

The Expression Patterns of Exogenous Plant miRNAs in Chickens

Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation and Small RNA Sequencing
2.2. Analysis of Illumina Sequencing Data
2.3. Animals and Gavage Feeding
2.4. Cell Culture and Exogenous miRNA Uptake Assay in Cultured Cells
3. Results
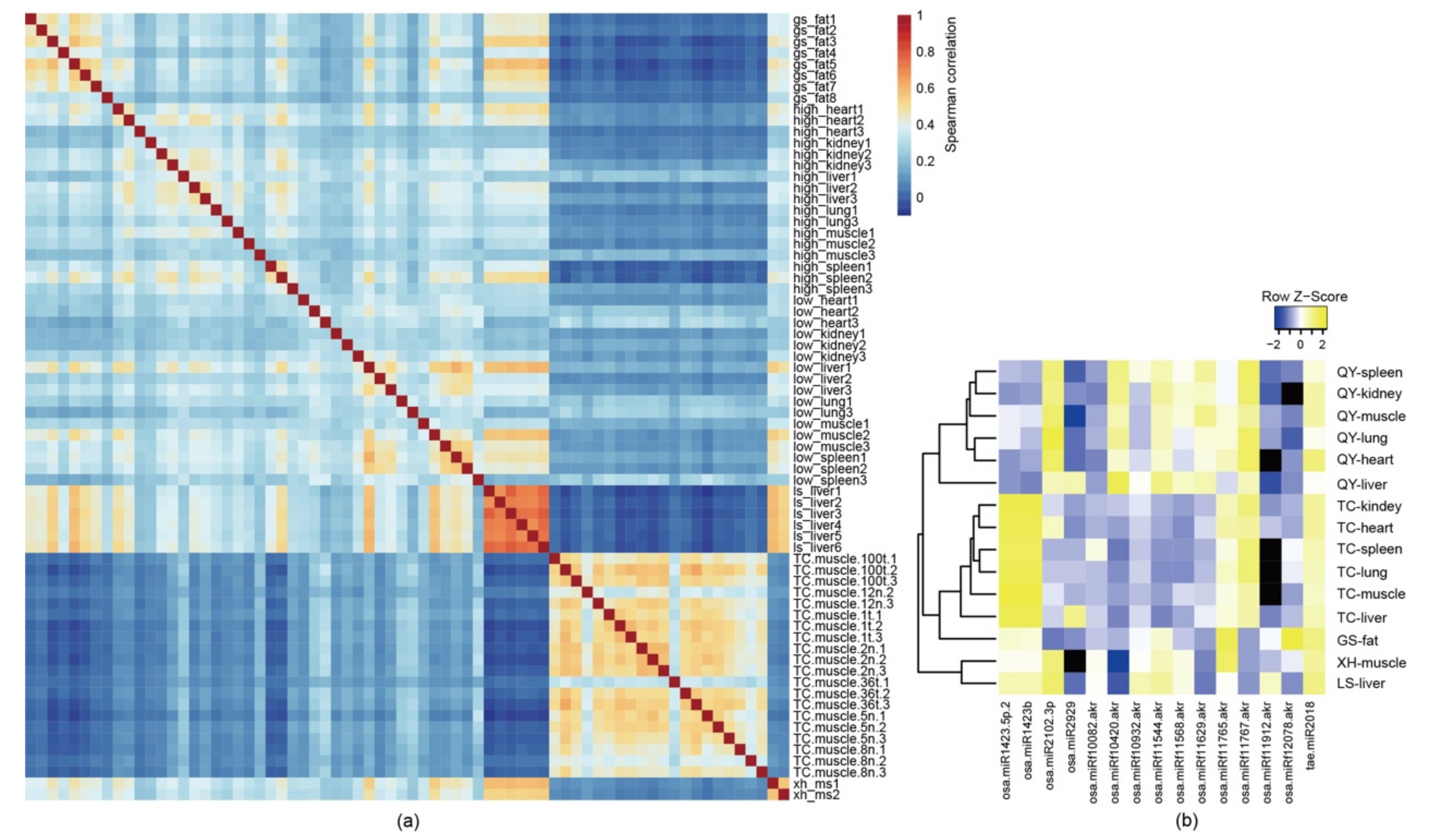
3.1. Plant miRNAs Are Detected in Multiple Tissues and Organs of Free-Range Chickens
3.2. Plant miRNAs Are Present in Chickens of Different Ages
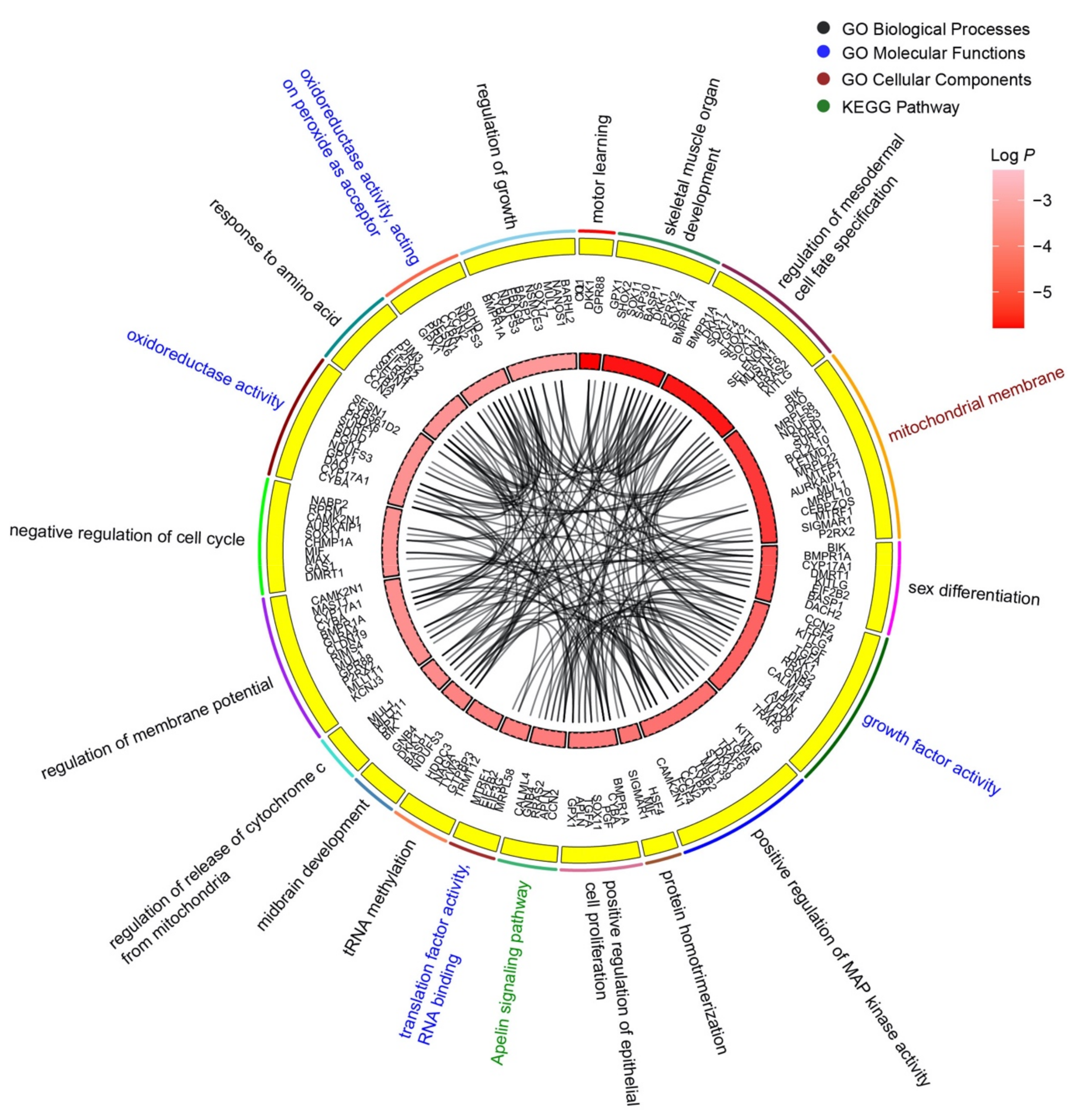
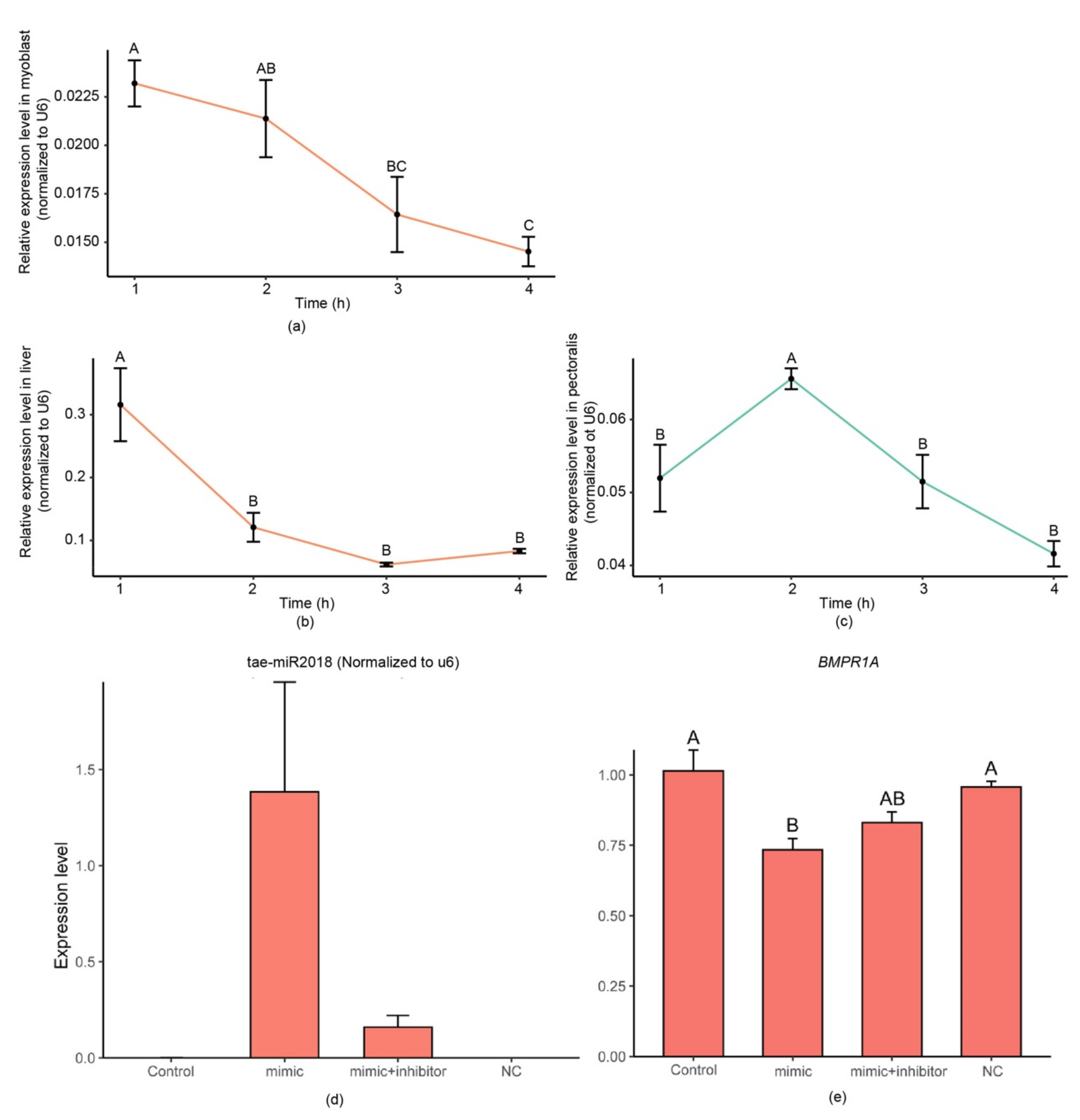
3.3. Wheat-miR2018 May Regulate the Development of Chicken Embryos
3.4. Synthetic Tae-miR2018 Can Be Absorbed by Chicken Myoblasts and by Chickens In Vivo
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of miRNA regulation. Genom. Proteom. Bioinform. 2009, 7, 147–154. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Zhang, Y.; Du, J.; Chen, L.; Zhu, L. MicroRNA-23a regulates 3T3-L1 adipocyte differentiation. Gene 2016, 575, 761–764. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.T.; Chu, K.; Oh, H.J.; Im, W.S.; Lim, J.Y.; Kim, S.K.; Park, C.K.; Jung, K.H.; Lee, S.K.; Kim, M. Let-7 microRNA inhibits the proliferation of human glioblastoma cells. J. Neuro Oncol. 2011, 102, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.L.; Yang, L.Q.; Yin, J.M.; Su, D.S.; Pan, Z.Y.; Li, P.Y.; Wang, X.D. MicroRNA-15b deteriorates hypoxia/reoxygenation-induced cardiomyocyte apoptosis by downregulating Bcl-2 and MAPK3. J. Investig. Med. 2018, 66, 39–45. [Google Scholar] [CrossRef]
- Olsen, I.; Singhrao, S.K.; Osmundsen, H. Periodontitis, pathogenesis and progression: miRNA-mediated cellular responses to Porphyromonas gingivalis. J. Oral Microbiol. 2017, 9, 1333396. [Google Scholar] [CrossRef]
- LaMonte, G.; Philip, N.; Reardon, J.; Lacsina, J.R.; Majoros, W.; Chapman, L.; Thornburg, C.D.; Telen, M.J.; Ohler, U.; Nicchitta, C.V.; et al. Translocation of sickle cell erythrocyte microRNAs into Plasmodium falciparum inhibits parasite translation and contributes to malaria resistance. Cell Host Microbe 2012, 12, 187–199. [Google Scholar] [CrossRef]
