


Reproductive and Agronomic Characterization of Novel Apomictic Hybrids of Paspalum (Poaceae)

 , ,
, ,  and
and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material and Agronomic Evaluation
2.1.1. P. notatum
2.1.2. P. simplex
2.2. Evaluation of Apomixis Expressivity in Seeds of P. simplex Hybrids
2.3. Genotypic Homogeneity within the Progeny
3. Results
3.1. Agronomic Evaluations
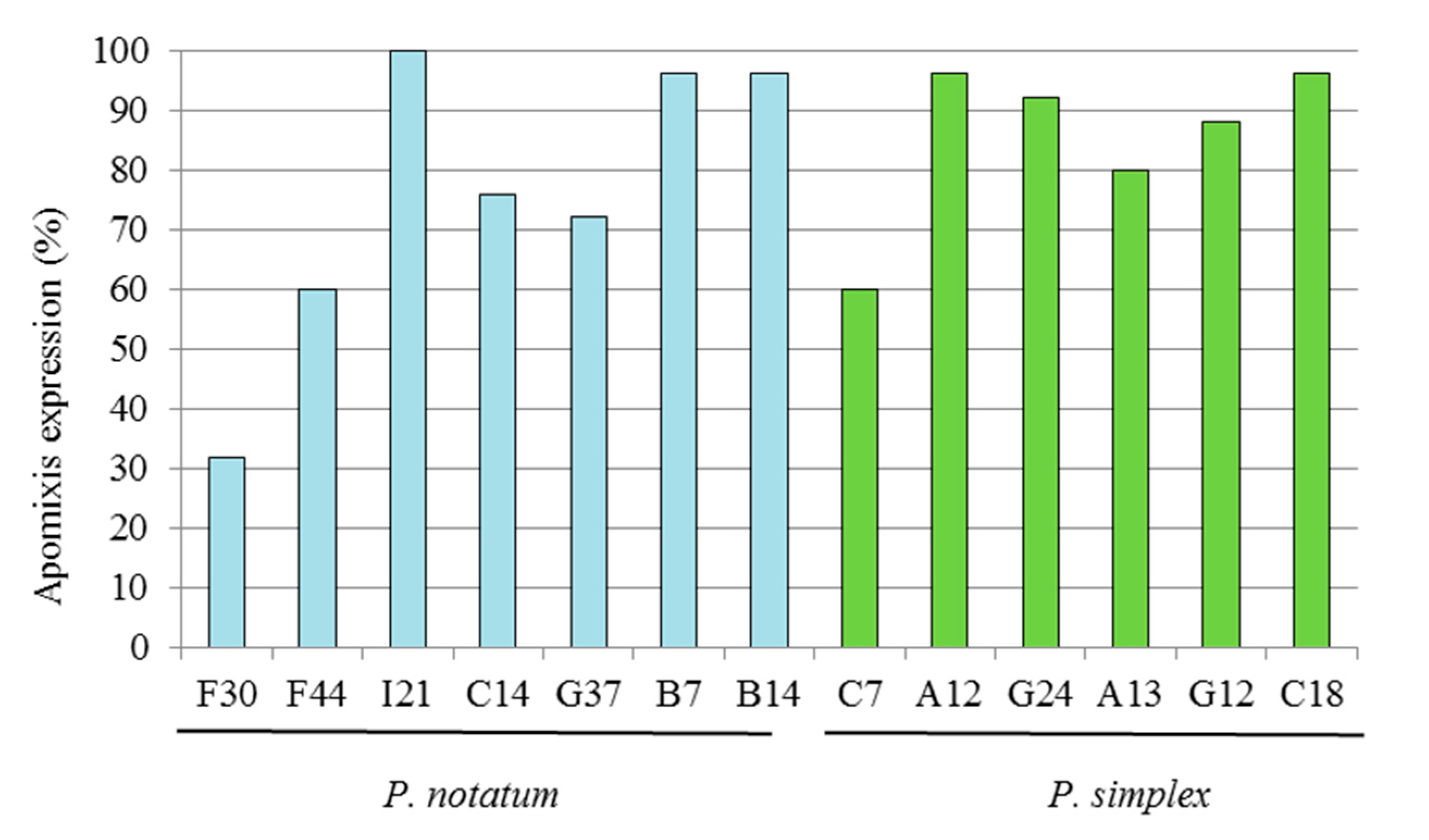
3.1.1. P. notatum
3.1.2. P. simplex
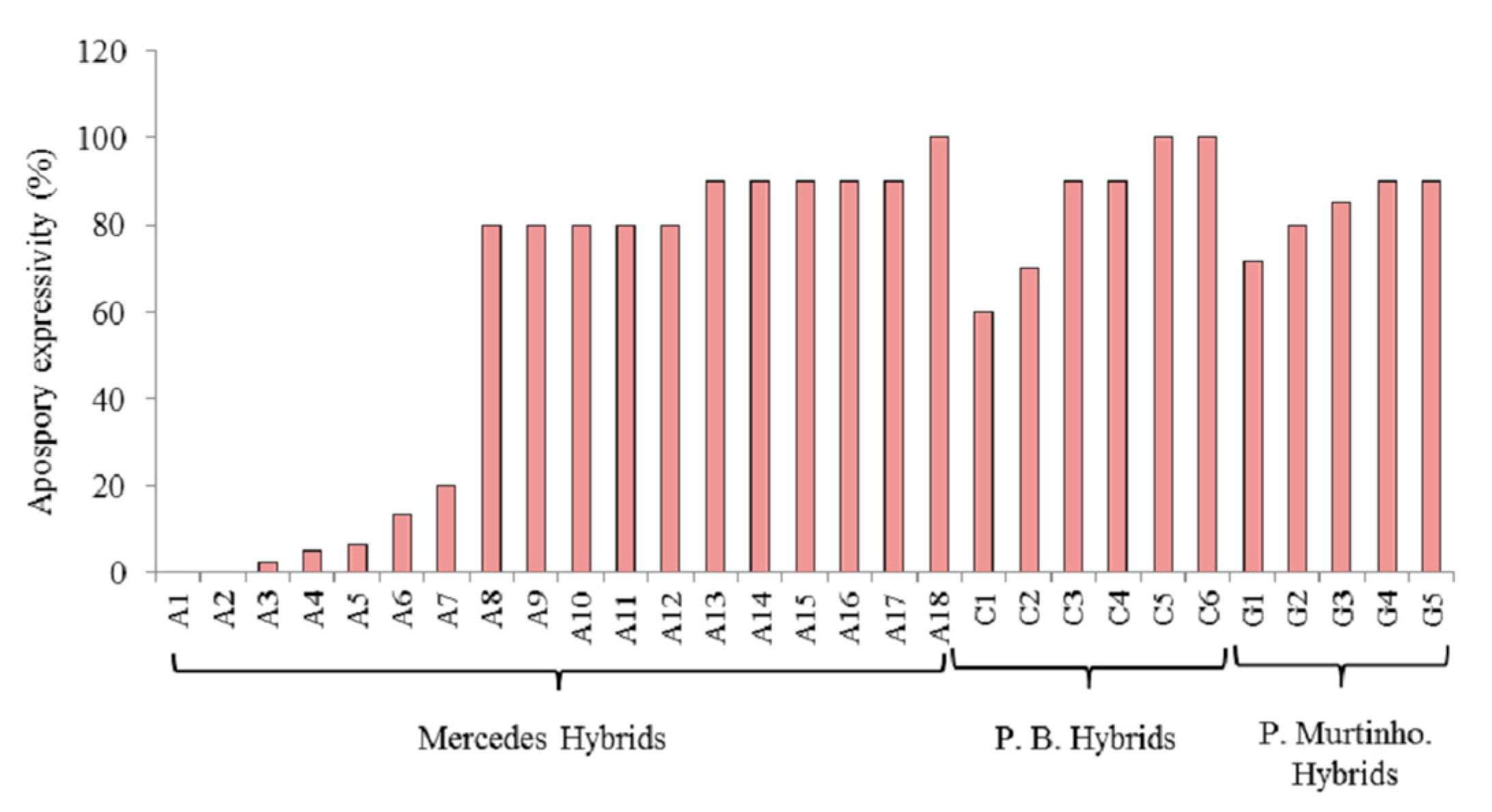
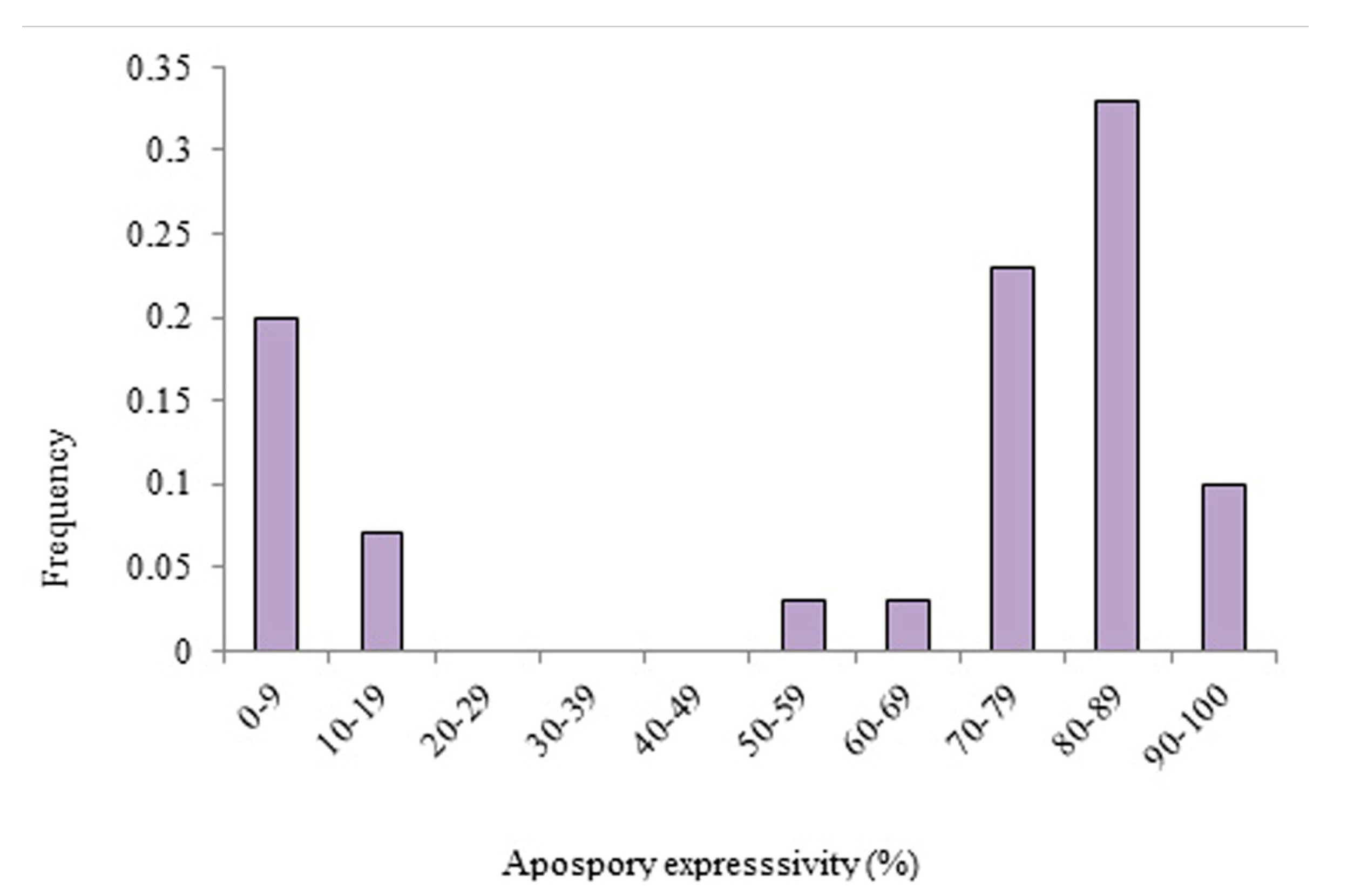
3.2. Apomixis Expressivity in P. simplex
3.3. Genotypic Homogeneity in Progenies of P. notatum and P. simplex Hybrids
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Skerman, P.J.; Riveros, F. Gramíneas Tropicales. In Colección FAO: Protección Vegetal, 23rd ed.; Organización de las Naciones Unidas para la Agricultura y la Alimentación: Rome, Italy, 1992; p. 849. [Google Scholar]
- Jaurena, M.; Durante, M.; Devincenzi, T.; Savian, J.V.; Bendersky, D.; Moojen, F.G.; Pereira, M.; Soca, P.; Quadros, F.L.F.; Pizzio, R.; et al. Native grasslands at the core: A new paradigm of intensification for the campos of Southern South America to increase economic and environmental sustainability. Front. Sustain. Food Syst. 2021, 5, 547834. [Google Scholar] [CrossRef]
- Burkart, A. Evolution of grasses and grasslands in South America. Taxon 1975, 24, 53–66. [Google Scholar] [CrossRef]
