
Improved RAPD Method for Candida parapsilosis Fingerprinting
 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Strains
2.2. DNA Extraction
2.3. Primers
2.4. Optimization of RAPD-PCR Reaction
2.5. Calculation of Discrimination Power of RAPD-PCR
2.6. RAPD-PCR Conditions
2.7. Gel Electrophoresis
2.8. Analysis of the Amplification Results
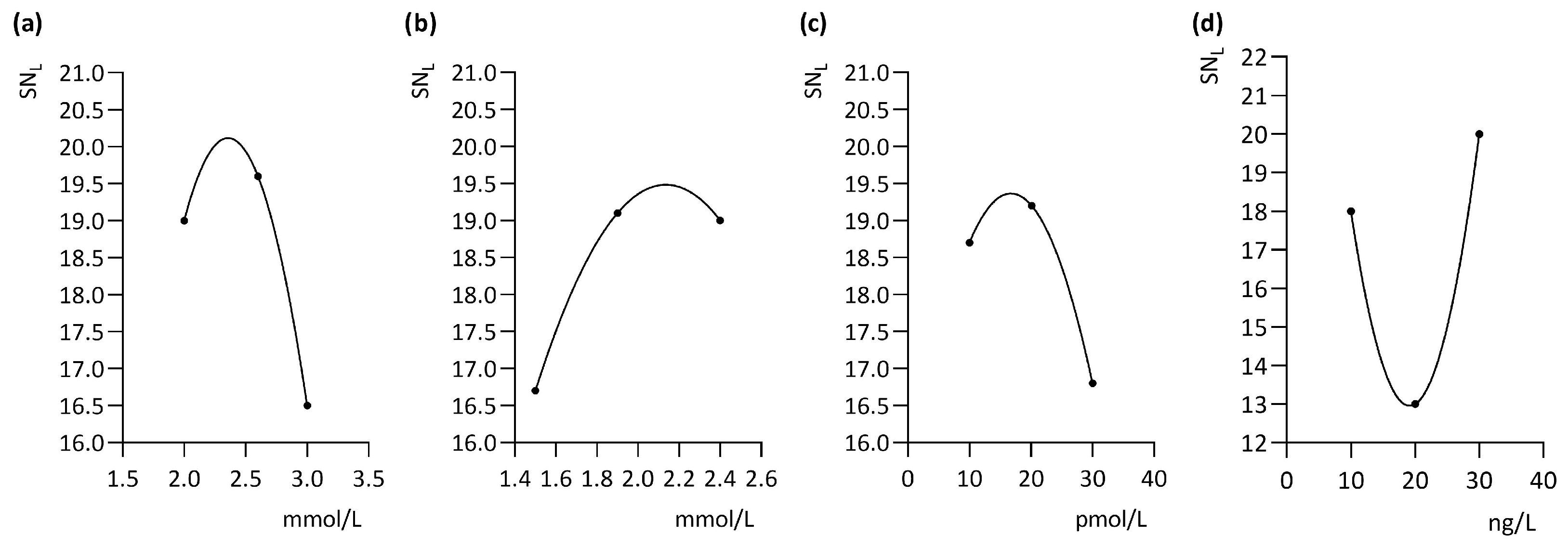
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Micheli, M.R.; Bova, R.; Calissano, P.; D’Ambrosio, E. Randomly amplified polymorphic DNA fingerprinting using combinations of oligonucleotide primers. Biotechniques 1993, 15, 388–390. [Google Scholar] [PubMed]
- Reinoso, E.B.; Bettera, S.G. Random amplified polymorphic DNA PCR in the teaching of molecular epidemiology. Biochem. Mol. Biol. Educ. 2016, 44, 391–396. [Google Scholar] [CrossRef] [PubMed]
- Cobb, B.D.; Clarkson, J.M. A simple procedure for optimising the polymerase chain reaction (PCR) using modified Taguchi methods. Nucleic Acids Res. 1994, 22, 3801–3805. [Google Scholar] [CrossRef]
- Williams, D.W.; Wilson, M.J.; Lewis, M.A. Deoxyribonucleic acid typing methods for medically important microorganisms. Br. J. Biomed. Sci. 1999, 56, 56–65. [Google Scholar]
- Steffan, P.; Vazquez, J.A.; Boikov, D.; Xu, C.; Sobel, J.D.; Akins, R.A. Identification of Candida species by randomly amplified polymorphic DNA fingerprinting of colony lysates. J. Cinical Microbiol. 1997, 35, 2031–2039. [Google Scholar] [CrossRef]
- Dabrowski, W.; Czekajlo-Kolodziej, U.; Medrala, D.; Giedrys-Kalemba, S. Optimisation of AP-PCR fingerprinting discriminatory power for clinical isolates of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 2003, 218, 51–57. [Google Scholar] [CrossRef]
- Power, E.G. RAPD typing in microbiology—a technical review. J. Hosp. Infect. 1996, 34, 247–265. [Google Scholar] [CrossRef]
- Welsh, J.; McClelland, M. Fingerprinting genomes using PCR with arbitrary primers. Nucleic Acids Res. 1990, 18, 7213–7218. [Google Scholar] [CrossRef] [PubMed]
- Williams, J.G.; Kubelik, A.R.; Livak, K.J.; Rafalski, J.A.; Tingey, S.V. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers. Nucleic Acids Res. 1990, 18, 6531–6535. [Google Scholar] [CrossRef]
- Williams, J.G.; Hanafey, M.K.; Rafalski, J.A.; Tingey, S.V. Genetic analysis using random amplified polymorphic DNA markers. Methods Enzymol. 1993, 218, 704–740. [Google Scholar] [CrossRef] [PubMed]
- Stefańska, I.; Kwiecień, E.; Górzyńska, M.; Sałamaszyńska-Guz, A.; Rzewuska, M. RAPD-PCR-based fingerprinting method as a tool for epidemiological analysis of Trueperella pyogenes infections. Pathogens 2022, 11, 562. [Google Scholar] [CrossRef]