- Stern-Ginossar, N.; Elefant, N.; Zimmermann, A.; Wolf, D.G.; Saleh, N.; Biton, M.; Horwitz, E.; Prokocimer, Z.; Prichard, M.; Hahn, G.; et al. Host immune system gene targeting by a viral miRNA. Science 2007, 317, 376–381. [Google Scholar] [CrossRef]
- Wang, Z.-Z.; Ye, X.-Q.; Shi, M.; Li, F.; Wang, Z.-H.; Zhou, Y.-N.; Gu, Q.-J.; Wu, X.-T.; Yin, C.-L.; Guo, D.-H.; et al. Parasitic insect-derived miRNAs modulate host development. Nat. Commun. 2018, 9, 2205. [Google Scholar] [CrossRef]
- Liu, X.; Happel, C.; Ziegelbauer, J.M. Kaposi’s Sarcoma-Associated Herpesvirus MicroRNAs Target GADD45B To Protect Infected Cells from Cell Cycle Arrest and Apoptosis. J. Virol. 2017, 91, e02045-16. [Google Scholar] [CrossRef]
- Liang, G.F.; Zhu, Y.L.; Sun, B.; Shao, Y.H.; Jing, A.H.; Wang, J.H.; Xiao, Z.D. Assessing the survival of exogenous plant microRNA in mice. Food Sci. Nutr. 2014, 2, 380–388. [Google Scholar] [CrossRef]
- Liang, H.W.; Zhang, S.Y.; Fu, Z.; Wang, Y.B.; Wang, N.; Liu, Y.Q.; Zhao, C.H.; Wu, J.H.; Hu, Y.Q.; Zhang, J.F.; et al. Effective detection and quantification of dietetically absorbed plant microRNAs in human plasma. J. Nutr. Biochem. 2015, 26, 505–512. [Google Scholar] [CrossRef]
- Luo, Y.; Wang, P.J.; Wang, X.; Wang, Y.H.; Mu, Z.P.; Li, Q.Z.; Fu, Y.H.; Xiao, J.; Li, G.J.; Ma, Y.; et al. Detection of dietetically absorbed maize-derived microRNAs in pigs. Sci. Rep. 2017, 7, 645. [Google Scholar] [CrossRef]
- Li, J.; Zhang, Y.; Li, D.; Liu, Y.; Chu, D.; Jiang, X.; Hou, D.; Zen, K.; Zhang, C.-Y. Small non-coding RNAs transfer through mammalian placenta and directly regulate fetal gene expression. Protein Cell 2015, 6, 391–396. [Google Scholar] [CrossRef]
- Zhang, L.; Hou, D.; Chen, X.; Li, D.; Zhu, L.; Zhang, Y.; Li, J.; Bian, Z.; Liang, X.; Cai, X.; et al. Exogenous plant MIR168a specifically targets mammalian LDLRAP1: Evidence of cross-kingdom regulation by microRNA. Cell Res. 2012, 22, 107–126. [Google Scholar] [CrossRef]