- Zuloaga, F.O.; Morrone, O. Revisión de las Especies de Paspalum para América del Sur Austral (Argentina, Bolivia, sur de Brasil, Chile, Paraguay y Uruguay); Misouri Botanical Garden Press: St. Louis, MO, USA, 2005; p. 298. [Google Scholar]
- Urbani, M.H.; Acuña, C.A.; Doval, D.W.; Sartor, M.E.; Galdeano, F.; Blount, A.R.; Quesenberry, K.H.; Mackowiak, C.L.; Quarin, C.L. Registration of ‘Boyero UNNE’ Bahiagrass. J. Plant Regist. 2017, 11, 26–32. [Google Scholar] [CrossRef]
- Acuña, C.A.; Martínez, E.J.; Zilli, A.L.; Brugnoli, E.A.; Espinoza, F.; Marcón, F.; Urbani, M.H.; Quarin, C.L. Reproductive systems in Paspalum: Relevance for germplasm collection and conservation, breeding techniques, and adoption of released cultivars. Front. Plant Sci. 2019, 10, 1377. [Google Scholar] [CrossRef] [PubMed]
- Blount, A.R.; Acuña, C.A. Bahiagrass. In Genetic Resources, Chromosome Engineering, and Crop Improvement Series, in Forage Crops; Singh, R.J., Ed.; CRC Press: Boca Raton, FL, USA, 2009; Volume 5, pp. 81–101. [Google Scholar] [CrossRef]
- Brugnoli, A.E.; Urbani, M.H.; Quarin, C.L.; Zilli, A.L.; Martínez, E.J.; Acuña, C.A. Diversity in apomictic populations of Paspalum simplex Morong. Crop Sci. 2014, 53, 1509–1516. [Google Scholar] [CrossRef]
- Morrone, O.; Denham, S.S.; Aliscioni, S.S.; Zuloaga, F.O. Revisión de las especies de Paspalum (Panicoideae: Paniceae), subgénero Anachyris. Candollea 2000, 55, 105–135. [Google Scholar]
- Daurelio, L.D.; Espinoza, F.; Quarin, C.L.; Pessino, S.C. Genetic diversity in sexual diploid and apomictic tetraploid populations of Paspalum notatum situated in sympatry or allopatry. Plant Syst. Evol. 2004, 244, 189–199. [Google Scholar] [CrossRef]
