
The Association of Calcium Signaling Pathway Gene Variants, Bone Mineral Density and Mild Cognitive Impairment in Elderly People
 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects and Procedure
2.2. Measures
2.2.1. Cognitive Function
2.2.2. Bone Mineral Density
2.3. Gene Selection
2.4. SNPs Selection and Imputation
2.5. Calculation of the PRS
2.6. Statistical Analysis
3. Results
3.1. Sample Characteristics
3.2. Results of the Relationship between Gene Polymorphism and MCI
3.3. Results of the Relationship between PRS and MCI
3.4. Sensitivity Analysis
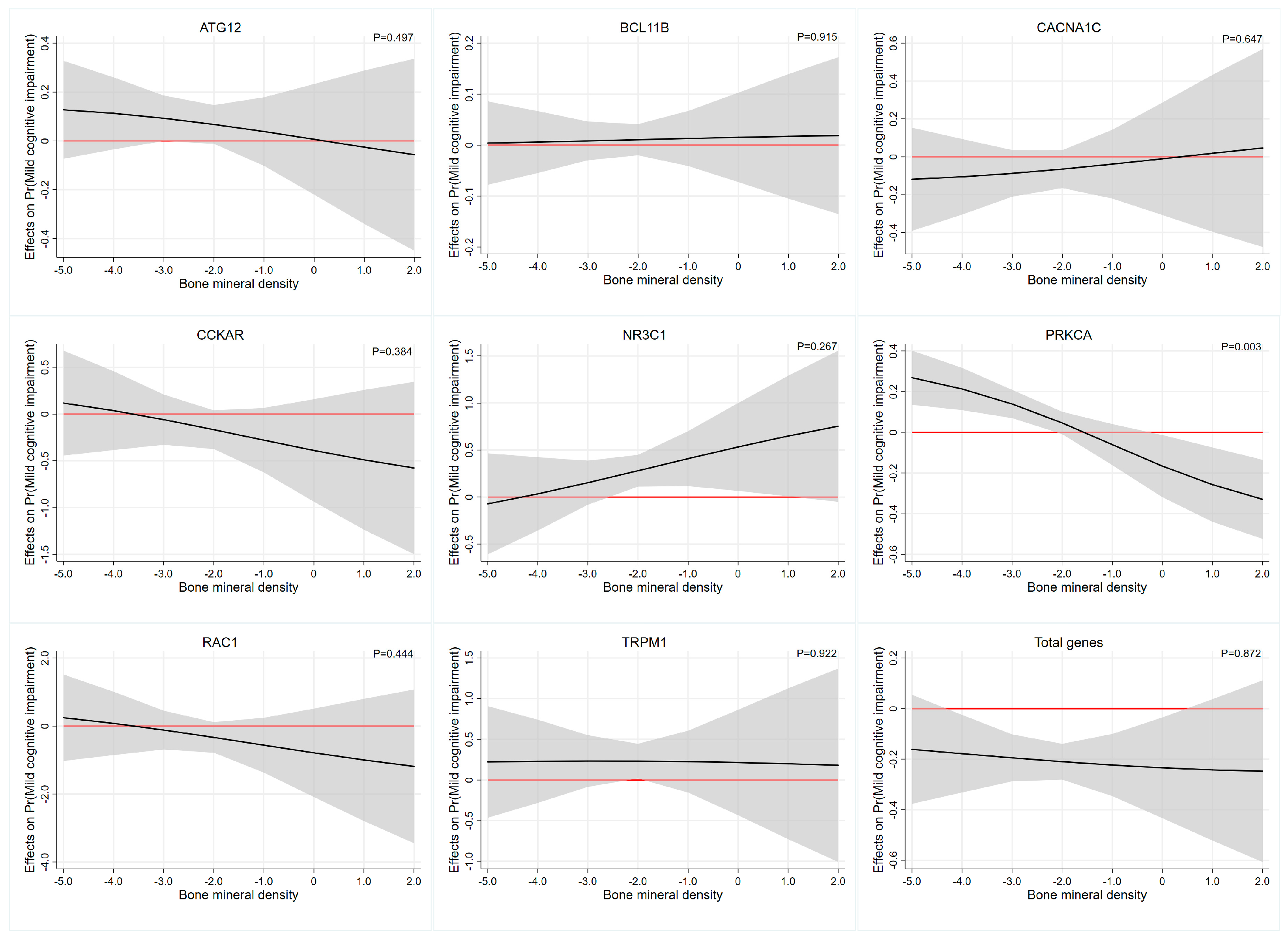
3.5. Interaction Effect Analysis between PRS and Bone Mineral Density
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Vega, J.N.; Newhouse, P.A. Mild cognitive impairment: Diagnosis, longitudinal course, and emerging treatments. Curr. Psychiatry Rep. 2014, 16, 490. [Google Scholar] [CrossRef]
- Petersen, R.C.; Lopez, O.; Armstrong, M.J.; Getchius, T.S.D.; Ganguli, M.; Gloss, D.; Gronseth, G.S.; Marson, D.; Pringsheim, T.; Day, G.S.; et al. Practice guideline update summary: Mild cognitive impairment: Report of the Guideline Development, Dissemination, and Implementation Subcommittee of the American Academy of Neurology. Neurology 2018, 90, 126–135. [Google Scholar] [CrossRef] [PubMed]
- Gavelin, H.M.; Dong, C.; Minkov, R.; Bahar-Fuchs, A.; Ellis, K.A.; Lautenschlager, N.T.; Mellow, M.L.; Wade, A.T.; Smith, A.E.; Finke, C.; et al. Combined physical and cognitive training for older adults with and without cognitive impairment: A systematic review and network meta-analysis of randomized controlled trials. Ageing Res. Rev. 2021, 66, 101232. [Google Scholar] [CrossRef] [PubMed]
- Jia, L.; Du, Y.; Chu, L.; Zhang, Z.; Li, F.; Lyu, D.; Li, Y.; Li, Y.; Zhu, M.; Jiao, H.; et al. Prevalence, risk factors, and management of dementia and mild cognitive impairment in adults aged 60 years or older in China: A cross-sectional study. Lancet Public Health 2020, 5, e661–e671. [Google Scholar] [CrossRef]
- Lissek, V.; Suchan, B. Preventing dementia? Interventional approaches in mild cognitive impairment. Neurosci. Biobehav. Rev. 2021, 122, 143–164. [Google Scholar] [CrossRef]
- Pan, G.; King, A.; Wu, F.; Simpson-Yap, S.; Woodhouse, A.; Phipps, A.; Vickers, J.C. The potential roles of genetic factors in predicting ageing-related cognitive change and Alzheimer’s disease. Ageing Res. Rev. 2021, 70, 101402. [Google Scholar] [CrossRef] [PubMed]
- Trampush, J.W.; Yang, M.L.; Yu, J.; Knowles, E.; Davies, G.; Liewald, D.C.; Starr, J.M.; Djurovic, S.; Melle, I.; Sundet, K.; et al. GWAS meta-analysis reveals novel loci and genetic correlates for general cognitive function: A report from the COGENT consortium. Mol. Psychiatry 2017, 22, 336–345. [Google Scholar] [CrossRef]