- Chin, A.R.; Fong, M.Y.; Somlo, G.; Wu, J.; Swiderski, P.; Wu, X.; Wang, S.E. Cross-kingdom inhibition of breast cancer growth by plant miR159. Cell Res. 2016, 26, 217–228. [Google Scholar] [CrossRef]
- Zhang, T.; Zhao, Y.L.; Zhao, J.H.; Wang, S.; Jin, Y.; Chen, Z.Q.; Fang, Y.Y.; Hua, C.L.; Ding, S.W.; Guo, H.S. Cotton plants export microRNAs to inhibit virulence gene expression in a fungal pathogen. Nat. Plants 2016, 2, 16153. [Google Scholar] [CrossRef]
- Shahid, S.; Kim, G.; Johnson, N.R.; Wafula, E.; Wang, F.; Coruh, C.; Bernal-Galeano, V.; Phifer, T.; de Pamphilis, C.W.; Westwood, J.H.; et al. MicroRNAs from the parasitic plant Cuscuta campestris target host messenger RNAs. Nature 2018, 553, 82–85. [Google Scholar] [CrossRef]
- Zhou, Z.; Li, X.; Liu, J.; Dong, L.; Chen, Q.; Liu, J.; Kong, H.; Zhang, Q.; Qi, X.; Hou, D.; et al. Honeysuckle-encoded atypical microRNA2911 directly targets influenza A viruses. Cell Res. 2015, 25, 39–49. [Google Scholar] [CrossRef]
- Zhu, K.; Liu, M.; Fu, Z.; Zhou, Z.; Kong, Y.; Liang, H.; Lin, Z.; Luo, J.; Zheng, H.; Wan, P. Plant microRNAs in larval food regulate honeybee caste development. PLoS Genet. 2017, 13, e1006946. [Google Scholar] [CrossRef]
- Teng, Y.; Ren, Y.; Sayed, M.; Hu, X.; Lei, C.; Kumar, A.; Hutchins, E.; Mu, J.; Deng, Z.; Luo, C.; et al. Plant-Derived Exosomal MicroRNAs Shape the Gut Microbiota. Cell Host Microbe 2018, 24, 637–652.e8. [Google Scholar] [CrossRef]
- Ponte, P.I.; Rosado, C.M.; Crespo, J.P.; Crespo, D.G.; Mourao, J.L.; Chaveiro-Soares, M.A.; Bras, J.L.; Mendes, I.; Gama, L.T.; Prates, J.A.; et al. Pasture intake improves the performance and meat sensory attributes of free-range broilers. Poult. Sci. 2008, 87, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Fontes, C.M.; Ponte, P.I.; Reis, T.C.; Soares, M.C.; Gama, L.T.; Dias, F.M.; Ferreira, L.M. A family 6 carbohydrate-binding module potentiates the efficiency of a recombinant xylanase used to supplement cereal-based diets for poultry. Br. Poult. Sci. 2004, 45, 648–656. [Google Scholar] [CrossRef]