- Miles, J.W. Apomixis for cultivar development in tropical forage grasses. Crop Sci. 2007, 47, 238–249. [Google Scholar] [CrossRef]
- Jank, L.; Barrios, S.C.; do Valle, C.; Simeão, R.A.; Alves, G.F. The value of improved pastures to Brazilian beef production. Crop Pasture Sci. 2014, 65, 1132–1137. [Google Scholar] [CrossRef]
- Vogel, K.P.; Burson, B.L. Breeding and genetics. In Warm-Season (C4) Grasses; Moser, L., Burson, B., Sollenberger, L., Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 51–94. [Google Scholar]
- Muir, J.P.; Jank, L. Guineagrass. In Warm-Season (C4) Grasses; Moser, L., Burson, B., Sollenberger, L., Eds.; American Society of Agronomy: Madison, WI, USA, 2004; pp. 589–622. [Google Scholar]
- Argel, P.; Miles, J.; Guiot, J.; Lescano, C. Cultivar Mulato II (Brachiaria híbrido CIAT 36087): Gramínea de alta Calidad y Producción Forrajera, Resistente a Salivazo y Adaptada a Suelos Tropicales Ácidos bien Drenados; Centro Internacional de Agricultura Tropical—CIAT: Cali, Colombia, 2007; p. 21. [Google Scholar]
- Argel, P.; Miles, J.; Guiot, J.; Lescano, C. Cultivar Mulato (Brachiaria Híbrido CIAT 36061): Gramínea de Alta Producción y Calidad Forrajera para los Trópicos; Centro Internacional de Agricultura Tropical—CIAT: Cali, Colombia, 2006; p. 22. [Google Scholar]
- Acuña, C.A.; Blount, A.R.; Quesenberry, K.H.; Kenworthy, K.E.; Hanna, W.W. Tetraploid bahiagrass hybrids: Breeding technique, genetic variability and proportion of heterotic hybrids. Euphytica 2011, 179, 227–235. [Google Scholar] [CrossRef]