- Ohi, K.; Sumiyoshi, C.; Fujino, H.; Yasuda, Y.; Yamamori, H.; Fujimoto, M.; Shiino, T.; Sumiyoshi, T.; Hashimoto, R. Genetic Overlap between General Cognitive Function and Schizophrenia: A Review of Cognitive GWASs. Int. J. Mol. Sci. 2018, 19, 3822. [Google Scholar] [CrossRef]
- Xu, C.; Zhang, D.; Wu, Y.; Tian, X.; Pang, Z.; Li, S.; Tan, Q. A genome-wide association study of cognitive function in Chinese adult twins. Biogerontology 2017, 18, 811–819. [Google Scholar] [CrossRef]
- Berridge, M.J. Calcium signalling and psychiatric disease: Bipolar disorder and schizophrenia. Cell Tissue Res. 2014, 357, 477–492. [Google Scholar] [CrossRef]
- Sushma; Mondal, A.C. Role of GPCR signaling and calcium dysregulation in Alzheimer’s disease. Mol. Cell Neurosci. 2019, 101, 103414. [Google Scholar] [CrossRef]
- Heck, A.; Fastenrath, M.; Coynel, D.; Auschra, B.; Bickel, H.; Freytag, V.; Gschwind, L.; Hartmann, F.; Jessen, F.; Kaduszkiewicz, H.; et al. Genetic Analysis of Association Between Calcium Signaling and Hippocampal Activation, Memory Performance in the Young and Old, and Risk for Sporadic Alzheimer Disease. JAMA Psychiatry 2015, 72, 1029–1036. [Google Scholar] [CrossRef]
- Kumar, A. Calcium Signaling During Brain Aging and Its Influence on the Hippocampal Synaptic Plasticity. Adv. Exp. Med. Biol. 2020, 1131, 985–1012. [Google Scholar]
- Alzheimer’s Association Calcium Hypothesis Workgroup; Khachaturian, Z.S. Calcium Hypothesis of Alzheimer’s disease and brain aging: A framework for integrating new evidence into a comprehensive theory of pathogenesis. Alzheimers Dement. 2017, 13, 178–182.e17. [Google Scholar]
- Burgess, S.; Thompson, S.G. Use of allele scores as instrumental variables for Mendelian randomization. Int. J. Epidemiol. 2013, 42, 1134–1144. [Google Scholar] [CrossRef]
- Burgess, S.; Dudbridge, F.; Thompson, S.G. Combining information on multiple instrumental variables in Mendelian randomization: Comparison of allele score and summarized data methods. Stat. Med. 2016, 35, 1880–1906. [Google Scholar] [CrossRef]
- Carvalho, A.; Rea, I.M.; Parimon, T.; Cusack, B.J. Physical activity and cognitive function in individuals over 60 years of age: A systematic review. Clin. Interv. Aging 2014, 9, 661–682. [Google Scholar] [PubMed]
- Pérez-Sousa, M.; Del Pozo-Cruz, J.; Olivares, P.R.; Cano-Gutiérrez, C.A.; Izquierdo, M.; Ramírez-Vélez, R. Role for Physical Fitness in the Association between Age and Cognitive Function in Older Adults: A Mediation Analysis of the SABE Colombia Study. Int. J. Environ. Res. Public. Health 2021, 18, 751. [Google Scholar] [CrossRef]
- Sewell, K.R.; Erickson, K.I.; Rainey-Smith, S.R.; Peiffer, J.J.; Sohrabi, H.R.; Brown, B.M. Relationships between physical activity, sleep and cognitive function: A narrative review. Neurosci. Biobehav. Rev. 2021, 130, 369–378. [Google Scholar] [CrossRef]
