
Screening of Suitable Reference Genes for Immune Gene Expression Analysis Stimulated by Vibrio anguillarum and Copper Ions in Chinese Mitten Crab (Eriocheir sinensis)

 ,
,
Abstract
1. Introduction
2. Material and Methods
2.1. Animals
2.2. Vibrio anguillarum and Copper Ion Stimulation
2.3. RNA Extraction and Reverse Transcription (RT)
2.4. Primer Specificity and Amplification Efficiency
2.5. Relative Quantification of Gene Expression
2.6. Data Analysis
3. Results and Analysis
3.1. Specificity and Amplification Efficiency of Primers
3.2. Ct Values of 10 Candidate Reference Genes
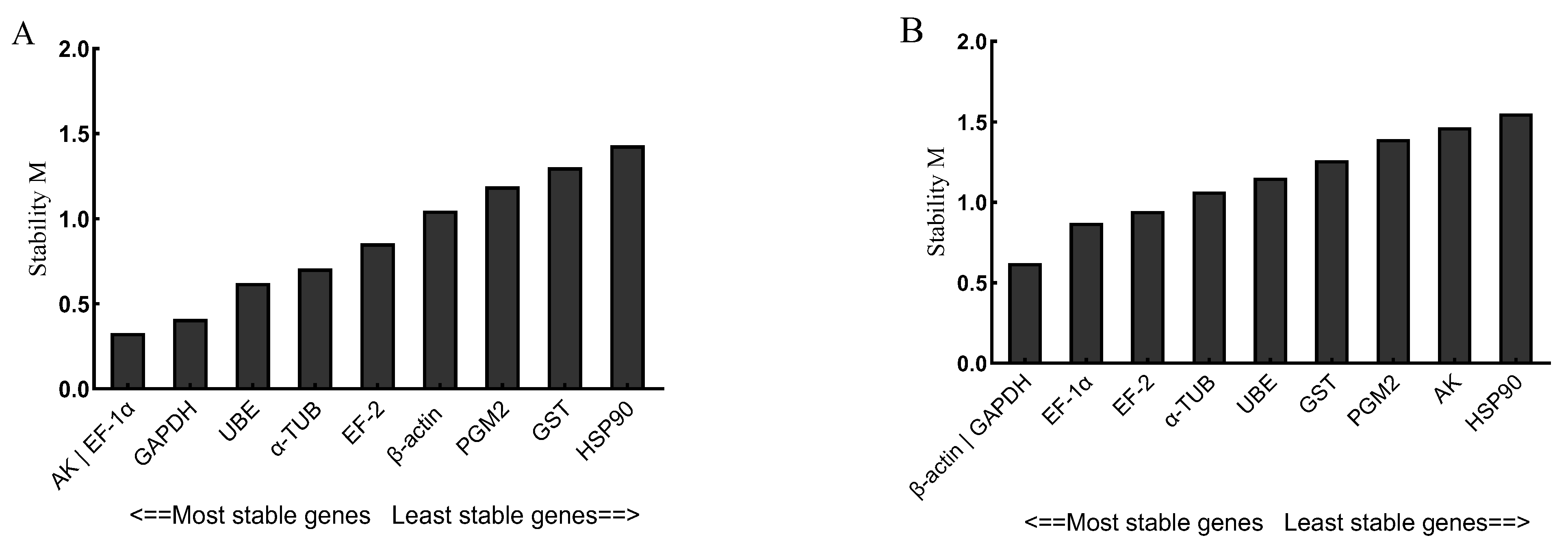
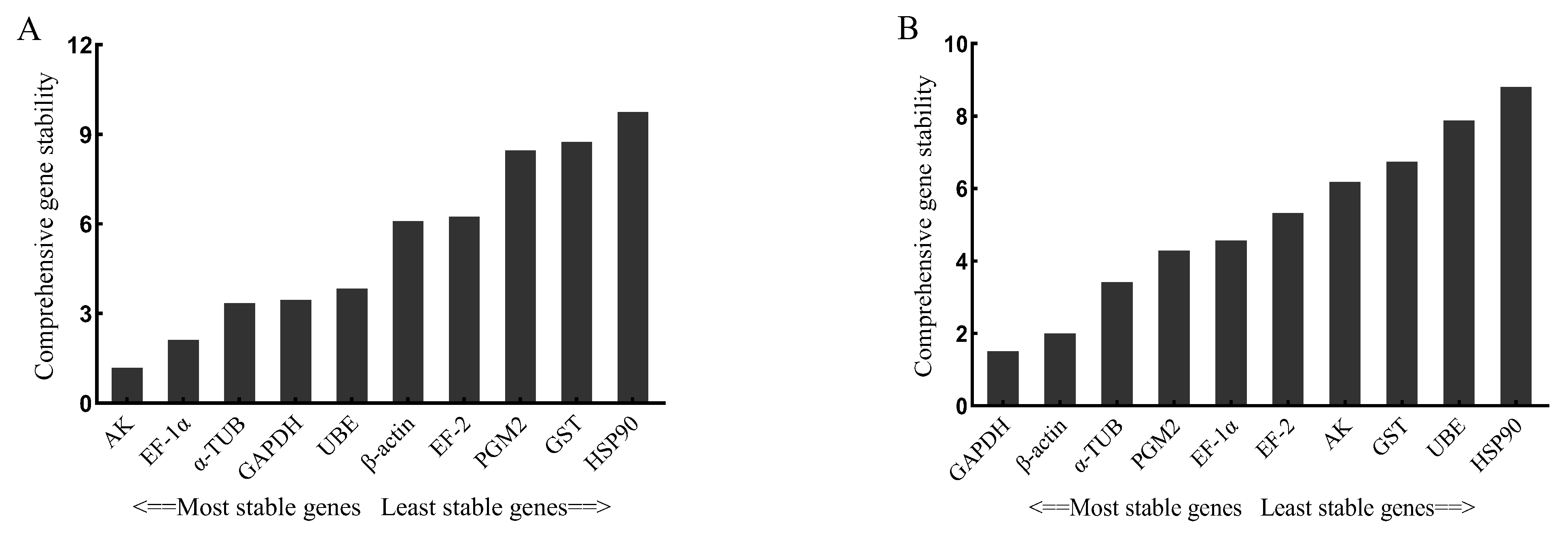
3.3. Stability of 10 Candidate Reference Genes
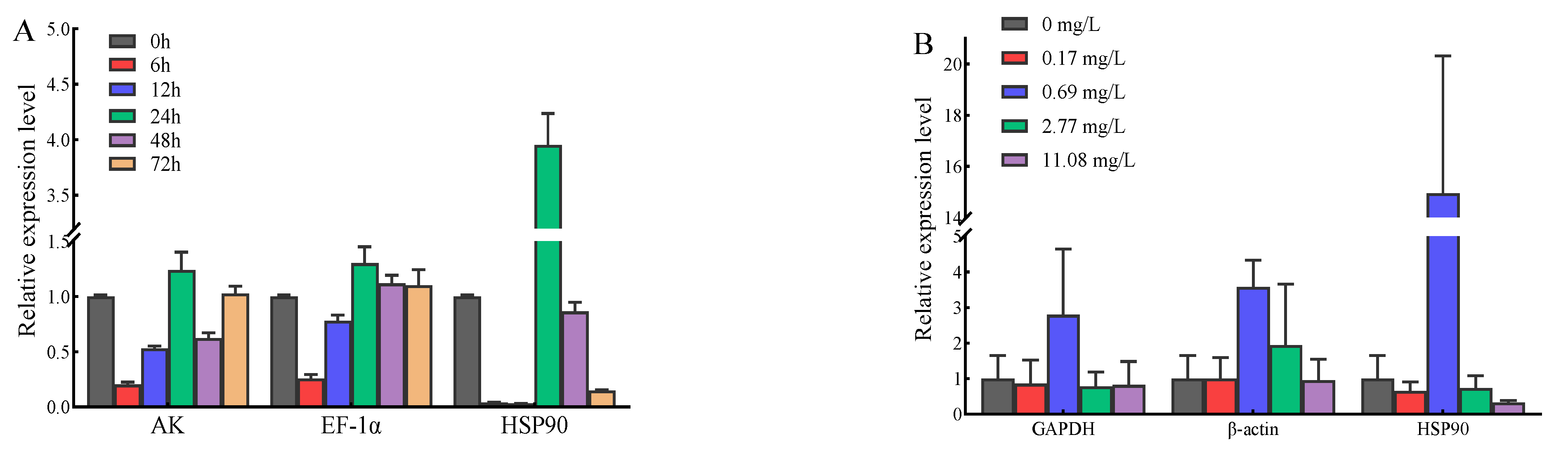
3.4. Validation of Reference Genes
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Valasek, M.A.; Repa, J.J. The power of real-time PCR. Adv. Physiol. Educ. 2005, 29, 151–159. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Bustin, S.A.; Benes, V.; Garson, J.A.; Hellemans, J.; Huggett, J.; Kubista, M.; Mueller, R.; Nolan, T.; Pfaffl, M.W.; Shipley, G.L.; et al. The MIQE Guidelines: Minimum information for publication of quantitative real-time PCR experiments. Clin. Chem. 2009, 55, 611–622. [Google Scholar] [CrossRef] [PubMed]
- Su, J.G.; Zhang, R.F.; Dong, J.; Yang, C.R. Evaluation of internal control genes for qRT-PCR normalization in tissues and cell culture for antiviral studies of grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 2011, 30, 830–835. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Zakrajsek, B.A. Effect of experimental treatment on housekeeping gene expression: Validation by real-time, quantitative RT-PCR. J. Biochem. Bioph. Meth. 2000, 46, 69–81. [Google Scholar] [CrossRef]
- Tang, Y.K.; Yu, J.H.; Xu, P.; Li, J.L.; Li, H.X.; Ren, H.T. Identification of housekeeping genes suitable for gene expression analysis in Jian carp (Cyprinus carpio var. jian). Fish Shellfish Immunol. 2012, 33, 775–779. [Google Scholar] [CrossRef]
- Olsvik, P.A.; Lie, K.K.; Jordal, A.E.O.; Nilsen, T.O.; Hordvik, I. Evaluation of potential reference genes in real-time RT-PCR studies of Atlantic salmon. BMC Mol. Biol. 2005, 6, 21. [Google Scholar] [CrossRef]
