
CRISPR/Cas9-Mediated Knockout of tnfaip1 in Zebrafish Plays a Role in Early Development


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Zebrafish Husbandry
2.2. Whole Mount In Situ Hybridization
2.3. Generation of tnfaip1 Mutants
2.4. Transcriptome Sequencing and Data Analysis
2.5. Quantitative Real-Time PCR
2.6. Protein Extraction and Western Blotting
2.7. Embryo Imaging
2.8. Statistical Analysis
3. Results
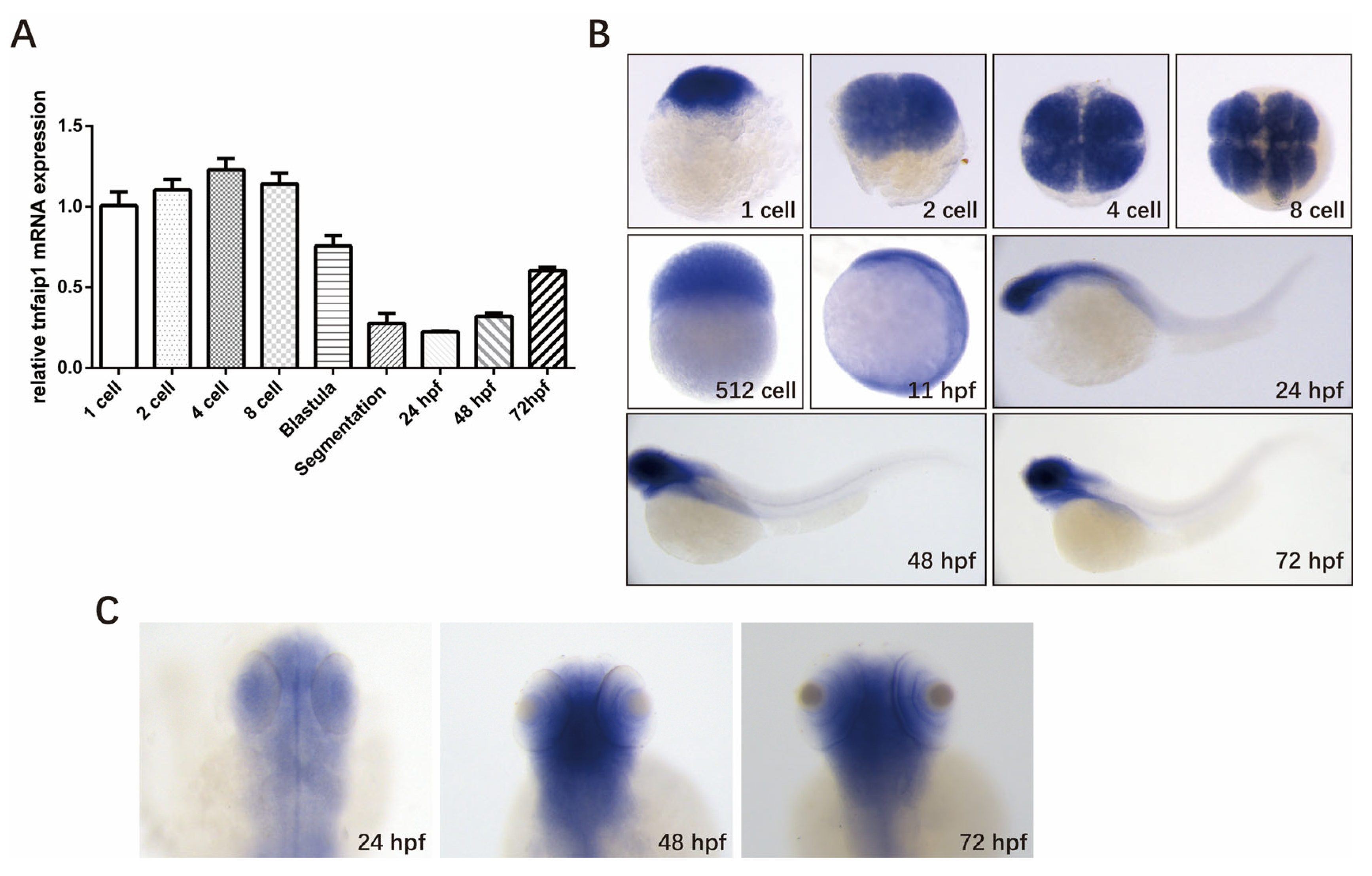
3.1. Spatio-Temporal Expression Pattern of tnfaip1 in Zebrafish during Early Development
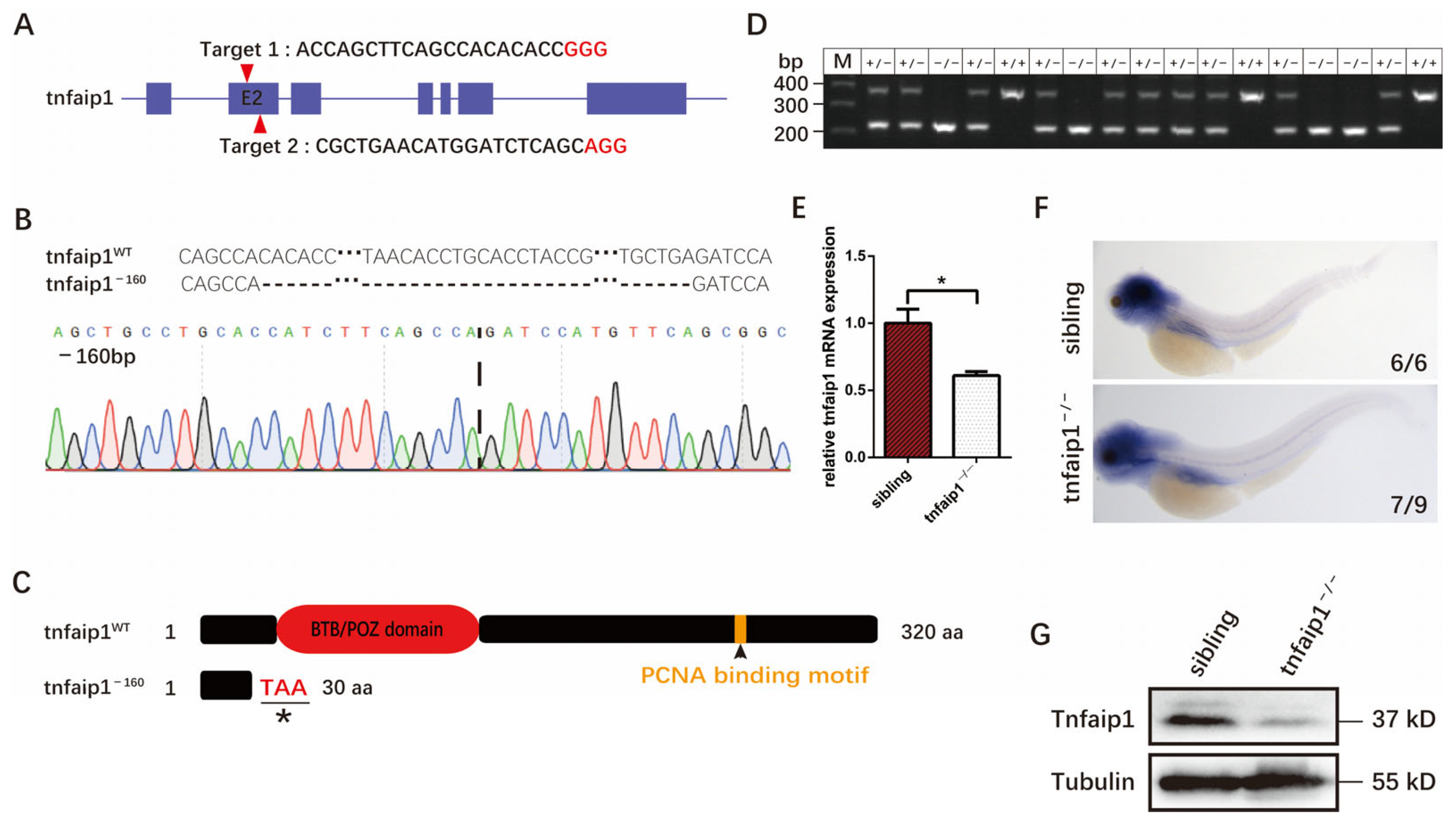
3.2. Generation of the tnfaip1 Mutant in Zebrafish Using the CRISPR/Cas9 System
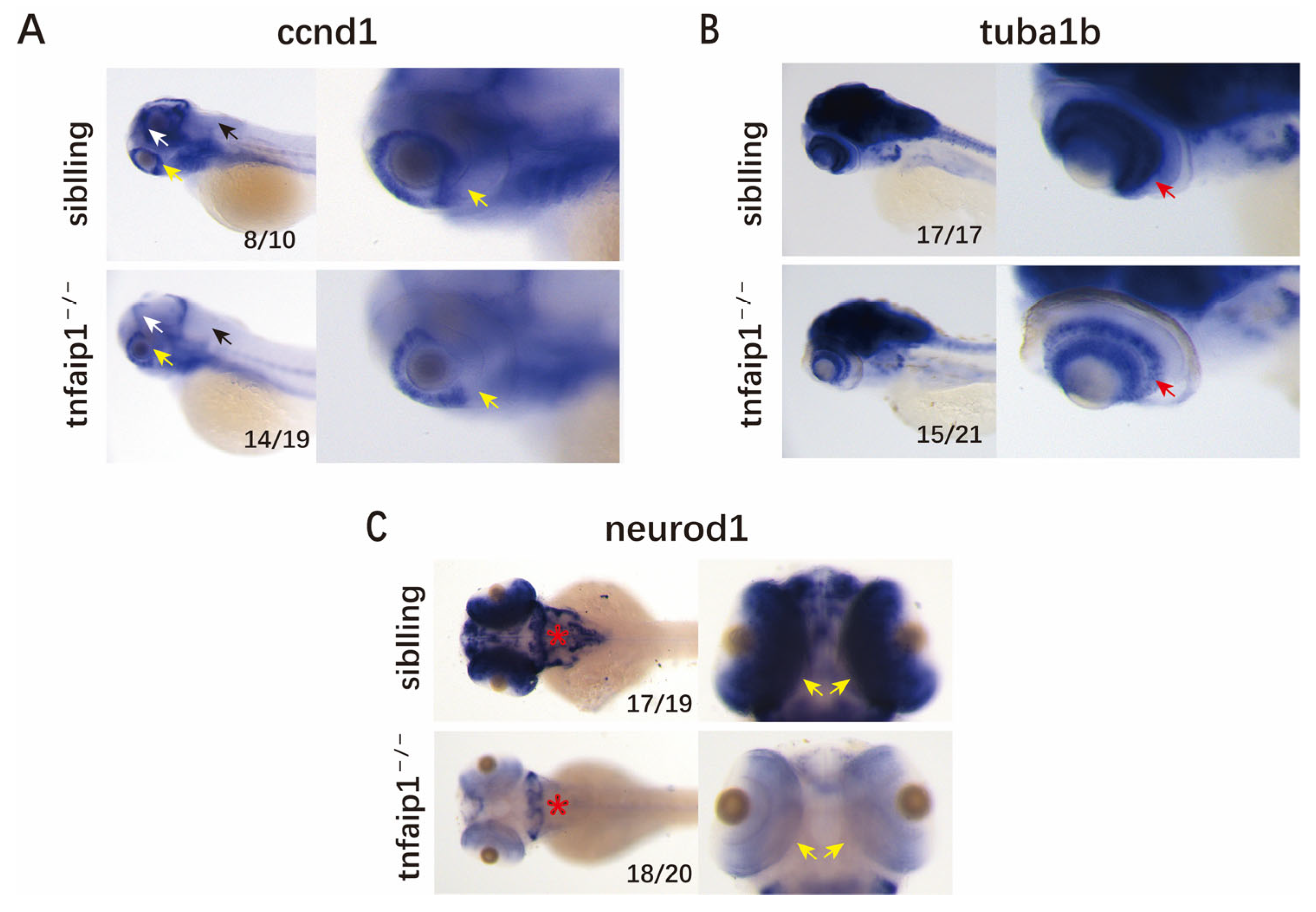
3.3. Mutation of tnfaip1 Affects the Early Development of Zebrafish Embryos
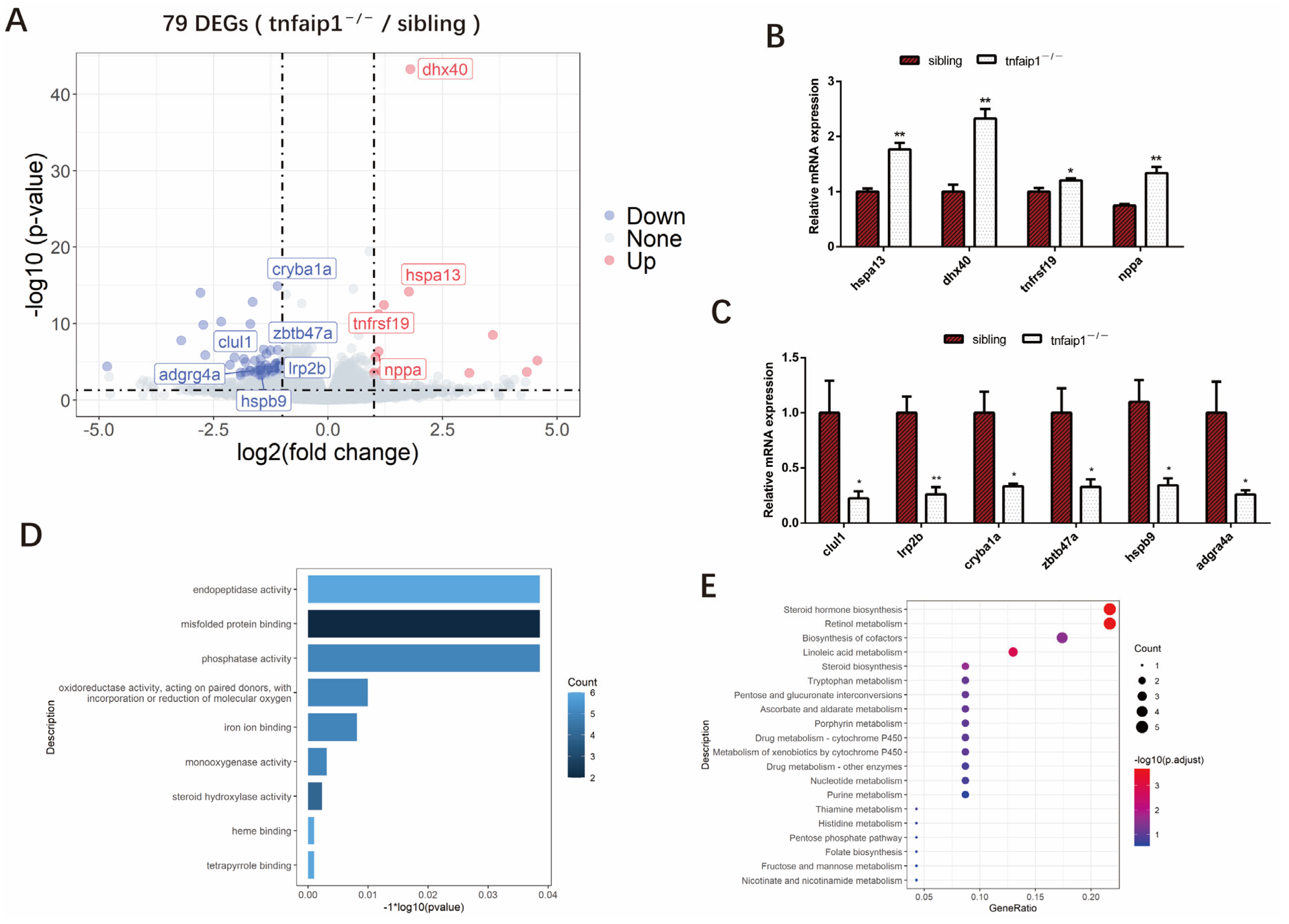
3.4. Transcriptome Levels Are Altered in tnfaip1 Mutants
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wolfs, F.W.; Marks, R.M.; Sarma, V.; Byers, M.G.; Katz, R.W.; Shows, T.B.; Dixit, V.M. Characterization of a Novel Tumor Necrosis Factor-a-induced Endothelial Primary Response Gene. J. Biol. Chem. 1992, 267, 1317–1326. [Google Scholar] [CrossRef]
- Liu, Z.; Xiang, Y.; Sun, G. The KCTD family of proteins: Structure, function, disease relevance. Cell Biosci. 2013, 3, 5. [Google Scholar] [CrossRef]
- Sloan, D.C.; Cryan, C.E.; Muntean, B.S. Multiple potassium channel tetramerization domain (KCTD) family members interact with Gbetagamma, with effects on cAMP signaling. J. Biol. Chem. 2023, 299, 102924. [Google Scholar] [CrossRef]