- McHugh Power, J.; Tang, J.; Kenny, R.A.; Lawlor, B.A.; Kee, F. Mediating the relationship between loneliness and cognitive function: The role of depressive and anxiety symptoms. Aging Ment. Health 2020, 24, 1071–1078. [Google Scholar] [CrossRef]
- Li, W.; Sun, H.; Xu, W.; Ma, W.; Yuan, X.; Wu, H.; Kou, C. Leisure activity and cognitive function among Chinese old adults: The multiple mediation effect of anxiety and loneliness. J. Affect. Disord. 2021, 294, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, M.J.; Sachdev, P. Brain reserve and dementia: A systematic review. Psychol. Med. 2006, 36, 441–454. [Google Scholar] [CrossRef]
- Killin, L.O.; Starr, J.M.; Shiue, I.J.; Russ, T.C. Environmental risk factors for dementia: A systematic review. BMC Geriatr. 2016, 16, 175. [Google Scholar] [CrossRef]
- Zhou, R.; Deng, J.; Zhang, M.; Zhou, H.D.; Wang, Y.J. Association between bone mineral density and the risk of Alzheimer’s disease. J. Alzheimers Dis. 2011, 24, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Laudisio, A.; Fontana, D.O.; Rivera, C.; Ruggiero, C.; Bandinelli, S.; Gemma, A.; Ferrucci, L.; Antonelli Incalzi, R. Bone Mineral Density and Cognitive Decline in Elderly Women: Results from the InCHIANTI Study. Calcif. Tissue Int. 2016, 98, 479–488. [Google Scholar] [CrossRef]
- Bendayan, R.; Kuh, D.; Cooper, R.; Muthuri, S.; Muniz-Terrera, G.; Adams, J.; Ward, K.; Richards, M. Associations of Childhood and Adulthood Cognition with Bone Mineral Density in Later Adulthood: A Population-Based Longitudinal Study. Front. Aging Neurosci. 2017, 9, 241. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.G.; Park, H.Y.; Ryu, H.U.; Suk, S.H. Bone mineral loss and cognitive impairment: The PRESENT project. Medicine 2018, 97, e12755. [Google Scholar] [CrossRef]
- Nasreddine, Z.S.; Phillips, N.A.; Bédirian, V.; Charbonneau, S.; Whitehead, V.; Collin, I.; Cummings, J.L.; Chertkow, H. The Montreal Cognitive Assessment, MoCA: A brief screening tool for mild cognitive impairment. J. Am. Geriatr. Soc. 2005, 53, 695–699. [Google Scholar] [CrossRef]
- Kanis, J.A. Diagnosis of osteoporosis and assessment of fracture risk. Lancet 2002, 359, 1929–1936. [Google Scholar] [CrossRef]
- Jorgensen, T.J.; Ruczinski, I.; Kessing, B.; Smith, M.W.; Shugart, Y.Y.; Alberg, A.J. Hypothesis-driven candidate gene association studies: Practical design and analytical considerations. Am. J. Epidemiol. 2009, 170, 986–993. [Google Scholar] [CrossRef]
- Singer, J.B. Candidate gene association analysis. Methods Mol. Biol. 2009, 573, 223–230. [Google Scholar]
- Jansen, J.H.; van der Reijden, B.A. Isolation of RNA and DNA from leukocytes and cDNA synthesis. Methods Mol. Med. 2006, 125, 1–11. [Google Scholar]