- Al-Khalaifa, H.; Al-Nasser, A.; Al-Surayee, T.; Al-Kandari, S.; Al-Enzi, N.; Al-Sharrah, T.; Ragheb, G.; Al-Qalaf, S.; Mohammed, A. Effect of dietary probiotics and prebiotics on the performance of broiler chickens. Poult. Sci. 2019, 98, 4465–4479. [Google Scholar] [CrossRef]
- Khan, S.; Moore, R.J.; Stanley, D.; Chousalkar, K.K. The Gut Microbiota of Laying Hens and Its Manipulation with Prebiotics and Probiotics To Enhance Gut Health and Food Safety. Appl. Environ. Microbiol. 2020, 86, e00600-20. [Google Scholar] [CrossRef] [PubMed]
- Rio, D.C.; Ares, M., Jr.; Hannon, G.J.; Nilsen, T.W. Purification of RNA using TRIzol (TRI reagent). Cold Spring Harb. Protoc. 2010, 2010, pdb-rot5439. [Google Scholar] [CrossRef]
- Langmead, B. Aligning short sequencing reads with Bowtie. Curr. Protoc. Bioinform. 2010, 32, 11.7.1–11.7.14. [Google Scholar] [CrossRef]
- Rehmsmeier, M.; Steffen, P.; Hochsmann, M.; Giegerich, R. Fast and effective prediction of microRNA/target duplexes. RNA 2004, 10, 1507–1517. [Google Scholar] [CrossRef]
- Helwak, A.; Kudla, G.; Dudnakova, T.; Tollervey, D. Mapping the human miRNA interactome by CLASH reveals frequent noncanonical binding. Cell 2013, 153, 654–665. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef]
- Liu, Y.-C.; Chen, W.L.; Kung, W.-H.; Huang, H.-D. Plant miRNAs found in human circulating system provide evidences of cross kingdom RNAi. BMC Genom. 2017, 18, 112. [Google Scholar] [CrossRef]
- Zhao, Y.; Cong, L.; Lukiw, W.J. Plant and Animal microRNAs (miRNAs) and Their Potential for Inter-kingdom Communication. Cell. Mol. Neurobiol. 2017, 38, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H.; Chupeau, Y. Ingested plant miRNAs regulate gene expression in animals. Cell Res. 2012, 22, 3–5. [Google Scholar] [CrossRef]
