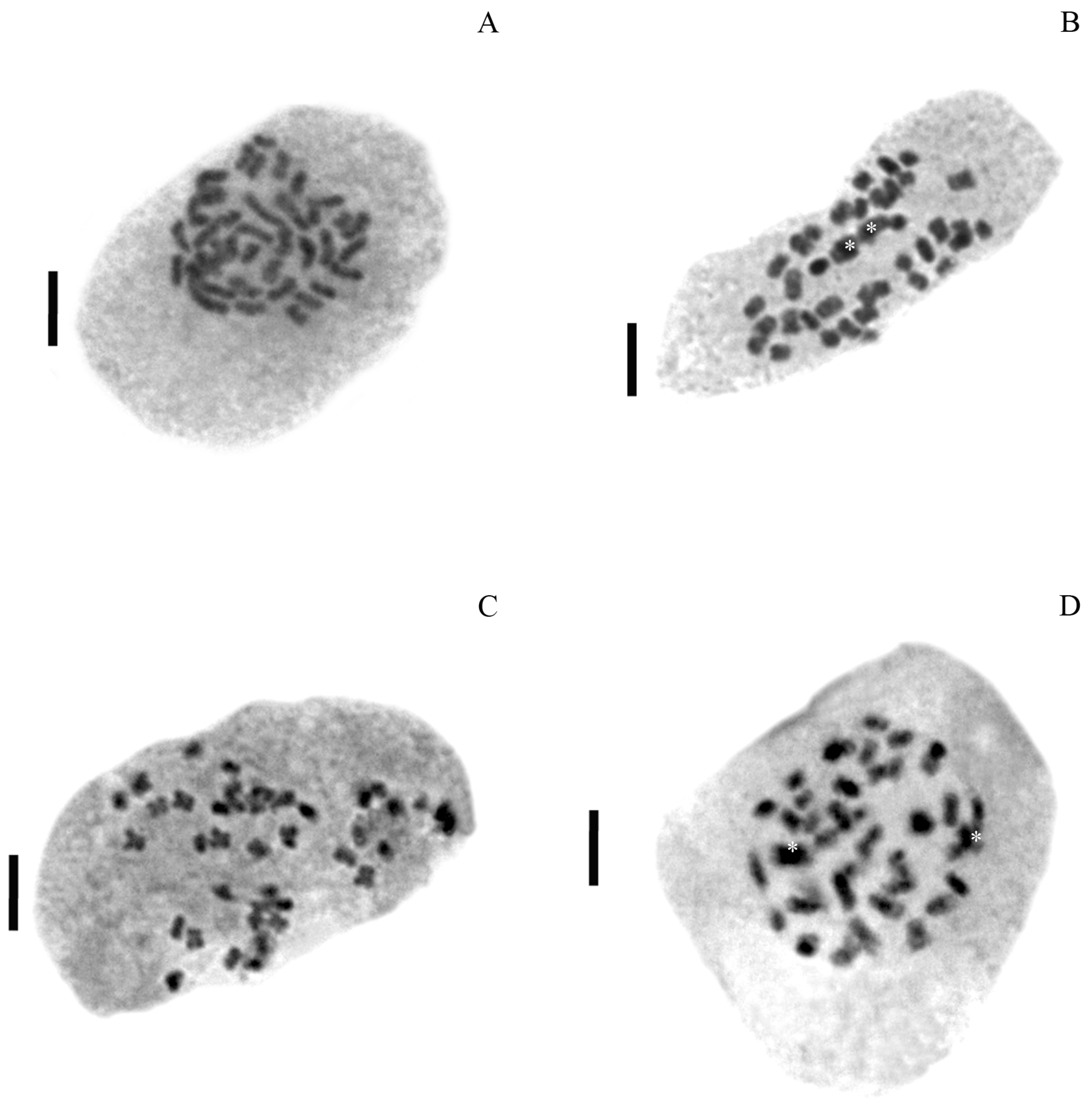
3.1. Ploidy Estimations
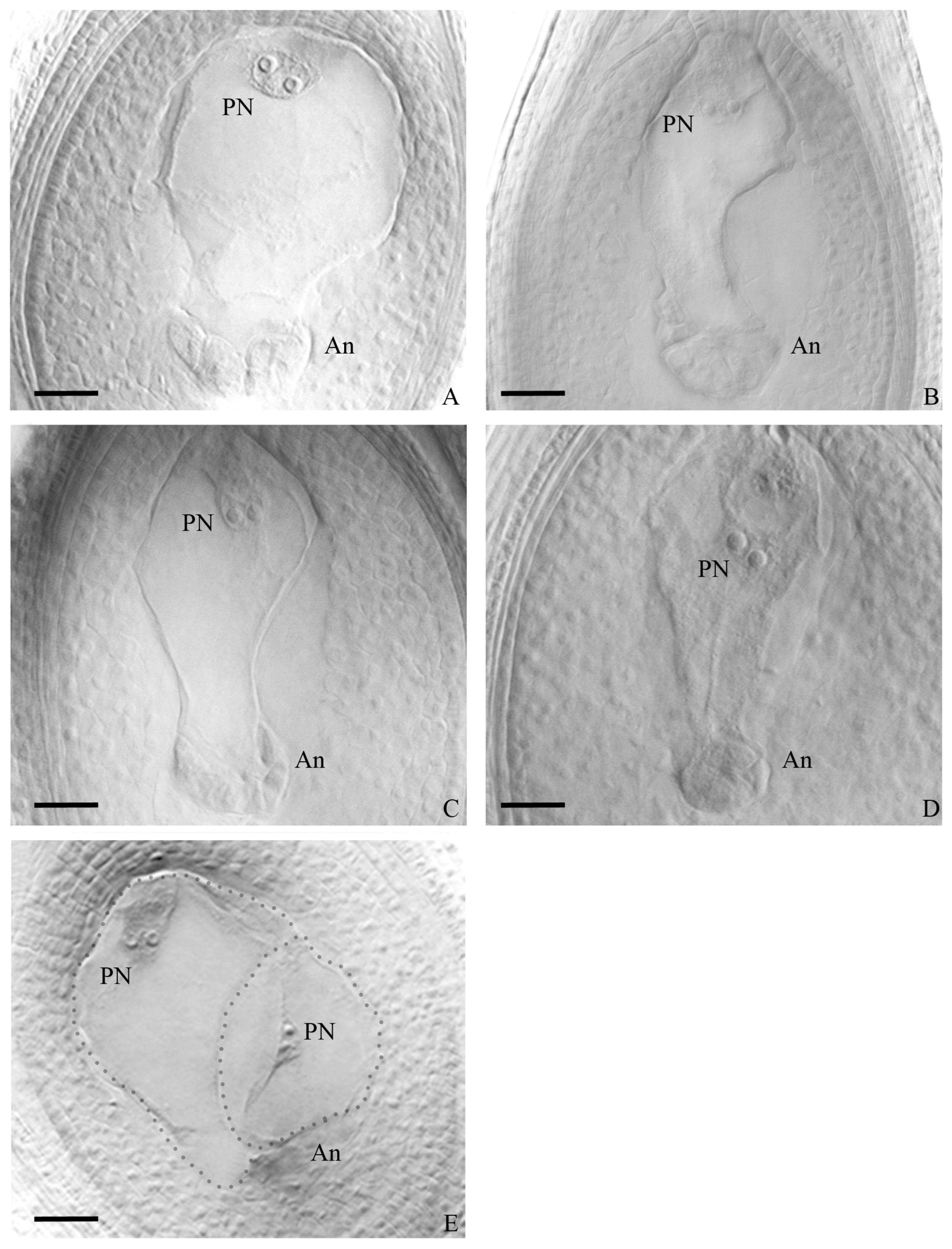
3.2. Embryo Sac Analysis
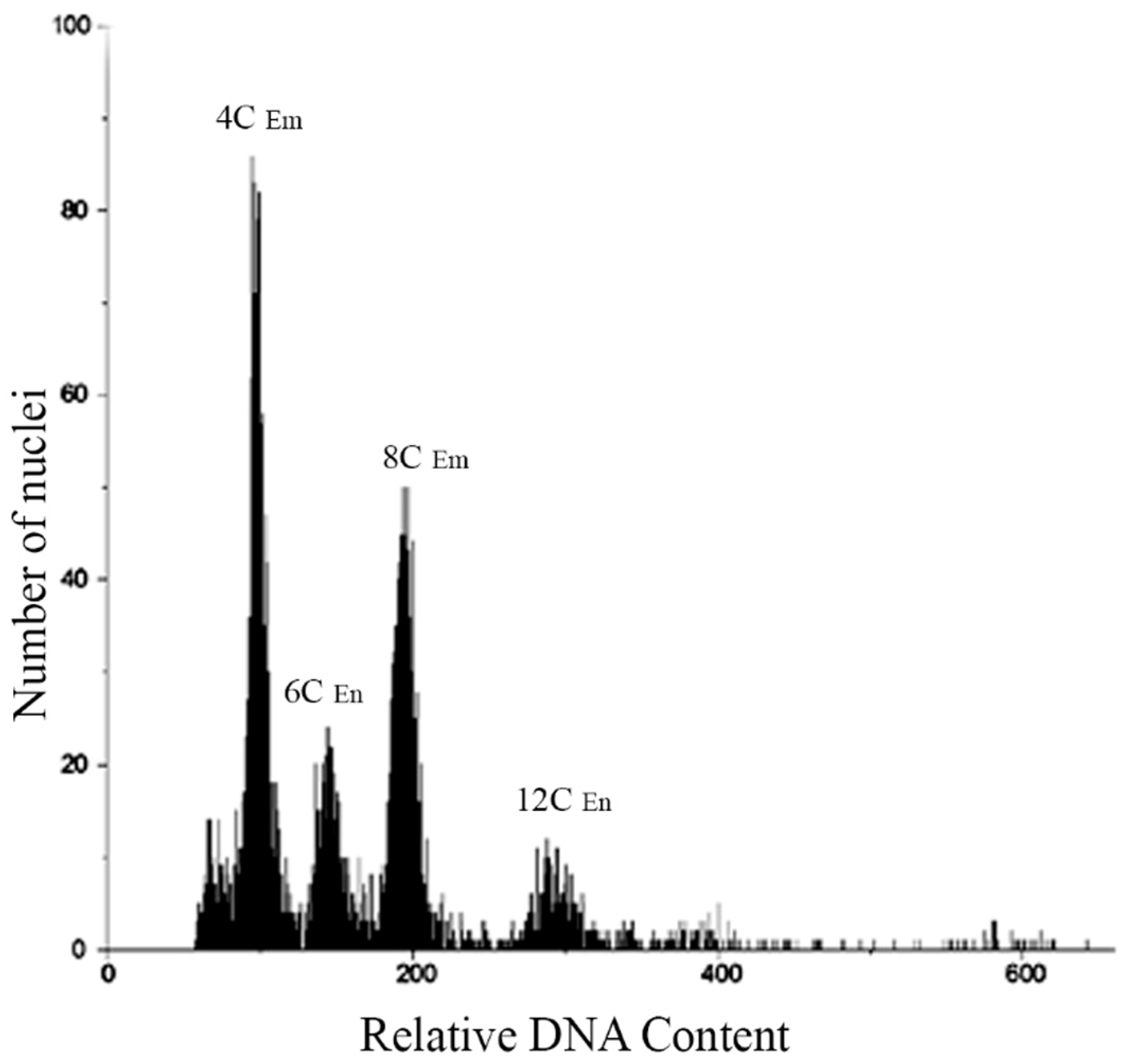
3.3. Flow Cytometric Seed Screen
3.4. Reproductive Variability in Ovules and Seeds
3.5. Mating System and Seed Fertility
4. Discussion
4.1. Single Ploidies Featured in All Species
4.2. Reproductive Pathways and Cytotype Stability
4.3. Two Mating Systems with Partial Breakdown of Self-Incompatibility
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meirmans, P.G.; Van Tienderen, P.H. The effects of inheritance in tetraploids on genetic diversity and population divergence. Heredity 2013, 110, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Wickett, N.J.; Ayampalayam, S.; Chanderbali, A.S.; Landherr, L.; Ralph, P.E.; Tomsho, L.P.; Hu, Y.; Liang, H.; Soltis, P.S.; et al. Ancestral polyploidy in seed plants and angiosperms. Nature 2011, 473, 97–100. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.L. Variation and Evolution in Plants; Columbia University Press: New York, NY, USA, 1950. [Google Scholar]
- Grant, V. Polyploidy; Columbia University Press: New York, NY, USA, 1981. [Google Scholar]
- Otto, S.P.; Whitton, J. Polyploid incidence and evolution. Ann. Rev. Genet. 2000, 34, 401–437. [Google Scholar] [CrossRef] [PubMed]
- Levin, D.A. The Role of Chromosomal Change in Plant Evolution; Oxford University Press: New York, NY, USA, 2002. [Google Scholar]
- Levin, D.A. Polyploidy and novelty in flowering plants. Amer. Nat. 1983, 122, 1–25. [Google Scholar] [CrossRef]
- Lewis, W.H. Polyploidy: Biological Relevance; Plenum Press: New York, NY, USA, 1980. [Google Scholar]
- Osborne, O.; Simon, S.; Collins, S. Attitudes towards science: A review of the literature and its implications. Int. J. Sci. Educ. 2003, 25, 1049–1079. [Google Scholar] [CrossRef]
- Coyne, J.A.; Orr, H.A. Speciation; Sinauer Associates: Sunderland, MA, USA, 2004. [Google Scholar]
- Suda, J.; Kron, P.; Husband, B.C.; Travnıcek, P. Flow cytometry and ploidy: Applications in plant systematics, ecology and evolutionary biology. In Flow Cytometry with Plant Cells: Analysis of Genes, Chromosomes and Genomes; Dolezel, J., Greilhuber, J., Suda, J., Eds.; Wiley–VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2007; pp. 103–130. [Google Scholar]
- Lumaret, R. Polyploidy and the critical size of natural populations: The case of cocksfoot (Dactylis glomerata L.), a grass used as a fodder plant. Bocconea 1997, 7, 133–139. [Google Scholar]
- Karunarathne, P.; Schedler, M.; Martínez, E.J.; Honfi, A.I.; Novichkova, A.; Hojsgaard, D.H. Intraspecific ecological niche divergence and reproductive shifts foster cytotype displacement and provide ecological opportunity to polyploids. Ann. Bot. 2018, 121, 1183–1196. [Google Scholar] [CrossRef]
- Karunarathne, P.; Reutemann, A.V.; Schedler, M.; Glücksberg, A.; Martínez, E.J.; Honfi, A.I.; Hojsgaard, D.H. Sexual modulation in a polyploidy grass: A reproductive contest between environmentally inducible sexual and genetically dominant apomictic pathways. Sci. Rep. 2020, 10, 8319. [Google Scholar] [CrossRef]
- Rua, G.H.; Speranza, P.R.; Vaio, M.; Arakaki, M. A phylogenetic analysis of the genus Paspalum (Poaceae) based on cpDNA and morphology. Plant Syst. Evol. 2010, 288, 227–243. [Google Scholar] [CrossRef]