- McCurley, A.T.; Callard, G.V. Characterization of housekeeping genes in zebrafish: Male-female differences and effects of tissue type, developmental stage and chemical treatment. BMC Mol. Biol. 2008, 9, 102. [Google Scholar] [CrossRef]
- Suzuki, T.; Higgins, P.J.; Crawford, D.R. Control selection for RNA quantitation. Biotechniques 2000, 29, 332–337. [Google Scholar] [CrossRef]
- Gong, J.W.; Li, Q.; Yu, H.; Liu, S.K.; Kong, L.F. Validation of housekeeping genes for gene expression analysis in iwagaki oyster (Crassostrea nippona) under salinity stress by quantitative real-time PCR. J. Ocean Univ. China 2020, 19, 1441–1446. [Google Scholar] [CrossRef]
- Huang, S.; Chen, X.W.; Wang, J.; Chen, J.; Yue, W.C.; Lu, W.Q.; Lu, G.Q.; Wang, C.H. Selection of appropriate reference genes for qPCR in the Chinese mitten crab, Eriocheir sinensis (Decapoda, Varunidae). Crustaceana 2017, 90, 275–296. [Google Scholar] [CrossRef]
- Leelatanawit, R.; Klanchui, A.; Uawisetwathana, U.; Karoonuthaisiri, N. Validation of reference genes for real-time PCR of reproductive system in the black tiger shrimp. PLoS ONE 2012, 7, e52677. [Google Scholar] [CrossRef] [PubMed]
- Benjamin, M.; Isabelle, A.; Nicole, F.; Renault, T. Identification of genes from flat oyster Ostrea edulis as suitable housekeeping genes for quantitative real time PCR. Fish Shellfish Immunol. 2010, 29, 937–945. [Google Scholar]
- Sellars, M.J.; Vuocolo, T.; Leeton, L.A.; Coman, G.J.; Degnan, B.M.; Preston, N.P. Real-time RT-PCR quantification of Kuruma shrimp transcripts: A comparison of relative and absolute quantification procedures. J. Biotechnol. 2007, 129, 391–399. [Google Scholar] [CrossRef]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, 00341–003411. [Google Scholar] [CrossRef]
- Andersen, C.L.; Jensen, J.L.; Orntoft, T.F. Normalization of real-time quantitative reverse transcription-PCR data: A model-based variance estimation approach to identify genes suited for normalization, applied to bladder and colon cancer data sets. Cancer Res. 2004, 64, 5245–5250. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Meller, M.; Vadachkoria, S.; Luthy, D.A.; Williams, M.A. Evaluation of housekeeping genes in placental comparative expression studies. Placenta 2005, 26, 601–607. [Google Scholar] [CrossRef]