- Brugnoli, E.A.; Martínez, E.J.; Ferrari Usandizaga, S.C.; Zilli, A.L.; Urbani, M.H.; Acuña, C.A. Breeding tetraploid Paspalum simplex: Hybridization, early identification of apomicts, and impact of apomixis on hybrid performance. Crop Sci. 2019, 59, 1617–1624. [Google Scholar] [CrossRef]
- Zilli, A.L.; Brugnoli, E.A.; Marcón, F.; Billa, M.B.; Rios, E.F.; Martínez, E.J.; Acuña, C.A. Heterosis and expressivity of apospory in tetraploid bahiagrass hybrids. Crop Sci. 2015, 55, 1189–1201. [Google Scholar] [CrossRef]
- Novo, P.E.; Acuña, C.A.; Quarin, C.L.; Urbani, M.H.; Marcón, F.; Espinoza, F. Hybridization and heterosis in the Plicatula group of Paspalum. Euphytica 2017, 213, 198. [Google Scholar] [CrossRef]
- Motta, E.A.; Dall’AgnolI, M.; Nascimento, F.L.; Pereira, E.A.; Machado, J.M. Forage performance of Paspalum hybrids from an interspecific cross. Cienc. Rural. 2016, 46, 1025–1031. [Google Scholar] [CrossRef]
- Weiler, R.L.; Dall’Agnol, M.; Simioni, C.; Krycki, K.C.; Pereira, E.A.; Machado, J.M.; Minski da Motta, E.A. Intraspecific tetraploid hybrids of Paspalum notatum: Agronomic evaluation of segregating progeny. Sci. Agric. 2018, 75, 36–42. [Google Scholar] [CrossRef]
- Burton, G.W.; Forbes, I., Jr. The genetics and manipulation of obligate apomixis in common bahiagrass (Paspalum notatum Flüggé). In Proceedings of the 8th International Grasslands Congress, Oxford, UK, 11–21 July 1960; pp. 66–71. [Google Scholar]
