
Whole Genome Resequencing Identifies Single-Nucleotide Polymorphism Markers of Growth and Reproduction Traits in Zhedong and Zi Crossbred Geese
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
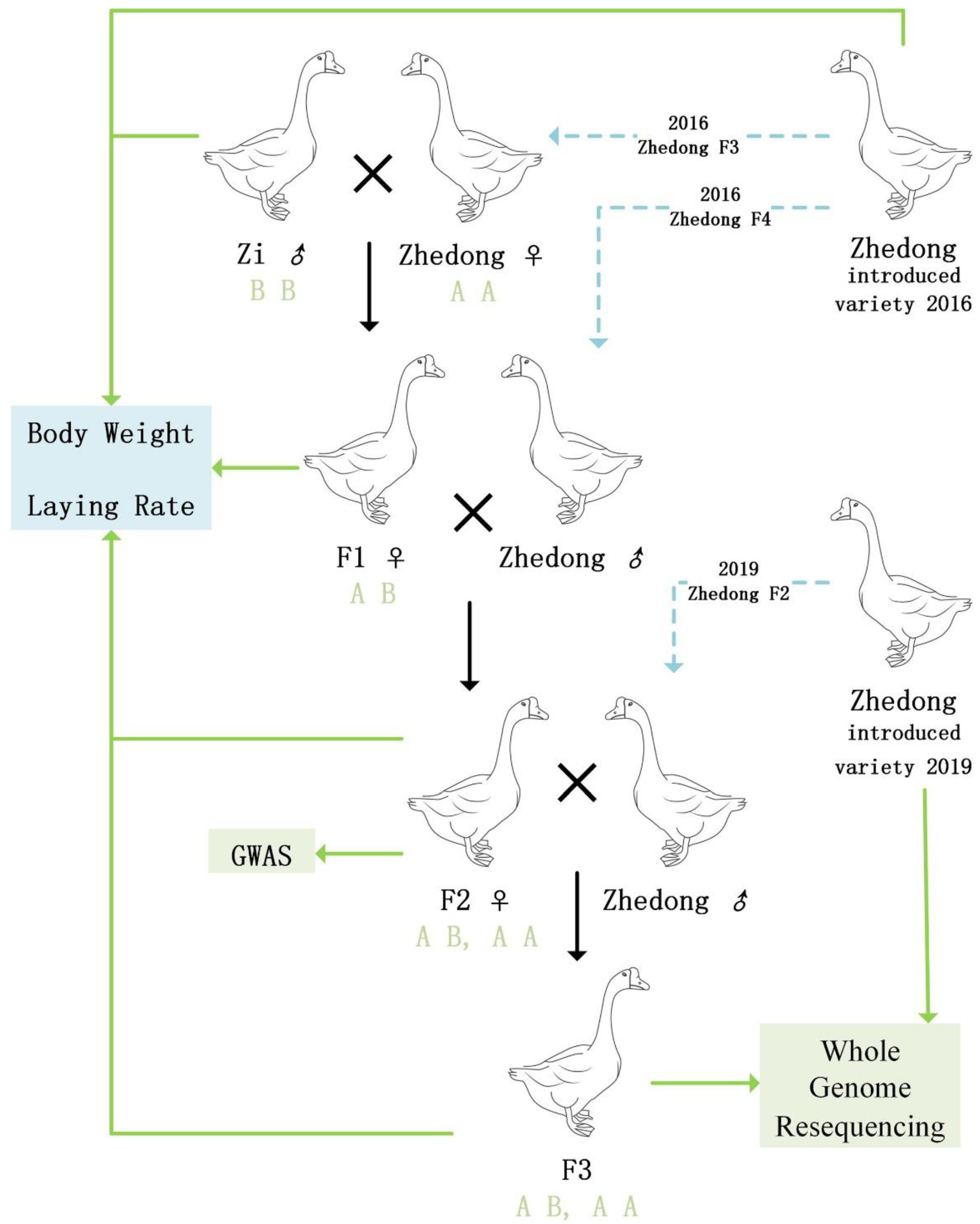
2.2. Animals