- Dhar, A.K.; Bowers, R.M.; Licon, K.S.; Veazey, G.; Read, B. Validation of reference genes for quantitative measurement of immune gene expression in shrimp. J. Mol. Immunol. 2009, 46, 1688–1695. [Google Scholar] [CrossRef]
- Quispe, R.L.; Justino, E.B.; Vieira, F.N.; Jaramillo, M.L.; Rosa, R.D.; Perazzolo, L.M. Transcriptional profiling of immune-related genes in Pacific white shrimp (Litopenaeus vannamei) during ontogenesis. J. Fish Shellfish Immunol. 2016, 58, 103–107. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Zhang, S.; Lian, Y.; Ding, H.; Du, X.; Li, Z.; De Silva, S.S. Sustainable farming practices of the Chinese mitten crab (Eriocheir sinensis) around Hongze Lake, lower Yangtze River Basin, China. Ambio 2016, 45, 361–373. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Wu, X.; Li, J. Chinese mitten crab culture: Current status and recent progress towards sustainable development. Aquac. China Success Stories Mod. Trends 2018, 197–217. [Google Scholar] [CrossRef]
- Wang, N.; Wang, X.; Lin, Z.; Chen, X.; Bu, X.; Liu, S.; Lei, Y.; Shi, Q.; Qin, J.; Chen, L. Effects of dietary Zn on growth, antioxidant capacity, immunity and tolerance to lipopolysaccharide challenge in juvenile Chinese mitten crab Eriocheir sinensis. Aquac. Res. 2022, 53, 1110–1120. [Google Scholar] [CrossRef]
- Li, X.; Li, Z.; Liu, J.; De Silva, S.S. Advances in precocity research of the Chinese mitten crab Eriocheir sinensis. Aquac. Int. 2011, 19, 251–267. [Google Scholar] [CrossRef]
- Bonami, J.R.; Zhang, S. Viral diseases in commercially exploited crabs: A review. Invetebr. Pathol. 2011, 106, 6–17. [Google Scholar] [CrossRef]
- Rad, M.; Shahsavani, D. Isolation and characterization of Vibrio (Listonella) anguillarum from catfish. J. Turk. J. Vet. Anim. Sci. 2010, 34, 413–415. [Google Scholar] [CrossRef]
- Engel, D.W.; Brouwer, M. Crustaceans as models for metal metabolism: I. Effects of the molt cycle on blue crab. Metal Metabolism and Metallothionein. Mar. Environ. Res. 1993, 35, 1–5. [Google Scholar] [CrossRef]
- Rainbow, P.S. Physiology, physicochemistry and metal uptake-A crustacean perspective. Mar. Pollut. Bull. 1995, 31, 55–59. [Google Scholar] [CrossRef]
- Le Moullac, G.; Haffner, P. Environmental factors affecting immune responses in Crustacea. J. Aquac. 2000, 191, 121–131. [Google Scholar] [CrossRef]