- Bonasora, M.G.; Pozzobon, M.T.; Honfi, A.I.; Rua, G.H. Paspalum schesslii (Poaceae, Paspaleae), a new species from Mato Grosso (Brazil) with an unusual base chromosome number. Plant Syst. Evol. 2015, 301, 2325–2339. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Morrone, O. Revisión de las especies de Paspalum para América del Sur austral (Argentina, Bolivia, sur del Brasil, Chile, Paraguay y Uruguay). Ann. Mo. Bot. Gard. Monogr. Syst. Bot. 2005, 102, 1–297. [Google Scholar]
- Giussani, L.M.; Zuloaga, F.O.; Quarin, C.L.; Cota-Sánchez, H.; Ubayasena, K.; Morrone, O. Phylogenetic relationships in the genus Paspalum (Poaceae: Panicoideae: Paniceae): An assessment of the Quadrifaria and Virgata informal groups. Syst. Bot. 2009, 34, 32–43. [Google Scholar] [CrossRef]
- Quarin, C.L. Relaciones citotaxonómicas entre Paspalurn almum Chase y P. hexastachyum Parodi (Gramineae). Bonplandia 1974, 3, 115–127. [Google Scholar] [CrossRef]
- Davidse, G.; Pohl, R.W. Chromosome numbers meiotic behaviour and notes on tropical American grasses (Gramineae). Can. J. Bot. 1974, 52, 317–328. [Google Scholar] [CrossRef]
- Ortiz, J.P.A.; Quarin, C.L.; Pessino, S.C.; Acuña, C.A.; Martínez, E.J.; Espinoza, F.; Hojsgaard, D.H.; Sartor, M.E.; Cáceres, M.E.; Pupilli, F. Harnessing apomictic reproduction in grasses: What we have learned from Paspalum. Ann. Bot. 2013, 112, 767–787. [Google Scholar] [CrossRef] [PubMed]
- Darlington, C.D. Evolution of Genetic Systems; Cambridge University Press: Cambridge, UK, 1939. [Google Scholar]
- Hojsgaard, D.H.; Martínez, E.J.; Quarin, C.L. Competition between meiotic and apomictic pathways during ovule and seed development results in clonality. N. Phytol. 2013, 197, 336–347. [Google Scholar] [CrossRef]
- Norrmann, G.A. Citología y modo de reproducción en dos especies de Paspalum (Gramineae). Bonplandia 1981, 17, 149–158. [Google Scholar]
- Quarin, C.L.; Norrmann, G.A.; Espinoza, F. Evidence for autoploidy in apomictic Paspalum rufum. Hereditas 1998, 129, 119–124. [Google Scholar] [CrossRef]
- Hörandl, E.; Paun, O. Patterns and sources of genetic diversity in apomictic plants: Implications for evolutionary potentials. In Apomixis: Evolutions, Mechanisms and Perspectives; Hörandl, E., Grossniklaus, U., van Dijk, P.J., Sharbel, T.F., Eds.; A.R.G. Gantner Verlag: Rugell, Liechtenstein, 2007; pp. 170–194. [Google Scholar]
- Brugnoli, E.A.; Urbani, M.H.; Quarin, C.L.; Zilli, A.L.; Martínez, E.J.; Acuña, C.A. Diversity in apomictic populations of Paspalum simplex Morong. Crop Sci. 2014, 54, 1656–1664. [Google Scholar] [CrossRef]
- Daurelio, L.D.; Espinoza, F.; Quarin, C.L.; Pessino, S.C. Genetic diversity in sexual diploid and apomictic tetraploid populations of Paspalum notatum situated in sympatry or allopatry. Plant Syst. Evol. 2004, 244, 189–199. [Google Scholar] [CrossRef]
- Sartor, M.E.; Quarin, C.L.; Urbani, M.H.; Espinoza, F. Ploidy levels and reproductive behaviour in natural populations of five Paspalum species. Plant Syst. Evol. 2011, 293, 31–41. [Google Scholar] [CrossRef]
- Urbani, M.H.; Quarin, C.L.; Espinoza, F.; Penteado, M.I.O.; Rodrigues, I.F. Cytogeography and reproduction of the Paspalum simplex polyploid complex. Plant Syst. Evol. 2002, 236, 99–105. [Google Scholar] [CrossRef]
- Gornall, R.J. Population genetic structure in agamospermous plants. In Molecular Systematics and Plant Evolution; Hollingsworth, P.M., Bateman, R.M., Gornall, R.J., Eds.; Taylor & Francis: London, UK, 1999; pp. 118–138. [Google Scholar]