- Mesini, A.; Mikulska, M.; Giacobbe, D.R.; Del Puente, F.; Gandolfo, N.; Codda, G.; Orsi, A.; Tassinari, F.; Beltramini, S.; Marchese, A.; et al. Changing epidemiology of candidaemia: Increase in fluconazole-resistant Candida parapsilosis. Mycoses 2020, 63, 361–368. [Google Scholar] [CrossRef]
- Khalili, Y.; Memar, M.Y.; Farajnia, S.; Adibkia, K.; Kafil, H.S.; Ghotaslou, R. Molecular epidemiology and carbapenem resistance of Pseudomonas aeruginosa isolated from patients with burns. J. Wound Care 2021, 30, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Lischewski, A.; Harmsen, D.; Hacker, J.; Morschhäuser, J. Standardized molecular typing of Candida albicans strains. Mycoses 1997, 40, 369–372. [Google Scholar] [CrossRef] [PubMed]
- Dassanayake, R.S.; Samaranayake, Y.H.; Samaranayake, L.P. Genomic diversity of oral Candida krusei isolates as revealed by DNA fingerprinting and electrophoretic karyotyping. APMIS 2000, 108, 697–704. [Google Scholar] [CrossRef]
- Di Francesco, L.F.; Barchiesi, F.; Caselli, F.; Cirioni, O.; Scalise, G. Comparison of four methods for DNA typing of clinical isolates of Candida glabrata. J. Med. Microbiol. 1999, 48, 955–963. [Google Scholar] [CrossRef]
- Olive, D.M.; Bean, P. Principles and applications of methods for DNA-based typing of microbial organisms. J. Clin. Microbiol. 1999, 37, 1661–1669. [Google Scholar] [CrossRef]
- Singh, J.P.; Verma, R.; Chaudhuri, P. Random amplified polymorphic DNA (RAPD) analysis of Mycobacterium tuberculosis strains in India. J. Vet. Sci. 2006, 7, 181–187. [Google Scholar] [CrossRef][Green Version]
- Bogiel, T.; Mikucka, A.; Kanarek, P. Agarose gel electrophoresis-based RAPD-PCR- An optimization of the conditions to rapidly detect similarity of the alert pathogens for the purpose of epidemiological studies. Gels 2022, 8, 760. [Google Scholar] [CrossRef]
- Neela, V.; Mariana, N.S.; Radu, S.; Zamberi, S.; Raha, A.R.; Rosli, R. Use of RAPD to investigate the epidemiology of Staphylococcus aureus infection in Malaysian hospitals. World J. Microbiol. Biotechnol. 2005, 21, 245–251. [Google Scholar] [CrossRef]
- Chuma, I.S.; Nonga, H.E.; Mdegela, R.H.; Kazwala, R.R. Epidemiology and RAPD-PCR typing of thermophilic campylobacters from children under five years and chickens in Morogoro Municipality, Tanzania. BMC Infect. Dis. 2016, 16, 692. [Google Scholar] [CrossRef]
- Wojciechowska-Koszko, I.; Kwiatkowski, P.; Roszkowska, P.; Krasnodębksa-Szponder, B.; Sławiński, M.; Gabrych, A.; Giedrys-Kalemba, S.; Dołęgowska, B.; Kowalczyk, E.; Sienkiewicz, M. Genetic diversity of Candida spp. isolates colonizing twins and their family members. Pathogens 2022, 11, 1532. [Google Scholar] [CrossRef]