- Quarin, C.L.; Espinoza, F.; Martínez, E.J.; Pessino, S.C.; Bovo, O.A. A rise of ploidy level induces the expression of apomixis in Paspalum notatum. Sex Plant Reprod. 2001, 13, 243–249. [Google Scholar] [CrossRef]
- Cáceres, M.E.; Pupilli, F.; Quarin, C.L.; Arcioni, S. Feulgen-DNA densitometry of embryo sacs permits discrimination between sexual and apomictic plants in Paspalum simplex. Euphytica 1999, 110, 161–167. [Google Scholar] [CrossRef]
- Quesenberry, K.H.; Dampier, J.M.; Lee, Y.Y.; Smith, R.L.; Acuña, C.A. Doubling the chromosome number of bahiagrass via tissue culture. Euphytica 2010, 175, 43–50. [Google Scholar] [CrossRef]
- Caponio, I.; Quarin, C.L. El sistema genético de Paspalum simplex y de un híbrido interespecífico con P. dilatatum. Kurtziana 1987, 19, 35–45. [Google Scholar]
- Matzk, F.; Meister, A.; Schubert, I. An efficient screen for the reproductive pathways using mature seeds of monocots and dicots. Plant J. 2000, 21, 97–108. [Google Scholar] [CrossRef]
- Hojsgaard, D.; Schegg, E.; Valls, J.F.M.; Martínez, E.J.; Quarin, C.L. Sexuality, apomixis, ploidy levels, and genomic relationships among four Paspalum species of the subgenus Anachyris (Poaceae). Flora 2008, 203, 535–547. [Google Scholar] [CrossRef]
- Hojsgaard, D.H.; Martínez, E.J.; Acuña, C.A.; Quarin, C.L.; Pupilli, F. A molecular map of the apomixis-control locus in Paspalum procurrens and its comparative analysis with other species of Paspalum. Theor. Appl. Genet. 2011, 123, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Versión 2019. Available online: https://www.infostat.com.ar/ (accessed on 4 March 2021).