- Quarin, C.L. The nature of apomixis and its origin in Panicoid grasses. Apomixis Newsl. 1992, 5, 8–15. [Google Scholar]
- Burson, B.L. Cytology of Paspalum chacoense and P. durifolium and their relationship to P. dilatatum. Bot. Gaz. 1985, 146, 124–129. [Google Scholar] [CrossRef]
- Burson, B.L.; Bennett, H.W. Cytology and reproduction of three Paspalum species. J. Hered. 1970, 61, 129–132. [Google Scholar] [CrossRef]
- Honfi, A.I.; Quarin, C.L. Citología y modo de reproducción de Paspalum durifolium Mez pentaploide (Poaceae: Panicoideae: Paniceae). Lilloa 2009, 45, 85. [Google Scholar]
- Moraes Fernandes, M.I.B.; Barreto, I.; Salzano, F.M.; Sacchet, A.M.O.F. Cytological and evolutionary relationships in Brazilian forms of Paspalum (Gramineae). Caryologia 1974, 27, 455–465. [Google Scholar]
- Quarin, C.L.; Norrmann, G.A. Cytology and reproductive behavior of Paspalum equitans, P. ionanthum, and their hybrids with diploid and tetraploid cytotypes of P. cromyorrhizon. Bot. Gaz. 1987, 148, 386–391. [Google Scholar] [CrossRef]
- Pozzobon, M.T.; Valls, J.F.M.; Dos Santos, S. Contagens cromossômicas em espécies brasileiras de Paspalum L. (Gramineae). Acta Bot. Bras. 2000, 14, 151–162. [Google Scholar] [CrossRef]
- Quarin, C.L. A tetraploid cytotype of Paspalum durifolium: Cytology, reproductive behavior and its relationship to diploid P. intermedium. Hereditas 1994, 121, 115–118. [Google Scholar] [CrossRef]
- Martínez, E.J.; Quarin, C.L.; Hayward, M.D. Genetic control of apospory in apomictic Paspalum species. Cytologia 1999, 64, 425–433. [Google Scholar] [CrossRef]
- Pozzobon, M.T.; Carvalho Machado, A.C.; Vaio, M.; Valls, J.F.M.; de Souza Peñaloza, A.D.; dos Santos, S.; Côrtes, A.L.; Rua, G.H. Cytogenetic analyses in Paspalum L. reveal new diploid species and accessions. Ciência Rural. 2008, 38, 1292–1298. [Google Scholar] [CrossRef]
- Bashaw, E.C.; Hovin, A.W.; Holt, E.C. Apomixis, its evolutionary significance and utilization in plant breeding. In Proceedings of the XI International Grassland Congress, Surfers Paradise, QLD, Australia, 13–23 April 1970; Norman, M.J.T., Ed.; University of Queensland Press: St. Lucia, Australia, 1970; pp. 245–248. [Google Scholar]
- Honfi, A.I.; Quarin, C.L.; Valls, J.F.M. Estudios cariológicos en gramíneas sudamericanas. Darwiniana 1990, 30, 87–94. [Google Scholar]
- Pagliarini, M.S.; Carraro, L.R.; Freitas, P.M.; Adamowsky, E.V.; Rocha Batista, L.A.; Valls, J.F.M. Cytogenetic characterization of Brazilian Paspalum accessions. Hereditas 2002, 135, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Hojsgaard, D.H.; Honfi, A.I.; Rua, G.H.; Daviña, J.R. Chromosome numbers and ploidy levels of Paspalum species from subtropical South America (Poaceae). Genet. Resour. Crop Evol. 2009, 56, 533–545. [Google Scholar] [CrossRef]
- Pozzobon, M.T.; Valls, J.F.M. Chromosome number in germplasm accessions of Paspalum notatum (Gramineae). Braz. J. Genet. 1997, 20, 29–34. [Google Scholar] [CrossRef]
- Brugnoli, A.E.; Urbani, M.H.; Quarin, C.L.; Martínez, E.J.; Acuña, C.A. Diversity in diploid, tetraploid and mixed diploid-tetraploid populations of Paspalum simplex. Crop Sci. 2013, 53, 1509–1516. [Google Scholar] [CrossRef]
- Hijmans, R.J.; Guarino, L.; Cruz, M.; Rojas, E. Computer tools for spatial analysis of plant genetic resources data: 1. DIVA-GIS. Plant Gen. Resour. News. 2001, 127, 15–29. [Google Scholar]