- Graham, G.C.; Mayers, P.; Henry, R.J. A simplified method for the preparation of fungal genomic DNA for PCR and RAPD analysis. Biotechniques 1994, 16, 48–50. [Google Scholar] [PubMed]
- Taguchi, G. Introduction to Off-Line Quality Control; Central Japan Quality Control Association: Tokyo, Japan, 1979. [Google Scholar]
- Odds, F.C.; Abbott, A.B. A simple system for the presumptive identification of Candida albicans and differentiation of strains within the species. Sabouraudia 1980, 18, 301–317. [Google Scholar] [CrossRef]
- Meunier, J.R.; Grimont, P.A. Factors affecting reproducibility of random amplified polymorphic DNA fingerprinting. Res. Microbiol. 1993, 144, 373–379. [Google Scholar] [CrossRef] [PubMed]
- Dassanayake, R.S.; Samaranayake, L.P. Characterization of the genetic diversity in superficial and systemic human isolates of Candida parapsilosis by randomly amplified polymorphic DNA (RAPD). APMIS 2000, 108, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Paffetti, D.; Barberio, C.; Casalone, E.; Cavalieri, D.; Fani, R.; Fia, G.; Mori, E.; Polsinelli, M. DNA fingerprinting by random amplified polymorphic DNA and restriction fragment length polymorphism is useful for yeast typing. Res. Microbiol. 1995, 146, 587–594. [Google Scholar] [CrossRef]
- Kubelik, A.R.; Szabo, L.J. High-GC primers are useful in RAPD analysis of fungi. Curr. Genet. 1995, 28, 384–389. [Google Scholar] [CrossRef]
- Welsh, J.; McClelland, M. Genomic fingerprinting using arbitrarily primed PCR and a matrix of pairwise combinations of primers. Nucleic Acids Res. 1991, 19, 5275–5279. [Google Scholar] [CrossRef][Green Version]
- De Toro, M.; Torres, M.J.; Maite, R.; Aznar, J. Characterization of Candida parapsilosis complex isolates. Clin. Microbiol. Infect. 2011, 17, 418–424. [Google Scholar] [CrossRef]
- Guiver, M.; Levi, K.; Oppenheim, B.A. Rapid identification of Candida species by TaqMan PCR. J. Clin. Pathol. 2001, 54, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Al-Tekreeti, A.R.A.; Al-Halbosiy, M.M.F.; Dheeb, B.I.; Hashim, A.J.; Al-Zuhairi, A.F.H.; Mohammad, F.I. Molecular identification of clinical Candida isolates by simple and randomly amplified polymorphic DNA-PCR. Arab. J. Sci. Eng. 2018, 43, 163–170. [Google Scholar] [CrossRef]
- Vrioni, G.; Matsiota-Bernard, P. Molecular typing of Candida isolates from patients hospitalized in an intensive care unit. J. Infect. 2001, 42, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Lupetti, A.; Tavanti, A.; Davini, P.; Ghelardi, E.; Corsini, V.; Merusi, I.; Boldrini, A.; Campa, M.; Senesi, S. Horizontal transmission of Candida parapsilosis candidemia in a neonatal intensive care unit. J. Clin. Microbiol. 2002, 40, 2363–2369. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Number of Reaction | Component 1 | Component 2 | Component 3 | Component 4 |