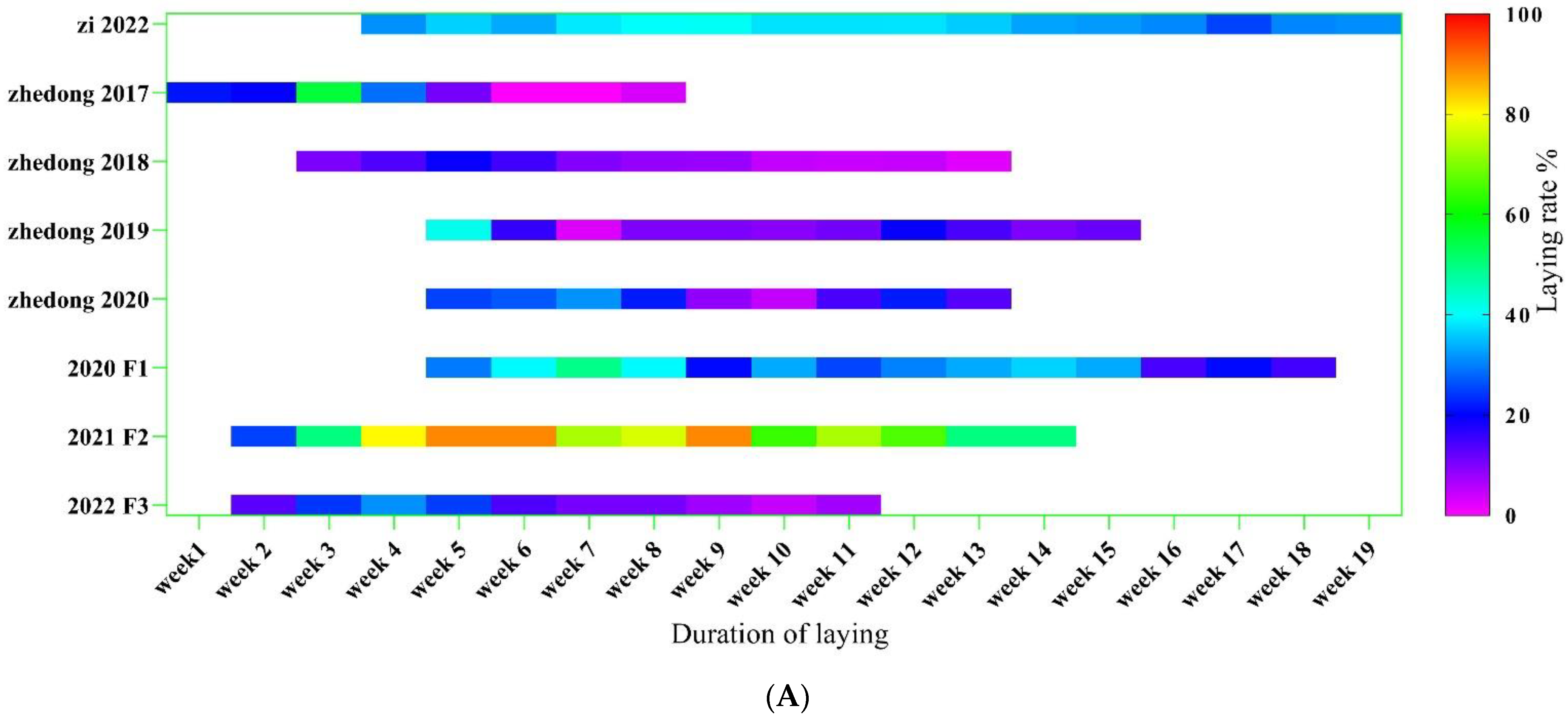
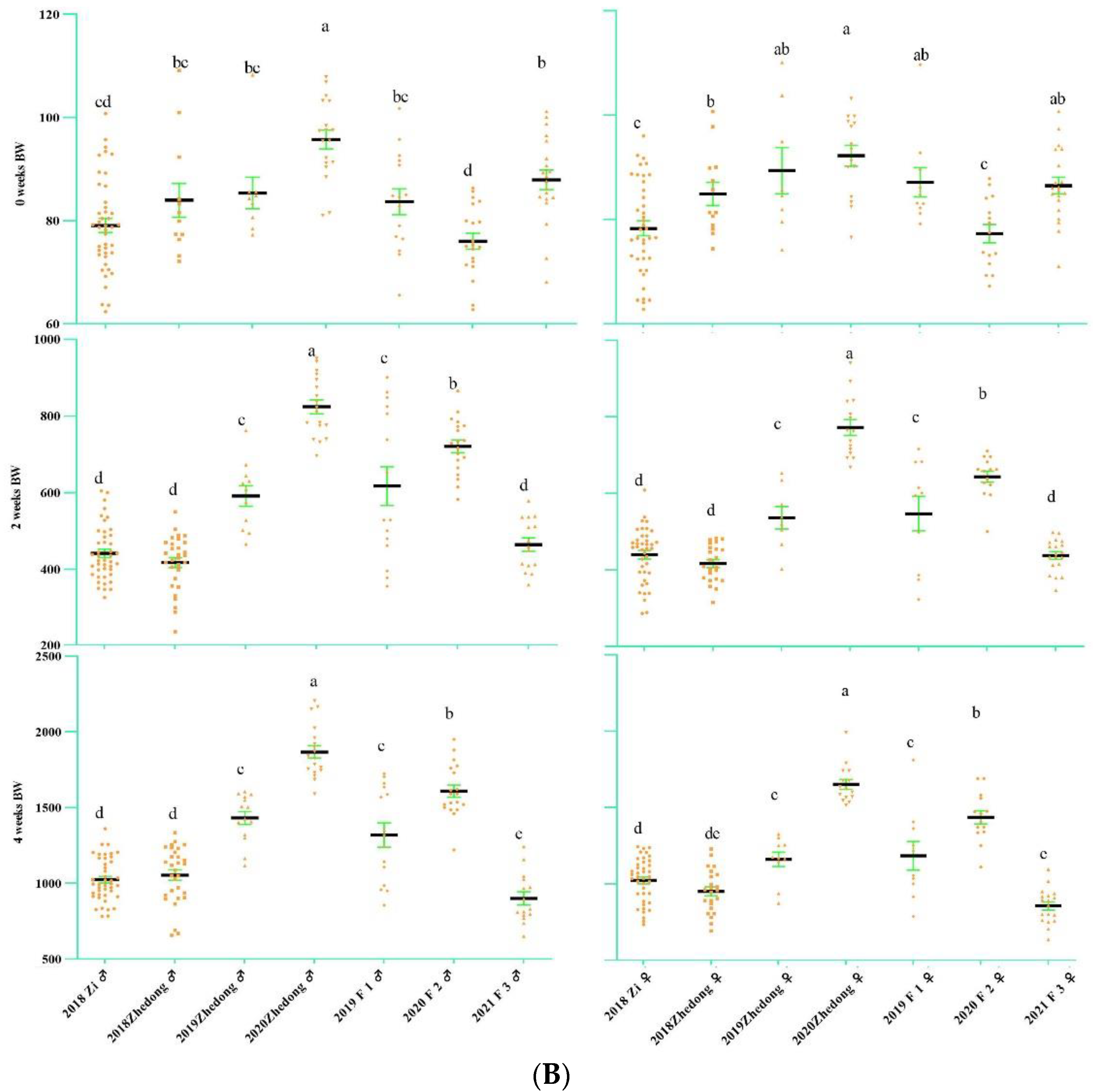
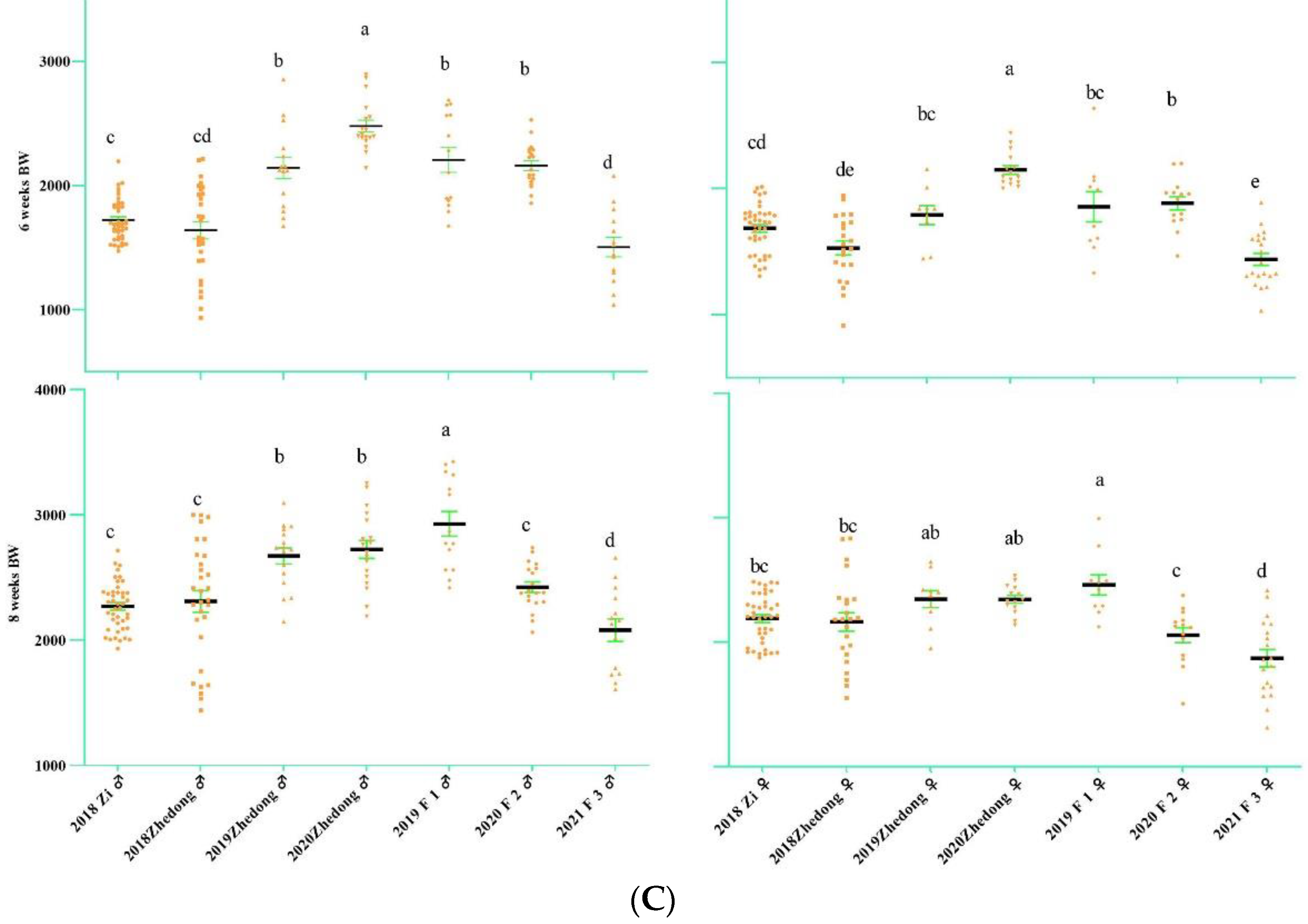