- Galdeano, F.; Urbani, M.H.; Sartor, M.E.; Honfi, A.I.; Espinoza, F.; Quarin, C.L. Relative DNA content in diploid, polyploid, and multiploid species of Paspalum (Poaceae) with relation to reproductive mode and taxonomy. J. Plant Res. 2016, 129, 697–710. [Google Scholar] [CrossRef]
- Young, B.A.; Sherwood, R.T.; Bashaw, E.C. Cleared-pistil and thick-sectioning techniques for detecting aposporous apomixis in grasses. Canad. J. Bot. 1979, 57, 1668–1672. [Google Scholar] [CrossRef]
- Zilli, A.L.; Brugnoli, E.A.; Marcón, F.; Billa, M.B.; Rios, E.F.; Martínez, E.J.; Acuña., C.A. Heterosis and expressivity of apospory in tetraploid Bahiagrass hybrids. Crop Sci. 2015, 55, 1189–1201. [Google Scholar] [CrossRef]
- Matzk, F.; Meister, A.; Schubert, I. An efficient screen for reproductive pathways using mature seeds of monocots and dicots. Plant J. 2000, 21, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Siena, L.A.; Sartor, M.E.; Espinoza, F.; Quarin, C.L.; Ortiz, J.P.A. Genetic and embryological evidences of apomixis at the diploid level in Paspalum rufum support recurrent auto-polyploidization in the species. Sex. Plant Reprod. 2008, 21, 205–215. [Google Scholar] [CrossRef]
- Reutemann, A.V.; Honfi, A.I.; Karunarathne, P.; Eckers, F.; Hojsgaard, D.H.; Martínez, E.J. Variation of residual sexuality rates along reproductive development in apomictic tetraploids of Paspalum. Plants 2022, 11, 1639. [Google Scholar] [CrossRef]
- Balzarini, M.; Di Rienzo, J. Info-Gen: Software Para anÁlisis Estadístico de Datos Genéticos; Facultad de Ciencias Agropecuarias, Universidad Nacional de Córdoba: Córdoba, Argentina, 2003. [Google Scholar]
- Saura, F. Cariología de gramíneas. Rev. De La Fac. De Agron. íA Y Vet. Univ. De Buenos Aires 1943, 2, 63–74. [Google Scholar]
- Pagliarini, M.S.; De Freitas, P.M.; Takayama, S.Y.; Batista, L.A.R. An original meiotic mutation in Paspalum regnellii. Sex. Plant Reprod. 1998, 11, 17–21. [Google Scholar] [CrossRef]
- Burton, G.W. A cytological study of some species in the genus Paspalum. J. Agric. Res. 1940, 60, 193–198. [Google Scholar]
- Burton, G.W. A cytological study of some species in the tribe Paniceae. Am. J. Bot. 1942, 19, 355–359. [Google Scholar] [CrossRef]
- Brown, W.V. A cytological study of some Texas gramineae. Bull. Torrey Bot. Club 1950, 77, 63–76. [Google Scholar] [CrossRef]
- Brown, W.V.; Emery, H.P. Apomixis in the Gramineae: Panicoideae. Am. J. Bot. 1958, 45, 253–263. [Google Scholar] [CrossRef]
- Nielsen, E.L. Grass studies III. Additional somatic chromosome complements. Am. J. Bot. 1939, 26, 366–372. [Google Scholar] [CrossRef]
- Reutemann, A.V.; Martínez, E.J.; Schedler, M.; Daviña, J.R.; Hojsgaard, D.H.; Honfi, A.I. Uniparentality: Advantages for range expansion in diploid and diploid-autopolyploid species. Bot. J. Linn. Soc. 2022, 200, 563–585. [Google Scholar] [CrossRef]
- Delgado, L.; Galdeano, F.; Sartor, M.E.; Quarin, C.L.; Espinoza, F.; Ortiz, J.P. Analysis of variation for apomictic reproduction in diploid Paspalum rufum. Ann. Bot. 2014, 113, 1211–1218. [Google Scholar] [CrossRef] [PubMed]
- Quarin, C.L. Seasonal changes in the incidences of apomixis of diploid, triploid, and tetraploid plants of Paspalum cromyorrhizon. Euphytica 1986, 35, 515–522. [Google Scholar] [CrossRef]
- Norrmann, G.A.; Quarin, C.L.; Burson, B.L. Cytogenetics and reproductive behavior of different chromosome races in six Paspalum species. J. Hered. 1989, 80, 24–28. [Google Scholar] [CrossRef]