- Muntean, B.S.; Marwari, S.; Li, X.; Sloan, D.C.; Young, B.D.; Wohlschlegel, J.A.; Martemyanov, K.A. Members of the KCTD family are major regulators of cAMP signaling. Proc. Natl. Acad. Sci. USA 2022, 119, e2119237119. [Google Scholar] [CrossRef] [PubMed]
- Lin, G.N.; Corominas, R.; Lemmens, I.; Yang, X.; Tavernier, J.; Hill, D.E.; Vidal, M.; Sebat, J.; Iakoucheva, L.M. Spatiotemporal 16p11.2 protein network implicates cortical late mid-fetal brain development and KCTD13-Cul3-RhoA pathway in psychiatric diseases. Neuron 2015, 85, 742–754. [Google Scholar] [CrossRef] [PubMed]
- Golzio, C.; Willer, J.; Talkowski, M.E.; Oh, E.C.; Taniguchi, Y.; Jacquemont, S.; Reymond, A.; Sun, M.; Sawa, A.; Gusella, J.F.; et al. KCTD13 is a major driver of mirrored neuroanatomical phenotypes of the 16p11.2 copy number variant. Nature 2012, 485, 363–367. [Google Scholar] [CrossRef]
- Hu, X.; Gan, S.; Xie, G.; Li, L.; Chen, C.; Ding, X.; Han, M.; Xiang, S.; Zhang, J. KCTD10 is critical for heart and blood vessel development of zebrafish. Acta Biochim. Biophys. Sin. 2014, 46, 377–386. [Google Scholar] [CrossRef]
- Tong, X.; Zu, Y.; Li, Z.; Li, W.; Ying, L.; Yang, J.; Wang, X.; He, S.; Liu, D.; Zhu, Z.; et al. Kctd10 regulates heart morphogenesis by repressing the transcriptional activity of Tbx5a in zebrafish. Nat. Commun. 2014, 5, 3153. [Google Scholar] [CrossRef]