- Lukasik, A.; Zielenkiewicz, P. In Silico Identification of Plant miRNAs in Mammalian Breast Milk Exosomes—A Small Step Forward? PLoS ONE 2014, 9, e99963. [Google Scholar] [CrossRef]
- Zhang, H.; Li, Y.P.; Liu, Y.N.; Liu, H.M.; Wang, H.Y.; Jin, W.; Zhang, Y.M.; Zhang, C.; Xu, D. Role of plant MicroRNA in cross-species regulatory networks of humans. BMC Syst. Biol. 2016, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhang, F.; Dong, L.; Wu, H.; Xu, J.; Li, H.; Wang, J.; Zhou, Z.; Liu, C.; Wang, Y.; et al. SIDT1-dependent absorption in the stomach mediates host uptake of dietary and orally administered microRNAs. Cell Res. 2020, 31, 247–258. [Google Scholar] [CrossRef]
- Mabelebele, M.; Norris, D.; Brown, D.; Ginindza, M.M.; Ngambi, J.W. Breed and Sex Differences in the Gross Anatomy, Digesta pH and Histomorphology of the Gastrointestinal Tract of Gallus Gallus Domesticus. Braz. J. Poult. Sci. 2017, 19, 339–346. [Google Scholar] [CrossRef]
- O’Neill, M.J.; Bourre, L.; Melgar, S.; O’Driscoll, C.M. Intestinal delivery of non-viral gene therapeutics: Physiological barriers and preclinical models. Drug Discov. Today 2011, 16, 203–218. [Google Scholar] [CrossRef]
- Stadelman, W.J. Egg Science and Technology, 4th ed.; CRC Press: Boca Raton, FL, USA, 1995. [Google Scholar]
- Ayerza, R.; Coates, W. Dietary levels of chia: Influence on yolk cholesterol, lipid content and fatty acid composition for two strains of hens. Poult. Sci. 2000, 79, 724–739. [Google Scholar] [CrossRef]
- Saadoun, A.; Leclercq, B. Comparison of in vivo fatty acid synthesis of the genetically lean and fat chickens. Comp. Biochem. Physiol. Part B Comp. Biochem. 1983, 75, 641–644. [Google Scholar] [CrossRef]
- Klasing, K.C. Comparative avian nutrition. In Quarterly Review of Biology; Cab International: Wallingford, UK, 1998. [Google Scholar]
- Chen, D.; Zhao, M.; Mundy, G.R. Bone morphogenetic proteins. Growth Factors 2004, 22, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yuan, Q.; Niu, M.; Hou, J.; He, Z. BMP4 promotes mouse iPS cell differentiation to male germ cells via Smad1/5, Gata4, Id1 and Id2. Reproduction 2017, 153, 211. [Google Scholar] [CrossRef]