- Quarin, C.L.; Espinoza, F.; Martínez, E.J.; Pessino, S.C.; Bovo, O.A. A rise of ploidy level induces the expression of apomixis in Paspalum notatum. Sex. Plant Reprod. 2021, 13, 243–249. [Google Scholar] [CrossRef]
- Naumova, T.N.; Hayward, M.D.; Wagenvoort, M. Apomixis and sexuality in diploid and tetraploid accessions of Brachiaria decumbens. Sex. Plant Rep. 1999, 12, 43–52. [Google Scholar] [CrossRef]
- Rebozzio, R.N.; Sartor, M.E.; Quarin, C.L.; Espinoza, F. Residual sexuality and its seasonal variation in natural apomictic Paspalum notatum accessions. Biol. Plant. 2011, 55, 391–395. [Google Scholar] [CrossRef]
- Urbani, M.H. Estudios sobre citología, sistema reproductivo y compatibilidad polen–pistilo de Panicum dichotomiflorum y Paspalum fasciculatum (Gramineae:Paniceae). Darwiniana 1996, 34, 193–198. [Google Scholar]
- Burton, G.W.; Forbes, I., Jr.; Jackson, J. Effect of ploidy on fertility and heterosis in Pensacola Bahiagrass. Crop Sci. 1970, 10, 63–66. [Google Scholar] [CrossRef]
- Quarin, C.L. Morfología, citología y sistema reproductivo de una nueva especie de gramínea: Paspalum procurrens. Bol. Soc. Arg. Bot. 1993, 29, 73–76. [Google Scholar]
- Quarin, C.L. Effect of pollen source and pollen ploidy on endosperm formation and seed set in pseudogamous apomictic Paspalum notatum. Sex. Plant Reprod. 1999, 11, 331–335. [Google Scholar] [CrossRef]
- Hörandl, E. The evolution of self-fertility in apomictic plants. Sex. Plant Rep. 2010, 23, 73–86. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, P.W.; Thorogood, D. Breakdown of self-incompatibility in perennial ryegrass at high temperature and its uses in breeding. Euphytica 1992, 64, 65–69. [Google Scholar] [CrossRef]
- Hirosaki, A.; Niikura, S. Developmental and environmental factors affecting level of self-incompatibility response in Brassica rapa L. Sex. Plant Rep. 2008, 21, 123–132. [Google Scholar] [CrossRef]
- Zilli, A.L.; Acuña, C.A.; Schulz, R.R.; Brugnoli, E.A.; Guidalevich, V.; Quarin, C.L.; Martínez, E.J. Widening the gene pool of sexual tetraploid Bahiagrass: Generation and reproductive characterization of a sexual synthetic tetraploid population. Crop Sci. 2018, 58, 762–772. [Google Scholar] [CrossRef]
- Yamamoto, M.; Nishimura, K.; Kitashiba, H.; Sakamoto, W.; Takeshi, N. High temperature causes breakdown of S haplotype-dependent stigmatic self-incompatibility in self-incompatible Arabidopsis thaliana. J. Exp. Bot. 2019, 70, 5745–5751. [Google Scholar] [CrossRef]
- Carter, A.L.; McNeilly, T. Effects of increased seed humidity on pollen tube growth and seed set following self-pollination in Brussels sprout (Brassica oleracea var. gemmifera). Euphytica 1975, 24, 805–813. [Google Scholar] [CrossRef]
- Ockendon, D.J. Effect of hexane and humidity on self-incompatibility in Brassica oleracea. Theor. Appl. Genet. 1978, 52, 113–117. [Google Scholar] [CrossRef]
- Barrett, S.C.H.; Harder, L.D. The ecology of mating and its evolutionary consequences in seed plants. Annu. Rev. Ecol. Evol. Syst. 2017, 48, 135–157. [Google Scholar] [CrossRef]
- Caponio, I.; Quarin, C.L. Intra- and interspecific hybridization between Dallisgrass and Vaseygrass. Crop Sci. 1990, 30, 362–364. [Google Scholar] [CrossRef]
- Mable, B.K. Polyploidy and self-compatibility: Is there an association? N. Phytol. 2004, 162, 803–811. [Google Scholar] [CrossRef] [PubMed]