- White, I.R.; Royston, P.; Wood, A.M. Multiple imputation using chained equations: Issues and guidance for practice. Stat. Med. 2011, 30, 377–399. [Google Scholar] [CrossRef]
- Lin, X.; Song, K.; Lim, N.; Yuan, X.; Johnson, T.; Abderrahmani, A.; Vollenweider, P.; Stirnadel, H.; Sundseth, S.S.; Lai, E.; et al. Risk prediction of prevalent diabetes in a Swiss population using a weighted genetic score—The CoLaus Study. Diabetologia 2009, 52, 600–608. [Google Scholar] [CrossRef]
- Barral, S.; Habeck, C.; Gazes, E.; De Jager, P.L.; Bennett, D.A.; Stern, Y. A Dopamine Receptor genetic variant enhances perceptual speed in cognitive healthy subjects. Alzheimers Dement. 2017, 3, 254–261. [Google Scholar] [CrossRef]
- Zheng, C.; Fu, Q.; Shen, Y.; Xu, Q. Investigation of allelic heterogeneity of the CCK-A receptor gene in paranoid schizophrenia. Am. J. Med. Genet. B Neuropsychiatr. Genet. 2012, 159, 741–747. [Google Scholar] [CrossRef]
- Porcelli, S.; Lee, S.J.; Han, C.; Patkar, A.A.; Serretti, A.; Pae, C.U. CACNA1C gene and schizophrenia: A case-control and pharmacogenetic study. Psychiatr. Genet. 2015, 25, 163–167. [Google Scholar] [CrossRef]
- Khalil, M.; Hollander, P.; Raucher-Chéné, D.; Lepage, M.; Lavigne, K.M. Structural brain correlates of cognitive function in schizophrenia: A meta-analysis. Neurosci. Biobehav. Rev. 2022, 132, 37–49. [Google Scholar] [CrossRef]
- Plieger, T.; Felten, A.; Splittgerber, H.; Duke, É.; Reuter, M. The role of genetic variation in the glucocorticoid receptor (NR3C1) and mineralocorticoid receptor (NR3C2) in the association between cortisol response and cognition under acute stress. Psychoneuroendocrinology 2018, 87, 173–180. [Google Scholar] [CrossRef]
- De Kloet, E.R.; Joëls, M.; Holsboer, F. Stress and the brain: From adaptation to disease. Nat. Rev. Neurosci. 2005, 6, 463–475. [Google Scholar] [CrossRef] [PubMed]
- McNerney, M.W.; Sheng, T.; Nechvatal, J.M.; Lee, A.G.; Lyons, D.M.; Soman, S.; Liao, C.P.; O’Hara, R.; Hallmayer, J.; Taylor, J.; et al. Integration of neural and epigenetic contributions to posttraumatic stress symptoms: The role of hippocampal volume and glucocorticoid receptor gene methylation. PLoS ONE 2018, 13, e0192222. [Google Scholar] [CrossRef]
- Keller, J.; Gomez, R.; Williams, G.; Lembke, A.; Lazzeroni, L.; Murphy, G.M., Jr.; Schatzberg, A.F. HPA axis in major depression: Cortisol, clinical symptomatology and genetic variation predict cognition. Mol. Psychiatry 2017, 22, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.-H. Protein kinase C (PKC) isozymes and cancer. New J. Sci. 2014, 2014, 231418. [Google Scholar] [CrossRef]
- Domínguez-García, S.; Gómez-Oliva, R.; Geribaldi-Doldán, N.; Hierro-Bujalance, C.; Sendra, M.; Ruiz, F.A.; Carrascal, L.; Macías-Sánchez, A.J.; Verástegui, C.; Hernández-Galán, R.; et al. Effects of classical PKC activation on hippocampal neurogenesis and cognitive performance: Mechanism of action. Neuropsychopharmacology 2021, 46, 1207–1219. [Google Scholar] [CrossRef]