- Muñoz-Sanjuán, I.; Brivanlou, A.H. Neural induction, the default model and embryonic stem cells. Nat. Rev. Neurosci. 2002, 3, 271–280. [Google Scholar] [CrossRef] [PubMed]
- Marti, E. Expression of chick BMP-1/Tolloid during patterning of the neural tube and somites. Mech. Dev. 2000, 91, 415–419. [Google Scholar] [CrossRef]
- Song, X.; Liu, S.; Qu, X.; Hu, Y.; Zhang, X.; Wang, T.; Wei, F. BMP2 and VEGF promote angiogenesis but retard terminal differentiation of osteoblasts in bone regeneration by up-regulating Id1. Acta Biochim. Biophys. Sin. (Shanghai) 2011, 43, 796–804. [Google Scholar] [CrossRef] [PubMed]
- Langenfeld, E.M.; Langenfeld, J.; Langenfeld, E.M.; Langenfeld, J. Bone morphogenetic protein-2 stimulates angiogenesis in developing tumors. Mol Cancer Res 2: 141-149. Mol. Cancer Res. 2004, 2, 141–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Ji, X.; Harris, M.A.; Feng, J.Q.; Karsenty, G.; Celeste, A.J.; Rosen, V.; Mundy, G.R.; Harris, S.E. Differential roles for bone morphogenetic protein (BMP) receptor type IB and IA in differentiation and specification of mesenchymal precursor cells to osteoblast and adipocyte lineages. J. Cell Biol. 1998, 142, 295–305. [Google Scholar] [CrossRef]
- Estrada, K.D.; Lyons, K.M. BMP Signaling in Skeletogenesis; Springer: London, UK, 2010. [Google Scholar]
- Yuhki, M. BMPR1A signaling is necessary for hair follicle cycling and hair shaft differentiation in mice. Development 2004, 131, 1825. [Google Scholar] [CrossRef]
- Cole, L.K.; Le Roux, I.; Nunes, F.; Laufer, E.; Lewis, J.; Wu, D.K. Sensory organ generation in the chicken inner ear: Contributions of bone morphogenetic protein 4, serrate1, and lunatic fringe. J. Comp. Neurol. 2000, 424, 509–520. [Google Scholar] [CrossRef]
- Huang, J.; Liu, Y.; Filas, B.; Gunhaga, L.; Beebe, D.C. Negative and positive auto-regulation of BMP expression in early eye development. Dev. Biol. 2015, 407, 256–264. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chicken Breed | Tissues | Number | NCBI Access Number | Age |