- Ren, K.; Yuan, J.; Yang, M.; Gao, X.; Ding, X.; Zhou, J.; Hu, X.; Cao, J.; Deng, X.; Xiang, S.; et al. KCTD10 is involved in the cardiovascular system and Notch signaling during early embryonic development. PLoS ONE 2014, 9, e112275. [Google Scholar] [CrossRef]
- Zhou, J.; Hu, X.; Xiong, X.; Liu, X.; Liu, Y.; Ren, K.; Jiang, T.; Hu, X.; Zhang, J. Cloning of two rat PDIP1 related genes and their interactions with proliferating cell nuclear antigen. J. Exp. Zool. Part A Comp. Exp. Biol. 2005, 303, 227–240. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Tan, Z.; Liu, X.; Yang, C.; Ding, X.; Hu, X.; Zhou, J.; Xiang, S.; Zhou, C.; et al. MicroRNA-373 is upregulated and targets TNFAIP1 in human gastric cancer, contributing to tumorigenesis. Oncol. Lett. 2013, 6, 1427–1434. [Google Scholar] [CrossRef]
- Kim, D.M.; Chung, K.S.; Choi, S.J.; Jung, Y.J.; Park, S.K.; Han, G.H.; Ha, J.S.; Song, K.B.; Choi, N.S.; Kim, H.M.; et al. RhoB induces apoptosis via direct interaction with TNFAIP1 in HeLa cells. Int. J. Cancer 2009, 125, 2520–2527. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Li, X.; Zhang, X.; Liu, X.; Tan, Z.; Yang, C.; Zhang, J. microRNA-372 maintains oncogene characteristics by targeting TNFAIP1 and affects NFkappaB signaling in human gastric carcinoma cells. Int. J. Oncol. 2013, 42, 635–642. [Google Scholar] [CrossRef] [PubMed]
- Ryu, T.Y.; Kim, K.; Han, T.S.; Lee, M.O.; Lee, J.; Choi, J.; Jung, K.B.; Jeong, E.J.; An, D.M.; Jung, C.R.; et al. Human gut-microbiome-derived propionate coordinates proteasomal degradation via HECTD2 upregulation to target EHMT2 in colorectal cancer. ISME J. 2022, 16, 1205–1221. [Google Scholar] [CrossRef] [PubMed]