- Choi, D.S.; Wang, D.; Yu, G.Q.; Zhu, G.; Kharazia, V.N.; Paredes, J.P.; Chang, W.S.; Deitchman, J.K.; Mucke, L.; Messing, R.O. PKCepsilon increases endothelin converting enzyme activity and reduces amyloid plaque pathology in transgenic mice. Proc. Natl. Acad. Sci. USA 2006, 103, 8215–8220. [Google Scholar] [CrossRef]
- Jablensky, A.; Morar, B.; Wiltshire, S.; Carter, K.; Dragovic, M.; Badcock, J.C.; Chandler, D.; Peters, K.; Kalaydjieva, L. Polymorphisms associated with normal memory variation also affect memory impairment in schizophrenia. Genes. Brain Behav. 2011, 10, 410–417. [Google Scholar] [CrossRef] [PubMed]
- Wilker, S.; Elbert, T.; Kolassa, I.T. The downside of strong emotional memories: How human memory-related genes influence the risk for posttraumatic stress disorder—A selective review. Neurobiol. Learn. Mem. 2014, 112, 75–86. [Google Scholar] [CrossRef]
- Lepichon, J.B.; Bittel, D.C.; Graf, W.D.; Yu, S. A 15q13.3 homozygous microdeletion associated with a severe neurodevelopmental disorder suggests putative functions of the TRPM1, CHRNA7, and other homozygously deleted genes. Am. J. Med. Genet. A 2010, 152, 1300–1304. [Google Scholar] [CrossRef]
- Hori, T.; Ikuta, S.; Hattori, S.; Takao, K.; Miyakawa, T.; Koike, C. Mice with mutations in Trpm1, a gene in the locus of 15q13.3 microdeletion syndrome, display pronounced hyperactivity and decreased anxiety-like behavior. Mol. Brain 2021, 14, 61. [Google Scholar] [CrossRef]
- Ershler, W.B. Interleukin-6: A Cytokine for Gerontolgists. J. Am. Geriatr. Soc. 1993, 41, 176–181. [Google Scholar] [CrossRef]
- Beek, K.J.; Rusman, T.; van der Weijden, M.A.C.; Lems, W.F.; van Denderen, J.C.; Konsta, M.; Visman, I.; Nurmohamed, M.T.; van der Horst-Bruinsma, I.E. Long-Term Treatment With TNF-α Inhibitors Improves Bone Mineral Density But Not Vertebral Fracture Progression in Ankylosing Spondylitis. J. Bone Min. Res. 2019, 34, 1041–1048. [Google Scholar] [CrossRef] [PubMed]
- Lopez, J.R.; Lyckman, A.; Oddo, S.; Laferla, F.M.; Querfurth, H.W.; Shtifman, A. Increased intraneuronal resting [Ca2+] in adult Alzheimer’s disease mice. J. Neurochem. 2008, 105, 262–271. [Google Scholar] [CrossRef]
- Khrimian, L.; Obri, A.; Ramos-Brossier, M.; Rousseaud, A.; Moriceau, S.; Nicot, A.S.; Mera, P.; Kosmidis, S.; Karnavas, T.; Saudou, F.; et al. Gpr158 mediates osteocalcin’s regulation of cognition. J. Exp. Med. 2017, 214, 2859–2873. [Google Scholar] [CrossRef]
- Rossi, M.; Battafarano, G.; Pepe, J.; Minisola, S.; Del Fattore, A. The Endocrine Function of Osteocalcin Regulated by Bone Resorption: A Lesson from Reduced and Increased Bone Mass Diseases. Int. J. Mol. Sci. 2019, 20, 4502. [Google Scholar] [CrossRef]