- Lorenzon, S.; Francese, M.; Smith, V.J.; Ferrero, E.A. Heavy metals affect the circulating haemocyte number in the shrimp Palaemon elegans. J. Fish Shellfish Immunol. 2001, 11, 459–472. [Google Scholar] [CrossRef]
- Yeh, S.T.; Liu, C.H.; Chen, J.C. Effect of copper sulfate on the immune response and susceptibility to Vibrio alginolyticus in the white shrimp Litopenaeus vannamei. J. Fish Shellfish Immunol. 2004, 17, 437–446. [Google Scholar] [CrossRef]
- Choi, H.J.; Kang, S.W.; Yang, C.H.; Rhee, S.G.; Ryu, S.E. Crystal structure of a novel human peroxidase enzyme at 2.0 Å resolution. Nat. Struct. Biol. 1998, 5, 400–406. [Google Scholar] [CrossRef]
- Wong, C.M.; Siu, K.L.; Jin, D.Y. Peroxiredoxin-null yeast cells are hypersensitive to oxidative stress and are genomically unstable. J. Biol. Chem. 2004, 279, 23207–23213. [Google Scholar] [CrossRef]
- Wood, Z.A.; Schröder, E.; Harris, J.R.; Poole, L.B. Structure, mechanism and regulation of peroxiredoxins. Trends Biochem. Sci. 2003, 28, 32–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Huang, J.; Li, F.; Liu, S.; Liu, Q.; Wei, J.; Xiang, J. Molecular characterization, immune response against white spot syndrome virus infection of peroxiredoxin 4 in Fenneropenaeus chinensis and its antioxidant activity. Fish Shellfish. Immunol. 2014, 37, 38–45. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T) (-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hellemans, J.; Mortier, G.; De Paepe, A.; Speleman, F.; Vandesompele, J. qBase relative quantification framework and software for management and automated analysis of real-time quantitative PCR data. Genome Biol. 2007, 8, R19. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.M.; Tao, Z.; Shen, C.; Qian, D.; Wang, C.L.; Zhou, Q.C.; Jin, S. β-actin gene expression is variable among individuals and not suitable for normalizing mRNA levels in Portunus trituberculatus. Fish Shellfish Immunol. 2018, 81, 338–342. [Google Scholar] [CrossRef]
- Fink, T.; Lund, P.; Pilgaard, L.; Rasmussen, J.G.; Duroux, M.; Zachar, V. Instability of standard PCR reference genes in adipose-derived stem cells during propagation, differentiation and hypoxic exposure. BMC Mol. Biol. 2008, 9, 98. [Google Scholar] [CrossRef]