- Barringer, B.C. Polyploidy and self-fertilization in flowering plants. Am. J. Bot. 2007, 94, 1527–1533. [Google Scholar] [CrossRef] [PubMed]
- Barrett, S.C.H.; Harder, L.D.; Worley, A.C. The comparative biology of pollination and mating in flowering plants. Phil. Trans. R. Soc. Lond. B Biol. Sci. 1996, 351, 1271–1280. [Google Scholar] [CrossRef]
- Bernardello, G.; Anderson, G.J.; Stuessy, T.F.; Crawford, D.J. A survey of floral traits, breeding systems, floral visitors, and pollination systems of the angiosperms of the Juan Fernández Islands (Chile). Bot. Rev. 2001, 67, 255–308. [Google Scholar] [CrossRef]
- Kao, T.H.; Tsukamoto, T. The molecular and genetic bases of S-RNase-based self-incompatibility. Plant Cell 2004, 16, S72–S83. [Google Scholar] [CrossRef]
- Charlesworth, D.; Vekemans, X.; Castric, V.; Glemin, S. Plant self-incompatibility systems: A molecular evolutionary perspective. N. Phytol. 2005, 168, 61–69. [Google Scholar] [CrossRef]
- Richards, A.J. Plant Breeding Systems, 2nd ed.; Chapman & Hall: London, UK, 1997. [Google Scholar]
- de Nettancourt, D. Incompatibility and Incongruity in Wild and Cultivated Plants, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2001. [Google Scholar] [CrossRef]
- Stone, J.L. Molecular mechanisms underlying the breakdown of gametophytic self-incompatibility. Quart. Rev. Biol. 2002, 77, 17–32. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Population Code (Voucher) | Collection Sites | Geographical Coordinates | No. Plants |
|---|---|---|---|---|
| P. durifolium | PD1 (M6) | Corrientes, Dpto. Ituzaingó, NR 12 | 27°37.622 S, 56°44.873 W | 19 |
| PD2 (M7) | Corrientes, Dpto. San Miguel, NR 118, and PR 5 | 27°55.901 S, 57°30.996 W | 18 | |
| PD3 (M16) | Corrientes, Dpto. Santo Tomé, Esteros del Iberá, PR 40 | 28°17.435 S, 56°46.219 W | 17 | |
| PD4 (M17) | Corrientes, Dpto. Santo Tomé, NR 120 | 27°49.513 S, 56°15.294 W | 18 | |
| PD5 (M18) | Corrientes, Dpto. Ituzaingó, NR 118 | 27°33.646 S, 57°09.184 W | 20 | |
| P. ionanthum | PI1 (M4) | Corrientes, Dpto. Ituzaingó, NR 120 | 27°49.265 S, 56°16.328 W | 20 |
| PI2 (M8) | Corrientes, Dpto. Concepción, Santa Rosa, NR 118 | 28°14.245 S, 58°03.252 W | 20 | |
| PI3 (M10) | Corrientes, Dpto. General Paz, Paso Florentín, PR 5 | 27°45.205 S, 57°45.658 W | 16 | |
| PI4 (M11) | Corrientes, Dpto. Mburucuyá, Parque Nacional Mburucuyá. | 28°02.024 S, 58°02.172 W | 20 | |
| PI5 (M12) | Corrientes, Dpto. Goya, Paraje Marucha, NR 12 | 29°09.906 S, 59°05.795 W | 20 | |
| P. regnellii | PR1 (M2) | Misiones, Dpto. Montecarlo, Puerto Laharrague, NR 12 | 26°32.492 S, 54°43.506 W | 20 |
| PR2 (M19) | Misiones, Dpto. San Ignacio, Jardín América, NR 12 | 27°03.170 S, 55°15.040 W | 19 | |
| PR3 (M20) | Misiones, Dpto. San Pedro, NR 14 | 26°43.626 S, 54°14.845 W | 20 | |
| PR4 (M21) | Misiones, Dpto. 25 de Mayo, PR 2 | 27°22.441 S, 54°25.503 W | 18 | |
| PR5 (M22) | Misiones, Dpto. 25 de Mayo, PR 4 | 27°46.628 S, 55°14.998 W | 19 | |
| P. urvillei | PU1 (M1) | Misiones, Dpto. San Ignacio, 2,2 Km East PR 210 | 27°16.613 S, 55°27.735 W | 17 |
| PU2 (M5) | Corrientes, Dpto. Ituzaingó, NR 120 | 27°49.265 S, 56°16.328 W | 20 | |
| PU3 (M14) | Entre Ríos, Dpto. La Paz, PR 6 | 31°01.223 S, 59°25.222 W | 20 | |
| PU4 (M23) | Santa Fe, Dpto. Gral. Obligado, NR 11 | 28°35.733 S, 59°25.005 W | 20 | |
| PU5 (M27) | Chaco, Dpto. Gral. San Martin, PR 90 | 26°24.117 S, 59°22.726 W | 17 |