- Funk, J.L.; Mortel, K.F.; Meyer, J.S. Effects of estrogen replacement therapy on cerebral perfusion and cognition among postmenopausal women. Dement. Geriatr. Cogn. Disord. 1991, 2, 268–272. [Google Scholar] [CrossRef]


{kind=link}
| Characteristics | Normal Cognitive Status (n = 307) | Mild Cognitive Impairment (n = 571) | p-Value |
|---|---|---|---|
| Age (years) | 66.92 ± 5.01 | 69.44 ± 6.54 | <0.001 1 |
| Gender (n (%)) | <0.001 2 | ||
| Male | 164 (53.4) | 186 (32.6) | |
| Female | 143 (46.6) | 385 (67.4) | |
| Education (n (%)) | <0.001 2 | ||
| Low | 3 (1.0) | 100 (17.5) | |
| Middle | 268 (87.3) | 455 (79.7) | |
| High | 36 (11.7) | 16 (2.8) | |
| Smoking status (n (%)) | <0.001 2 | ||
| Never | 169 (55.0) | 414 (72.5) | |
| Ever | 74 (24.1) | 82 (14.4) | |
| Current | 64 (20.9) | 75 (13.1) | |
| Alcohol drinking status (n (%)) | <0.001 2 | ||
| Never | 169 (55.0) | 393 (68.8) | |
| Ever | 10 (3.3) | 24 (4.2) | |
| Current | 128 (41.7) | 154 (27.0) | |
| BMD (T-value) | −2.08 ± 0.87 | −2.40 ± 0.83 | <0.001 1 |
| SNP | Model | Genotype 1 | OR | 95%CI | p-Value | |
|---|---|---|---|---|---|---|
| Low | Upper | |||||
| rs6877893 | Allele model | A vs. G | 1.02 | 0.81 | 1.29 | 0.881 |
| Dominant model | AA vs. AG + GG | 1.14 | 0.84 | 1.55 | 0.392 | |
| Recessive model | AA + AG vs. GG | 0.52 | 0.27 | 0.99 | 0.045 * | |
| Homozygote model | AA vs. GG | 0.57 | 0.29 | 1.12 | 0.101 | |
| Heterozygote model | AA vs. AG | 1.29 | 0.93 | 1.78 | 0.129 | |
| rs6448456 | Allele model | G vs. C | 1.07 | 0.81 | 1.43 | 0.628 |
| Dominant model | GG vs. GC + CC | 1.15 | 0.82 | 1.63 | 0.419 | |
| Recessive model | GG + GC vs. CC | 0.32 | 0.11 | 0.90 | 0.031 * | |
| Homozygote model | GG vs. CC | 0.34 | 0.12 | 0.98 | 0.045 * | |
| Heterozygote model | GG vs. GC | 1.29 | 0.90 | 1.86 | 0.167 | |
| rs723672 | Allele model | T vs. C | 1.12 | 0.89 | 1.42 | 0.347 |
| Dominant model | TT vs. TC + CC | 1.30 | 0.96 | 1.77 | 0.090 | |
| Recessive model | TT + TC vs. CC | 0.72 | 0.37 | 1.41 | 0.344 | |
| Homozygote model | TT vs. CC | 0.84 | 0.43 | 1.64 | 0.615 | |
| Heterozygote model | TT vs. TC | 1.40 | 1.01 | 1.93 | 0.042 * | |
| Gene | Crude Model 1 | Multivariate-Adjusted 2 | ||
|---|---|---|---|---|
| OR (95 %CI) | p-Vaule | OR (95 %CI) | p-Vaule | |
| ATG12 | 1.589 (1.113–2.270) | 0.011 * | 1.460 (0.998–2.133) | 0.051 |
| BCL11B | 1.064 (0.930–1.217) | 0.370 | 1.052 (0.908–1.218) | 0.502 |
| CACNA1C | 0.636 (0.410–0.986) | 0.043 * | 0.697 (0.432–1.124) | 0.139 |
| CCKAR | 0.370 (0.144–0.950) | 0.039 * | 0.480 (0.174–1.328) | 0.158 |
| NR3C1 | 4.739 (2.148–10.459) | <0.001 * | 4.012 (1.722–9.347) | 0.001 * |
| PRKCA | 1.511 (1.181–1.934) | 0.001 * | 1.414 (1.083–1.845) | 0.011 * |
| RAC1 | 0.143 (0.020–1.037) | 0.054 | 0.224 (0.027–1.862) | 0.166 |