- Jaramillo, M.L.; Ammar, D.; Quispe, R.L.; Guzman, F.; Margis, R.; Nazari, E.M.; Müller, Y.M. Identification and evaluation of reference genes for expression studies by RT-qPCR during embryonic development of the emerging model organism, Macrobrachium olfersii. Gene 2017, 598, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Jin, Y.; He, L.; Lu, W.; Li, M. Suitable reference gene selection for different strains and developmental stages of the carmine spider mite, Tetranychus cinnabarinus, using quantitative real-time PCR. J. Insect Sci. 2010, 10, 208. [Google Scholar] [CrossRef] [PubMed]
- Huggett, J.; Dheda, K.; Bustin, S.; Zumla, A. Real-time RT-PCR normalization; strategies and considerations. Genes Immun. 2005, 6, 279–284. [Google Scholar] [CrossRef]
- Zhang, Y.M.; Lin, C.Y.; Li, B.Z.; Cheng, Y.X.; Xu, W.B.; Xiao, Y.; Chen, D.Y.; Dong, W.R.; Shu, M.A. The health risk for consumers under heavy metal scenarios: Reduce bioaccumulation of Cd in estuary mud crab (Scylla paramamosain) through the antagonism of Se. Sci. Total Environ. 2022, 844, 157149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Qian, Z.; Lu, W.; Ding, H.; Yu, H.; Wang, H.; Li, J. Identification and characterizationof reference genes for normalizing expression data from red swamp crawfish Procambarus clarkii. Int. J. Mol. Sci. 2015, 16, 21591–21605. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequences (5′-3′) | Product Size (bp) | Correlation Coefficient (R2) | Amplification Efficiency (%) |
|---|---|---|---|---|
| β-ACTIN | F:GCATCCACGAGACCACTTACA R:CTCCTGCTTGCTGATCCACATC | 266 | 0.9937 | 106.7% |
| EF-1α | F:AGGTCGGCTACAACCCAACT R:TGAACTCGTAGGAGCCACTC | 138 | 0.9908 | 104% |
| EF-2 | F:TGATGGGTCGCTTTGTTGAG R:GGTCAGATGGGTTCTTGGGT | 192 | 0.9908 | 104.3% |
| HSP90 | F:TCACCAACGACTGGGAGGAT R:CAGGAAGAGGAGTGCCCTGA | 83 | 0.9918 | 104.5% |
| AK | F:GCTCAAGGCCAAGAAGACCA R:CATACACACCGACGCCAGAG | 92 | 0.9947 | 108.1% |
| UBE | F:TTGCGTTCACAACTCGTATCTACC R:GTCCGTGAGGAGGGAACAGA | 137 | 0.9927 | 107.3% |
| GST | F:GCTGTGGTGGAGCGACTCA R:TCCAACTCCTCTCCACGGAA | 98 | 0.9923 | 95.7% |
| GAPDH | F:GCGTGTTCACCACCATTGAG R:ACATGGGTGCATCAGCAGAG | 93 | 0.9979 | 100% |
| α-TUB | F:GTGGAGATCTGGCCAAGGTG R:CCCACATACCAGTGCACGAA | 136 | 0.9986 | 101% |