- Tan, Z.W.; Xie, S.; Hu, S.Y.; Liao, T.; Liu, P.; Peng, K.H.; Yang, X.Z.; He, Z.L.; Tang, H.Y.; Cui, Y.; et al. Caudatin targets TNFAIP1/NF-kappaB and cytochrome c/caspase signaling to suppress tumor progression in human uterine cancer. Int. J. Oncol. 2016, 49, 1638–1650. [Google Scholar] [CrossRef] [PubMed]
- Cui, R.; Meng, W.; Sun, H.-L.; Kim, T.; Ye, Z.; Fassan, M.; Jeon, Y.-J.; Li, B.; Vincenti, C.; Peng, Y.; et al. MicroRNA-224 promotes tumor progression in nonsmall cell lung cancer. Proc. Natl. Acad. Sci. USA 2015, 112, E4288–E4297. [Google Scholar] [CrossRef]
- Zhang, M.; Gao, C.; Yang, Y.; Li, G.; Dong, J.; Ai, Y.; Ma, Q.; Li, W. MiR-424 Promotes Non-Small Cell Lung Cancer Progression and Metastasis through Regulating the Tumor Suppressor Gene TNFAIP1. Cell. Physiol. Biochem. 2017, 42, 211–221. [Google Scholar] [CrossRef]
- Xiao, Y.; Huang, S.; Qiu, F.; Ding, X.; Sun, Y.; Wei, C.; Hu, X.; Wei, K.; Long, S.; Xie, L.; et al. Tumor necrosis factor alpha-induced protein 1 as a novel tumor suppressor through selective downregulation of CSNK2B blocks nuclear factor-kappaB activation in hepatocellular carcinoma. eBioMedicine 2020, 51, 102603. [Google Scholar] [CrossRef]
- Liu, Y.; Zhang, W.; Wang, S.; Cai, L.; Jiang, Y.; Pan, Y.; Liang, Y.; Xian, J.; Jia, L.; Li, L.; et al. Cullin3-TNFAIP1 E3 Ligase Controls Inflammatory Response in Hepatocellular Carcinoma Cells via Ubiquitination of RhoB. Front. Cell Dev. Biol. 2021, 9, 617134. [Google Scholar] [CrossRef]