| TRPM1 | 5.477 (1.966–15.261) | 0.001 * | 3.253 (1.116–9.484) | 0.031 * |
| Total | 0.265 (0.186–0.380) | <0.001 * | 0.330 (0.224–0.485) | <0.001 * |
| Gene | Male | Female | ||
|---|---|---|---|---|
| OR (95 %CI) | p-Vaule | OR (95 %CI) | p-Vaule | |
| ATG12 | 1.429 (0.820–2.488) | 0.207 | 1.364 (0.796–2.336) | 0.258 |
| BCL11B | 1.271 (0.997–1.620) | 0.053 | 0.957 (0.792–1.155) | 0.646 |
| CACNA1C | 0.555 (0.260–1.181) | 0.126 | 0.819 (0.436–1.540) | 0.536 |
| CCKAR | 0.589 (0.125–2.784) | 0.504 | 0.321 (0.080–1.290) | 0.109 |
| NR3C1 | 2.705 (0.768–9.520) | 0.121 | 5.805 (1.840–18.309) | 0.003 * |
| PRKCA | 1.174 (0.785–1.757) | 0.434 | 1.639 (1.142–2.353) | 0.007 * |
| RAC1 | 0.119 (0.006–2.322) | 0.160 | 0.535 (0.022–13.040) | 0.701 |
| TRPM1 | 1.439 (0.358–5.778) | 0.608 | 7.869 (1.464–42.286) | 0.016 * |
| Total | 0.333 (0.001–0.041) | <0.001 * | 0.329 (0.196–0.551) | <0.001 * |
| Gene | BMD × PRS 1 | BMD × PRS 2 | ||||
|---|---|---|---|---|---|---|
| OR | 95%CI | p-Value | OR | 95%CI | p-Value | |
| ATG12 | 0.917 | 0.601–1.398 | 0.686 | 0.858 | 0.551–1.336 | 0.497 |
| BCL11B | 1.003 | 0.855–1.178 | 0.964 | 1.010 | 0.847–1.203 | 0.915 |
| CACNA1C | 1.124 | 0.652–1.938 | 0.675 | 1.148 | 0.636–2.072 | 0.647 |
| CCKAR | 0.917 | 0.310–2.710 | 0.875 | 0.594 | 0.184–1.922 | 0.384 |
| NR3C1 | 1.804 | 0.685–4.750 | 0.232 | 1.852 | 0.625–5.491 | 0.267 |
| PRKCA | 0.638 | 0.463–0.879 | 0.006 * | 0.593 | 0.418–0.841 | 0.003 * |
| RAC1 | 0.304 | 0.025–3.686 | 0.349 | 0.348 | 0.023–5.199 | 0.444 |
| TRPM1 | 1.025 | 0.274–3.835 | 0.971 | 0.932 | 0.227–3.833 | 0.922 |
| Total | 0.900 | 0.589–1.376 | 0.626 | 0.962 | 0.603–1.535 | 0.872 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Wang, X.; Duan, H.; Chen, C.; Lu, Z.; Zhang, D.; Li, S. The Association of Calcium Signaling Pathway Gene Variants, Bone Mineral Density and Mild Cognitive Impairment in Elderly People. Genes 2023, 14, 828. https://doi.org/10.3390/genes14040828
Zhang J, Wang X, Duan H, Chen C, Lu Z, Zhang D, Li S. The Association of Calcium Signaling Pathway Gene Variants, Bone Mineral Density and Mild Cognitive Impairment in Elderly People. Genes. 2023; 14(4):828. https://doi.org/10.3390/genes14040828
Chicago/Turabian StyleZhang, Jiesong, Xueyan Wang, Haiping Duan, Chen Chen, Zhonghai Lu, Dongfeng Zhang, and Suyun Li. 2023. "The Association of Calcium Signaling Pathway Gene Variants, Bone Mineral Density and Mild Cognitive Impairment in Elderly People" Genes 14, no. 4: 828. https://doi.org/10.3390/genes14040828
APA StyleZhang, J., Wang, X., Duan, H., Chen, C., Lu, Z., Zhang, D., & Li, S. (2023). The Association of Calcium Signaling Pathway Gene Variants, Bone Mineral Density and Mild Cognitive Impairment in Elderly People. Genes, 14(4), 828. https://doi.org/10.3390/genes14040828