| PGM2 | F:CTGACGGGCTTCAAGTGGAT R:TCCTTGTCCAACACCTCCGA | 122 | 0.9931 | 107.2% |
| EsPrx4 | F:CACGAAAGGAAGGTGGACTG R:TCATCCACAGACCTTCCGACT | 193 | 0.9973 | 107.7% |
| Gene Name | Vibrio anguillarum | Copper Ions |
|---|---|---|
| EF-1α | 0.574 | 1.056 |
| AK | 0.500 | 1.465 |
| EF-2 | 1.133 | 1.094 |
| UBE | 0.590 | 1.381 |
| β-ACTIN | 1.212 | 0.436 |
| GST | 1.372 | 1.286 |
| GAPDH | 0.737 | 0.334 |
| α-TUB | 0.833 | 0.661 |
| HSP90 | 1.714 | 1.589 |
| PGM2 | 1.368 | 1.346 |
| Gene Name | Vibrio anguillarum | Copper Ion | ||
|---|---|---|---|---|
| SD (±Ct) | CV (%Ct) | SD (±Ct) | CV (%Ct) | |
| AK | 1.12 | 5.48 | 0.67 | 3.15 |
| UBE | 1.48 | 6.22 | 1.76 | 6.97 |
| GST | 2.05 | 7.98 | 1.54 | 5.71 |
| GAPDH | 1.13 | 4.98 | 1.19 | 5.16 |
| EF-1α | 1.23 | 6.34 | 1.66 | 7.78 |
| α-TUB | 1.11 | 4.86 | 0.79 | 3.38 |
| HSP90 | 2.14 | 8.89 | 1.51 | 5.57 |
| β-ACTIN | 1.16 | 5.52 | 1.14 | 4.64 |
| EF-2 | 1.56 | 7.4 | 1.65 | 7.71 |
| PGM2 | 2.15 | 7.2 | 0.58 | 1.98 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yan, F.; Li, H.; Chen, X.; Yu, J.; Su, S.; Li, J.; Ye, W.; Tang, Y. Screening of Suitable Reference Genes for Immune Gene Expression Analysis Stimulated by Vibrio anguillarum and Copper Ions in Chinese Mitten Crab (Eriocheir sinensis). Genes 2023, 14, 1099. https://doi.org/10.3390/genes14051099
Yan F, Li H, Chen X, Yu J, Su S, Li J, Ye W, Tang Y. Screening of Suitable Reference Genes for Immune Gene Expression Analysis Stimulated by Vibrio anguillarum and Copper Ions in Chinese Mitten Crab (Eriocheir sinensis). Genes. 2023; 14(5):1099. https://doi.org/10.3390/genes14051099
Chicago/Turabian StyleYan, Fengyuan, Hui Li, Xue Chen, Junjie Yu, Shengyan Su, Jianlin Li, Wei Ye, and Yongkai Tang. 2023. "Screening of Suitable Reference Genes for Immune Gene Expression Analysis Stimulated by Vibrio anguillarum and Copper Ions in Chinese Mitten Crab (Eriocheir sinensis)" Genes 14, no. 5: 1099. https://doi.org/10.3390/genes14051099
APA StyleYan, F., Li, H., Chen, X., Yu, J., Su, S., Li, J., Ye, W., & Tang, Y. (2023). Screening of Suitable Reference Genes for Immune Gene Expression Analysis Stimulated by Vibrio anguillarum and Copper Ions in Chinese Mitten Crab (Eriocheir sinensis). Genes, 14(5), 1099. https://doi.org/10.3390/genes14051099