- Galdeano, F.; Urbani, M.H.; Sartor, M.E.; Honfi, A.I.; Espinoza, F.; Quarin, C.L. Relative DNA content in diploid, polyploid, and multiploid species of Paspalum (Poaceae) with relation to reproductive mode and taxonomy. J. Plant Res. 2016, 129, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Meirmans, P.G.; Van Tienderen, P. GenoType and GenoDive: Two programs for the analysis of genetic diversity of asexual organisms. Mol. Ecol. Notes 2004, 4, 792–794. [Google Scholar] [CrossRef]
- Marcón, F.; Brugnoli, E.A.; Rodriguez Nunes, J.A.; Gutierrez Storti, V.I.; Martínez, E.J.; Acuña, C.A. Evaluating general combining ability for agromorphological raits in tetraploid bahiagrass. Euphytica 2021, 217, 208. [Google Scholar] [CrossRef]
- Marcón, F.; Martínez, E.J.; Rodríguez, G.R.; Zilli, A.L.; Brugnoli, E.A.; Acuña, C.A. Genetic distance and the relationship with heterosis and reproductive behavior in tetraploid bahiagrass hybrids. Genetic distance and the relationship with heterosis and reproductive behavior in tetraploid bahiagrass hybrids. Mol. Breed. 2019, 39, 89. [Google Scholar] [CrossRef]
- Savidan, Y. Apomixis: Genetics and breeding. Appl. Genet. 2000, 57, 153–156. [Google Scholar]
- Rebozzio, R.N.; Sartor, M.E.; Quarin, C.L.; Espinoza, F. Residual sexuality and its seasonal variation in natural apomictic Paspalum notatum accessions. Biol. Plant. 2011, 55, 391–395. [Google Scholar] [CrossRef]
- Soliman, M.; Espinoza, F.; Ortiz, J.P.; Delgado, L. Heterochronic reproductive developmental processes between diploid and tetraploid cytotypes of Paspalum rufum. Ann. Bot. 2018, 20, 1–15. [Google Scholar] [CrossRef]
- Brugnoli, A.E.; Urbani, M.H.; Quarin, C.L.; Zilli Martínez, E.J.; Acuña, C.A. Diversity in diploid, tetraploid, and mixed diploid–tetraploid populations of Paspalum simplex. Crop Sci. 2013, 53, 1509–1516. [Google Scholar] [CrossRef]
- Hojsgaard, D.H.; Martínez, E.J.; Quarin, C.L. Competition between meiotic and apomictic pathways during ovule and seed development results in clonality. New Phytol. 2012, 197, 336–347. [Google Scholar] [CrossRef] [PubMed]
- Reutemann, A.V.; Honfi, A.I.; Karunarathne, P.; Eckers, F.; Hojsgaard, D.H.; Martínez, E.J. Variation of residual sexuality rates along reproductive development in apomictic tetraploids of Paspalum. Plants 2022, 11, 1639. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Species | Female Sexual Parent | Male Apomictic Parents | Apomictic Hybrid Family |
|---|---|---|---|