- Lin, M.C.; Lee, N.P.; Zheng, N.; Yang, P.-H.; Wong, O.G.; Kung, H.-F.; Hui, C.-K.; Luk, J.M.; Lau, G.K.-K. Tumor necrosis factor-α-induced protein 1 and immunity to hepatitis B virus. World J. Gastroenterol. 2005, 11, 7564–7568. [Google Scholar] [CrossRef]
- Link, C.D.; Taft, A.; Kapulkin, V.; Duke, K.; Kim, S.; Fei, Q.; Wood, D.E.; Sahagan, B.G. Gene expression analysis in a transgenic Caenorhabditis elegans Alzheimer's disease model. Neurobiol. Aging 2003, 24, 397–413. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yu, Z.; Xun, Y.; Li, M.; Peng, X.; Xiao, Y.; Hu, X.; Sun, Y.; Yang, M.; Gan, S.; et al. TNFAIP1 contributes to the neurotoxicity induced by Abeta25-35 in Neuro2a cells. BMC Neurosci. 2016, 17, 51. [Google Scholar] [CrossRef] [PubMed]
- He, D.; Tan, J.; Zhang, J. miR-137 attenuates Abeta-induced neurotoxicity through inactivation of NF-kappaB pathway by targeting TNFAIP1 in Neuro2a cells. Biochem. Biophys. Res. Commun. 2017, 490, 941–947. [Google Scholar] [CrossRef]
- Li, Y.; Jin, L.; Wang, F.; Ren, L.; Pen, R.; Bo, G.; Wang, L. Epigenetic axis of SNHG19/miR-137/TNFAIP1 modulates amyloid beta peptide 25-35-induced SH-SY5Y cytotoxicity. Epigenomics 2022, 14, 187–198. [Google Scholar] [CrossRef]
- Xiong, L.; Zhang, J.; Shi, H.; Zhu, G.; Ji, X.; Li, M.; Zhu, P.; Luo, K. Downregulation of TNFAIP1 alleviates OGD/R-induced neuronal damage by suppressing Nrf2/GPX4-mediated ferroptosis. Exp. Ther. Med. 2023, 25, 25. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Li, Y.; Zhang, H.; Yang, L.; Jiang, Y.; Wei, C.; Feng, X.; Xun, Y.; Yuan, S.; Xiang, S.; et al. TNFAIP1 Is Upregulated in APP/PS1 Mice and Promotes Apoptosis in SH-SY5Y Cells by Binding to RhoB. J. Mol. Neurosci. MN 2021, 71, 1221–1233. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, S.; Xia, N.; Shi, Y.; Zhao, C.M. Effects of XIST/miR-137 axis on neuropathic pain by targeting TNFAIP1 in a rat model. J. Cell. Physiol. 2018, 233, 4307–4316. [Google Scholar] [CrossRef]
- Gladwyn-Ng, I.E.; Li, S.S.; Qu, Z.; Davis, J.M.; Ngo, L.; Haas, M.; Singer, J.; Heng, J.I. Bacurd2 is a novel interacting partner to Rnd2 which controls radial migration within the developing mammalian cerebral cortex. Neural Dev. 2015, 10, 9. [Google Scholar] [CrossRef]
- Gladwyn-Ng, I.; Huang, L.; Ngo, L.; Li, S.S.; Qu, Z.; Vanyai, H.K.; Cullen, H.D.; Davis, J.M.; Heng, J.I. Bacurd1/Kctd13 and Bacurd2/Tnfaip1 are interacting partners to Rnd proteins which influence the long-term positioning and dendritic maturation of cerebral cortical neurons. Neural Dev. 2016, 11, 7. [Google Scholar] [CrossRef]
- Varshney, G.K.; Carrington, B.; Pei, W.; Bishop, K.; Chen, Z.; Fan, C.; Xu, L.; Jones, M.; LaFave, M.C.; Ledin, J.; et al. A high-throughput functional genomics workflow based on CRISPR/Cas9-mediated targeted mutagenesis in zebrafish. Nat. Protoc. 2016, 11, 2357–2375. [Google Scholar] [CrossRef]
- Westerfield, M. The Zebrafish Book. A Guide for the Laboratory Use of Zebrafish (Danio rerio); Univeristy of Oregon Press: Eugene, OR, USA, 2000. [Google Scholar]