| Species | Pop | n | Number of Ovules Bearing (%) | Proportions | |||||
|---|---|---|---|---|---|---|---|---|---|
| MES | AES | MES + AES | AbES | SP | AP | p | |||
| P. durifolium | PD1 | 152 | 127 (83.5) | - | 1 (0.7) | 24 (15.8) | 0.99 | 0.01 | <0.001 |
| PD2 | 150 | 141 (94.0) | - | - | 9 (6.0) | 1.00 | 0.00 | <0.001 | |
| PD3 | 152 | 134 (88.2) | - | 6 (3.9) | 12 (7.9) | 0.96 | 0.04 | <0.001 | |
| PD4 | 153 | 129 (84.3) | - | 10 (6.5) | 14 (9.2) | 0.93 | 0.07 | <0.001 | |
| PD5 | 156 | 124 (79.5) | - | 5 (3.2) | 27 (17.3) | 0.96 | 0.04 | <0.001 | |
| P. ionanthum | PI1 | 158 | 150 (95.0) | - | 4 (2.5) | 4 (2.5) | 0.97 | 0.03 | <0.001 |
| PI2 | 151 | 147 (97.4) | - | 2 (1.3) | 2 (1.3) | 0.99 | 0.01 | <0.001 | |
| PI3 | 150 | 149 (99.3) | - | - | 1 (0.7) | 1.00 | 0.00 | <0.001 | |
| PI4 | 165 | 160 (97.0) | - | - | 5 (3.0) | 1.00 | 0.00 | <0.001 | |
| PI5 | 158 | 156 (98.7) | - | - | 2 (1.3) | 1.00 | 0.00 | <0.001 | |
| P. regnellii | PR1 | 160 | 112 (70.0) | - | - | 48 (30.0) | 1.00 | 0.00 | <0.001 |
| PR2 | 151 | 132 (87.4) | - | - | 19 (12.6) | 1.00 | 0.00 | <0.001 | |
| PR3 | 155 | 148 (95.5) | - | - | 7 (4.5) | 1.00 | 0.00 | <0.001 | |
| PR4 | 170 | 162 (95.3) | - | - | 8 (4.7) | 1.00 | 0.00 | <0.001 | |
| PR5 | 164 | 153 (93.3) | - | - | 11 (6.7) | 1.00 | 0.00 | <0.001 | |
| P. urvillei | PU1 | 167 | 160 (95.8) | - | - | 7 (4.2) | 1.00 | 0.00 | <0.001 |
| PU2 | 158 | 153 (96.8) | - | - | 5 (3.2) | 1.00 | 0.00 | <0.001 | |
| PU3 | 156 | 155 (99.4) | - | - | 1 (0.6) | 1.00 | 0.00 | <0.001 | |
| PU4 | 156 | 151 (96.8) | - | - | 5 (3.2) | 1.00 | 0.00 | <0.001 | |
| PU5 | 155 | 155 (100) | - | - | - | 1.00 | 0.00 | <0.001 | |
| Species | Pop | Sexual Pathways | Apomictic Pathways | Statistical Analysis | Reproductive Efficiency | ||||
|---|---|---|---|---|---|---|---|---|---|
| Ei | Oi | Ei | Oi | χ2 | p | Sex | Apo | ||
| P. durifolium | PD1 | 0.99 | 1.00 | 0.01 | 0.00 | 4.67 × 10−31 | 1.00 | 1.01 | - |
| PD2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.06 | - | |
| PD3 | 0.96 | 1.00 | 0.04 | 0.00 | 0.333 | 0.564 | 1.04 | - | |
| PD4 | 0.93 | 1.00 | 0.07 | 0.00 | 1.049 | 0.3055 | 1.07 | - | |
| PD5 | 0.96 | 1.00 | 0.04 | 0.00 | 0.237 | 0.626 | 1.04 | - | |
| P. ionanthum | PI1 | 0.97 | 1.00 | 0.03 | 0.00 | 0.036 | 0.849 | 1.03 | - |
| PI2 | 0.99 | 1.00 | 0.01 | 0.00 | 5.5 × 10−31 | 1.00 | 1.01 | - | |
| PI3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PI4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PI5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| P. regnellii | PR1 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - |
| PR2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PR5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| P. urvillei | PU1 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - |
| PU2 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU3 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU4 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| PU5 | 1.00 | 1.00 | 0.00 | 0.00 | - | - | 1.00 | - | |
| Species | Pop | Seed Set (%) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Self-Pollination | Open Pollination | Self- Pollination | Open Pollination | |||||||
| 1st Period | 2nd Period | p Value α | 1st Period | 2nd Period | p Value α | Total † | Total † | p Value β | ||
| P. durifolium | PD1 | 0.27 a | 0.61 a | 0.281 | 28.09 a | 29.58 a | 0.839 | 0.44 a | 28.83 b | <0.001 |