|---|---|---|---|---|
| 1. | A | A | A | A |
| 2. | A | B | B | B |
| 3. | A | C | C | C |
| 4. | B | A | B | C |
| 5. | B | B | C | A |
| 6. | B | C | A | B |
| 7. | C | A | C | B |
| 8. | C | B | A | C |
| 9. | C | C | B | A |
| Number of Bands (Base Pair) | Components | Concentrations of Tested Components | ||
|---|---|---|---|---|
| A (SNL) | B (SNL) | C (SNL) | ||
| >500 bp | MgCl2 | 6…6…6 (15.57) | 7…6…6 (15.97) | 5…5…5 (13.98) |
| dNTPs | 6…7…5 (15.32) | 6…6…5 (14.97) | 6…6…5 (14.97) | |
| Primers | 6…6…5 (14.97) | 6…7…5 (15.32) | 6…6…5 (14.97) | |
| DNA | 6…6…5 (14.97) | 6…6…5 (14.97) | 6…7…5 (15.32) | |
| <500 bp | MgCl2 | 3…4…5 (11.47) | 5…4…4 (12.60) | 5…1…4 (4.35) |
| dNTPs | 3…5…1 (4.16) | 4…4…5 (12.60) | 5…4…4 (12.60) | |
| Primers | 3…4…5 (11.47) | 4…5…4 (12.60) | 5…4…1 (4.35) | |
| DNA | 3…4…4 (11.04) | 4…4…1 (4.26) | 5…5…5 (13.98) | |
| Total | MgCl2 | 8…9…10 (19.00) | 11…9…9 (19.59) * | 9…5…8 (16.46) |
| dNTPs | 8…11…5 (16.71) | 9…9…9 (19.08) * | 10…9…8 (19.00) | |
| Primers | 8…9…9 (18.73) | 9…11…8 (19.18) * | 10…9…5 (16.84) | |
| DNA | 8…9…8 (18.39) | 9…9…5 (13.17) | 10…11…9 (19.91) * | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojciechowska-Koszko, I.; Mnichowska-Polanowska, M.; Roszkowska, P.; Sławiński, M.; Giedrys-Kalemba, S.; Dołęgowska, B.; Sienkiewicz, M.; Hukowska-Szematowicz, B.; Kwiatkowski, P. Improved RAPD Method for Candida parapsilosis Fingerprinting. Genes 2023, 14, 868. https://doi.org/10.3390/genes14040868
Wojciechowska-Koszko I, Mnichowska-Polanowska M, Roszkowska P, Sławiński M, Giedrys-Kalemba S, Dołęgowska B, Sienkiewicz M, Hukowska-Szematowicz B, Kwiatkowski P. Improved RAPD Method for Candida parapsilosis Fingerprinting. Genes. 2023; 14(4):868. https://doi.org/10.3390/genes14040868
Chicago/Turabian StyleWojciechowska-Koszko, Iwona, Magdalena Mnichowska-Polanowska, Paulina Roszkowska, Michał Sławiński, Stefania Giedrys-Kalemba, Barbara Dołęgowska, Monika Sienkiewicz, Beata Hukowska-Szematowicz, and Paweł Kwiatkowski. 2023. "Improved RAPD Method for Candida parapsilosis Fingerprinting" Genes 14, no. 4: 868. https://doi.org/10.3390/genes14040868
APA StyleWojciechowska-Koszko, I., Mnichowska-Polanowska, M., Roszkowska, P., Sławiński, M., Giedrys-Kalemba, S., Dołęgowska, B., Sienkiewicz, M., Hukowska-Szematowicz, B., & Kwiatkowski, P. (2023). Improved RAPD Method for Candida parapsilosis Fingerprinting. Genes, 14(4), 868. https://doi.org/10.3390/genes14040868