- Thisse, C.; Thisse, B. High-resolution in situ hybridization to whole-mount zebrafish embryos. Nat. Protoc. 2008, 3, 59–69. [Google Scholar] [CrossRef]
- Gagnon, J.A.; Valen, E.; Thyme, S.B.; Huang, P.; Ahkmetova, L.; Pauli, A.; Montague, T.G.; Zimmerman, S.; Richter, C.; Schier, A.F. Efficient Mutagenesis by Cas9 Protein-Mediated Oligonucleotide Insertion and Large-Scale Assessment of Single-Guide RNAs. PLoS ONE 2014, 9, e98186. [Google Scholar] [CrossRef] [PubMed]
- Dobin, A.; Davis, C.A.; Schlesinger, F.; Drenkow, J.; Zaleski, C.; Jha, S.; Batut, P.; Chaisson, M.; Gingeras, T.R. STAR: Ultrafast universal RNA-seq aligner. Bioinformatics 2013, 29, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Dewey, C.N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome. BMC Bioinform. 2011, 12, 323. [Google Scholar] [CrossRef] [PubMed]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Kurosaki, T.; Popp, M.W.; Maquat, L.E. Quality and quantity control of gene expression by nonsense-mediated mRNA decay. Nat. Rev. Mol. Cell Biol. 2019, 20, 406–420. [Google Scholar] [CrossRef] [PubMed]
- Nasif, S.; Contu, L.; Muhlemann, O. Beyond quality control: The role of nonsense-mediated mRNA decay (NMD) in regulating gene expression. Semin. Cell Dev. Biol. 2018, 75, 78–87. [Google Scholar] [CrossRef]
- Zhang, Q.; Ray, K.; Acland, G.M.; Czarnecki, J.M.; Aguirre, G.D. Molecular cloning, characterization and expression of a novel retinal clusterin-like protein cDNA. Gene 2000, 243, 151–160. [Google Scholar] [CrossRef]