| PD2 | 0.35 a | 0.65 a | 0.333 | 46.30 a | 51.54 a | 0.326 | 0.50 a | 48.92 a | <0.001 | |
| PD3 | 0.16 a | 0.65 a | 0.262 | 57.90 a | 45.16 a | 0.242 | 0.41 a | 51.53 a | <0.001 | |
| PD4 | 1.24 a | 0.47 a | 0.409 | 45.52 a | 42.70 a | 0.881 | 0.86 a | 44.11 ab | <0.001 | |
| PD5 | 0.17 a | 1.18 a | 0.119 | 33.17 a | 27.97 a | 0.507 | 0.68 a | 30.57 b | <0.001 | |
| p value γ | 0.284 | 0.701 | 0.173 | 0.058 | 0.74 | 0.006 | ||||
| P. ionanthum | PI1 | 1.00 a | 1.00 a | 0.995 | 35.60 a | 23.20 a | 0.168 | 1.04 a | 29.41 c | <0.001 |
| PI2 | 0.26 a | 1.22 a | 0.418 | 60.25 a | 31.27 b | 0.002 | 0.75 a | 45.80 ab | <0.001 | |
| PI3 | 4.26 a | 1.36 a | 0.337 | 49.66 a | 58.88 a | 0.661 | 2.79 a | 54.27 a | <0.001 | |
| PI4 | 0.16 a | 1.10 a | 0.164 | 52.80 a | 39.22 a | 0.092 | 0.61 a | 46.02 ab | <0.001 | |
| PI5 | 2.85 a | 0.71 a | 0.089 | 45.53 a | 27.82 a | 0.036 | 1.80 a | 36.69 bc | <0.001 | |
| p value γ | 0.565 | 0.972 | 0.191 | 0.004 | 0.574 | 0.017 | ||||
| P. regnellii | PR1 | 8.51 a | 4.96 a | 0.440 | 23.47 a | 34.19 a | 0.360 | 6.73 b | 28.83 b | <0.001 |
| PR2 | 25.93 a | 6.19 b | 0.018 | 38.14 a | 44.17 a | 0.602 | 16.06 ab | 41.16 ab | 0.001 | |
| PR3 | 12.44 a | 9.74 a | 0.704 | 24.06 a | 28.70 a | 0.682 | 11.09 ab | 26.38 b | 0.031 | |
| PR4 | 10.48 a | 2.34 a | 0.160 | 35.83 a | 20.77 a | 0.144 | 6.41 b | 28.30 b | <0.001 | |
| PR5 | 25.16 a | 12.47 a | 0.055 | 62.20 a | 37.35 a | 0.103 | 18.81 a | 49.77 a | 0.001 | |
| p value γ | 0.059 | 0.142 | 0.006 | 0.426 | 0.039 | 0.026 | ||||
| P. urvillei | PU1 | 44.61 a | 40.75 a | 0.828 | 80.15 a | 59.76 a | 0.12 | 42.68 a | 69.96 a | 0.018 |
| PU2 | 30.76 a | 52.99 a | 0.071 | 86.87 a | 74.41 a | 0.269 | 41.88 a | 80.64 a | <0.001 | |
| PU3 | 38.47 a | 32.75 a | 0.671 | 92.01 a | 64.01 b | 0.007 | 35.61 a | 78.01 a | <0.001 | |
| PU4 | 33.34 a | 52.22 a | 0.224 | 76.71 a | 69.56 a | 0.428 | 42.78 a | 73.32 a | 0.004 | |
| PU5 | 31.80 a | 54.44 a | 0.057 | 74.00 a | 66.05 a | 0.391 | 43.12 a | 70.03 a | 0.002 | |
| p value γ | 0.906 | 0.200 | 0.091 | 0.767 | 0.893 | 0.483 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schedler, M.; Reutemann, A.V.; Hojsgaard, D.H.; Zilli, A.L.; Brugnoli, E.A.; Galdeano, F.; Acuña, C.A.; Honfi, A.I.; Martínez, E.J. Alternative Evolutionary Pathways in Paspalum Involving Allotetraploidy, Sexuality, and Varied Mating Systems. Genes 2023, 14, 1137. https://doi.org/10.3390/genes14061137
Schedler M, Reutemann AV, Hojsgaard DH, Zilli AL, Brugnoli EA, Galdeano F, Acuña CA, Honfi AI, Martínez EJ. Alternative Evolutionary Pathways in Paspalum Involving Allotetraploidy, Sexuality, and Varied Mating Systems. Genes. 2023; 14(6):1137. https://doi.org/10.3390/genes14061137
Chicago/Turabian StyleSchedler, Mara, Anna Verena Reutemann, Diego Hernán Hojsgaard, Alex Leonel Zilli, Elsa Andrea Brugnoli, Florencia Galdeano, Carlos Alberto Acuña, Ana Isabel Honfi, and Eric Javier Martínez. 2023. "Alternative Evolutionary Pathways in Paspalum Involving Allotetraploidy, Sexuality, and Varied Mating Systems" Genes 14, no. 6: 1137. https://doi.org/10.3390/genes14061137
APA StyleSchedler, M., Reutemann, A. V., Hojsgaard, D. H., Zilli, A. L., Brugnoli, E. A., Galdeano, F., Acuña, C. A., Honfi, A. I., & Martínez, E. J. (2023). Alternative Evolutionary Pathways in Paspalum Involving Allotetraploidy, Sexuality, and Varied Mating Systems. Genes, 14(6), 1137. https://doi.org/10.3390/genes14061137