| P. notatum | SWSB (sexual white stigma bahiagrass, derived from hybrids originally generated by G.W. Burton by crossing an induced tetraploid plant with an apomictic natural tetraploid bahiagrass with white stigmata). | Q4064 (ecotype from Saladas, Corrientes, Argentina) | B [18] |
| Q4205 (obtained by self-pollination of SWSB) | Q3776 (ecotype Villa Tunari, Chapare region, Bolivia) | G [18] | |
| Q4205 | Q4064 | I [19] | |
| P. simplex | C1-2 (self-pollination of a colchicine-induced autotetraploid plant obtained from the Istituto di Ricerche sul Miglioramento Genetico delle Piante Foraggere del Consiglio Nazionale delle Ricerche, Perugia, Italy) [25,29] | Mercedes (ecotype from 29°10′ S, 58°04′ W) | A [18] |
| C1-2 | Piedras Blancas (ecotype from 31°11′ S, 59°57′ W) | C [18] | |
| C1B2 (self-pollination of a colchicine-induced autotetraploid plant obtained from the Istituto di Ricerche sul Miglioramento Genetico delle Piante Foraggere del Consiglio Nazionale delle Ricerche, Perugia, Italy [25,30] | Porto Murtinho (ecotype from 21°42′ S, 57°51′ W) | G [18] |
| Forage Yield First Year | Forage Yield Second Year | Forage Yield Third Year | Seed Yield | Cold Tolerance * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Febr | March | May | Sep | Dec | Accu | Febr | March | June | Nov | Accu | Febr | June | Sep | Febr | Accu | ||||
| Controls | cv. Argentine | 166 | 165 | 74 | 31 | 264 | 700 | 205 | 271 | 299 | 257 | 1032 | 202 | 252 | 16 | 198 | 668 | 200 | 3 |
| cv. Boyero UNNE | 274 | 215 | 148 | 86 | 268 | 991 | 282 | 261 | 304 | 292 | 1139 | 272 | 301 | 64 | 276 | 913 | 100 | 2.3 | |
| cv. Callide | 378 | 379 | 270 | 114 | 503 | 1644 | 59 | 350 | 304 | 137 | 850 | 225 | 197 | nd | 285 | 707 | nd | 1 | |
| P. notatutm hybrids | F44 | 260 | 257 | 146 | 86 | 219 | 968 | 269 | 259 | 223 | 240 | 991 | 259 | 280 | 41 | 244 | 824 | 80 | 1 |
| I21 | 223 | 226 | 119 | 56 | 217 | 841 | 164 | 230 | 235 | 258 | 887 | 230 | 301 | 43 | 271 | 845 | 90 | 2.3 | |
| I7 | 286 | 274 | 128 | 39 | 377 | 1104 | 273 | 262 | 232 | 204 | 971 | 269 | 330 | 9 | 273 | 881 | 240 | 3.3 | |
| C11 | 266 | 268 | 152 | 97 | 369 | 1152 | 120 | 264 | 254 | 361 | 999 | 335 | 360 | 43 | 425 | 1163 | 50 | 2 | |
| C14 | 276 | 276 | 184 | 93 | 343 | 1172 | 367 | 342 | 272 | 294 | 1275 | 384 | 360 | 70 | 280 | 1094 | 70 | 2 | |
| C20 | 215 | 211 | 358 | 73 | 224 | 1081 | 169 | 253 | 254 | 162 | 838 | 251 | 281 | 34 | 448 | 1014 | 40 | 2.3 | |
| C21 | 212 | 219 | 94 | 67 | 173 | 765 | 191 | 169 | 208 | 202 | 770 | 354 | 375 | 52 | 271 | 1052 | 50 | 1.3 | |
| C3 | 270 | 279 | 113 | 75 | 248 | 985 | 279 | 270 | 248 | 328 | 1125 | 267 | 347 | 53 | 358 | 1025 | 120 | 3.3 | |
| G37 | 252 | 255 | 148 | 63 | 359 | 1077 | 281 | 369 | 314 | 363 | 1327 | 321 | 352 | 31 | 283 | 987 | 50 | 1.7 | |