- Zhang, Q.; Beltran, W.A.; Mao, Z.; Li, K.; Johnson, J.L.; Acland, G.M.; Aguirre, G.D. Comparative analysis and expression of CLUL1, a cone photoreceptor-specific gene. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4542–4549. [Google Scholar] [CrossRef]
- Appelbaum, T.; Murgiano, L.; Becker, D.; Santana, E.; Aguirre, G.D. Candidate Genetic Modifiers for RPGR Retinal Degeneration. Investig. Ophthalmol. Vis. Sci. 2020, 61, 20. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Zhang, H.; Chen, W.; Su, N.; Yuan, C.; Zhang, J.; Xiang, S.; Hu, X. CRISPR/Cas9-Mediated Knockout of tnfaip1 in Zebrafish Plays a Role in Early Development. Genes 2023, 14, 1005. https://doi.org/10.3390/genes14051005
Huang S, Zhang H, Chen W, Su N, Yuan C, Zhang J, Xiang S, Hu X. CRISPR/Cas9-Mediated Knockout of tnfaip1 in Zebrafish Plays a Role in Early Development. Genes. 2023; 14(5):1005. https://doi.org/10.3390/genes14051005
Chicago/Turabian StyleHuang, Shulan, Hongning Zhang, Wen Chen, Na Su, Changyue Yuan, Jian Zhang, Shuanglin Xiang, and Xiang Hu. 2023. "CRISPR/Cas9-Mediated Knockout of tnfaip1 in Zebrafish Plays a Role in Early Development" Genes 14, no. 5: 1005. https://doi.org/10.3390/genes14051005
APA StyleHuang, S., Zhang, H., Chen, W., Su, N., Yuan, C., Zhang, J., Xiang, S., & Hu, X. (2023). CRISPR/Cas9-Mediated Knockout of tnfaip1 in Zebrafish Plays a Role in Early Development. Genes, 14(5), 1005. https://doi.org/10.3390/genes14051005