|---|---|---|---|---|
| Tibetan chicken (TC) | Heart | 3 | PRJNA511987 | >200 days |
| Liver | 3 | >200 days | ||
| Spleen | 3 | >200 days | ||
| Lung | 2 | >200 days | ||
| Kidney | 3 | >200 days | ||
| Leg muscle (Tibialis anterior) | 3 | >200 days | ||
| Qingyuan chicken (QY) | Heart | 3 | PRJNA511987 | >200 days |
| Liver | 3 | >200 days | ||
| Spleen | 3 | >200 days | ||
| Lung | 2 | >200 days | ||
| Kidney | 3 | >200 days | ||
| Leg muscle (Tibialis anterior) | 3 | >200 days | ||
| Xinghua chicken (XH) | Pectoralis muscle | 2 | PRJNA266323 | 7 Weeks |
| Gushi chicken (GS) | Fat (Abdominal adipose tissue) | 4 | PRJNA528858 | 6, 14, 22 and 30 weeks |
| Pectoralis muscle | 4 | PRJNA516961 | 6, 14, 22 and 30 weeks | |
| Lushi chicken (LS) | Liver | 6 | PRJNA299589 | 20 and 30 weeks |
| Tibetan chicken (TC) 1 | Pectoralis muscle | 21 | PRJNA699998 | Embryo stage (5, 7, 9, 12, 15, 18 and 20 days) |
| 20 | PRJNA699998 | After hatching (1, 36, 100 days and 2, 5, 8 and 12 years) | ||
| Broiler (ROSS308) | skeletal muscle | 24 | PRJNA516545 | Embryo stage (10, 13, 16 and 19 days) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, H.; Zhang, P.; Li, D.; Chen, B.; Li, J.; Wang, T. The Expression Patterns of Exogenous Plant miRNAs in Chickens. Genes 2023, 14, 760. https://doi.org/10.3390/genes14030760
Li H, Zhang P, Li D, Chen B, Li J, Wang T. The Expression Patterns of Exogenous Plant miRNAs in Chickens. Genes. 2023; 14(3):760. https://doi.org/10.3390/genes14030760
Chicago/Turabian StyleLi, Hao, Pu Zhang, Diyan Li, Binlong Chen, Jing Li, and Tao Wang. 2023. "The Expression Patterns of Exogenous Plant miRNAs in Chickens" Genes 14, no. 3: 760. https://doi.org/10.3390/genes14030760
APA StyleLi, H., Zhang, P., Li, D., Chen, B., Li, J., & Wang, T. (2023). The Expression Patterns of Exogenous Plant miRNAs in Chickens. Genes, 14(3), 760. https://doi.org/10.3390/genes14030760