| G6 | 220 | 214 | 69 | 39 | 245 | 787 | 212 | 235 | 199 | 220 | 866 | 163 | 238 | 48 | 261 | 710 | 50 | 3 | |
| B12 | 203 | 171 | 96 | 71 | 194 | 735 | 169 | 186 | 209 | 332 | 896 | 166 | 231 | 57 | 262 | 716 | 20 | 2.3 | |
| B14 | 149 | 171 | 109 | 49 | 305 | 783 | 212 | 237 | 259 | 281 | 989 | 199 | 280 | 38 | 368 | 885 | 60 | 3 | |
| B7 | 314 | 334 | 243 | 119 | 356 | 1366 | 330 | 313 | 323 | 367 | 1333 | 264 | 339 | 81 | 309 | 993 | 20 | 1 | |
| UF13 | 247 | 220 | 182 | 99 | 481 | 1229 | 322 | 279 | 246 | 380 | 1227 | 377 | 283 | 78 | 223 | 961 | 150 | 4.7 | |
| UF3 | 212 | 215 | 190 | 107 | 355 | 1079 | 276 | 270 | 198 | 287 | 1031 | 337 | 286 | 52 | 244 | 919 | 70 | 3.3 | |
| UF67 | 242 | 228 | 130 | 46 | 357 | 1003 | 226 | 271 | 244 | 176 | 917 | 268 | 197 | 24 | 195 | 684 | 160 | 2.3 | |
| UF93 | 225 | 221 | 130 | 47 | 416 | 1039 | 302 | 282 | 267 | 340 | 1191 | 294 | 254 | 40 | 230 | 818 | 70 | 3.3 | |
| MSD | 182 | 182 | 375 | 144 | 418 | 1029 | 324 | 237 | 190 | 258 | 844 | 389 | 323 | 58 | 222 | 745 | 110 | 2.9 | |
| ID | Forage Yield | Winter Regrowth * | |
|---|---|---|---|
| Control | M. maximus cv. Gatton | 166.0 | 3.4 |
| P. simplex ecotype | Reconquista | 244.5 | 3.7 |
| Villa Ana | 217.7 | 4.7 | |
| Santa Ana | 215.5 | 1 | |
| P. simplex hybrids | A26 | 250.5 | 4.3 |
| C 24 | 234.5 | 3.7 | |
| A 27 | 232.0 | 4.0 | |
| A 19 | 177.6 | 1.3 | |
| A 14 | 168.4 | 3.7 | |
| A17 | 103.1 | 2.3 | |
| A12 | 91.5 | 1.3 | |
| MSD | 130 | 1.7 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brugnoli, E.A.; Zilli, A.L.; Marcón, F.; Caballero, E.; Martínez, E.J.; Acuña, C.A. Reproductive and Agronomic Characterization of Novel Apomictic Hybrids of Paspalum (Poaceae). Genes 2023, 14, 631. https://doi.org/10.3390/genes14030631
Brugnoli EA, Zilli AL, Marcón F, Caballero E, Martínez EJ, Acuña CA. Reproductive and Agronomic Characterization of Novel Apomictic Hybrids of Paspalum (Poaceae). Genes. 2023; 14(3):631. https://doi.org/10.3390/genes14030631
Chicago/Turabian StyleBrugnoli, Elsa Andrea, Alex Leonel Zilli, Florencia Marcón, Enzo Caballero, Eric Javier Martínez, and Carlos Alberto Acuña. 2023. "Reproductive and Agronomic Characterization of Novel Apomictic Hybrids of Paspalum (Poaceae)" Genes 14, no. 3: 631. https://doi.org/10.3390/genes14030631
APA StyleBrugnoli, E. A., Zilli, A. L., Marcón, F., Caballero, E., Martínez, E. J., & Acuña, C. A. (2023). Reproductive and Agronomic Characterization of Novel Apomictic Hybrids of Paspalum (Poaceae). Genes, 14(3), 631. https://doi.org/10.3390/genes14030631