2.3. Body Weight and Egg-Laying Phenotypic Measurements
2.4. Genome Resequencing
2.5. SNP Calling and GWAS
2.6. Three-Dimensional (3D) Structure Prediction and Molecular Docking
3. Results
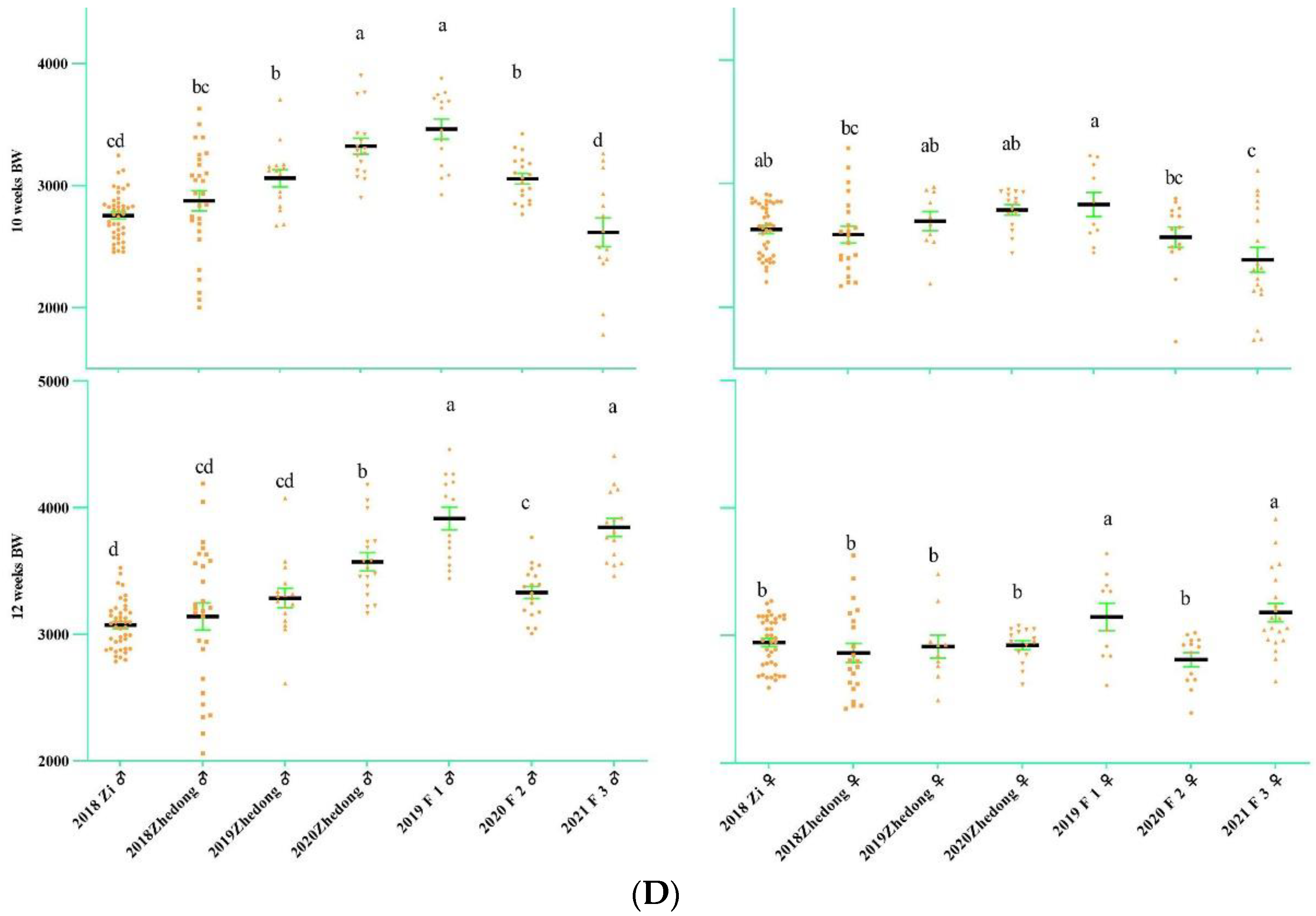
3.1. Analysis of Goose Body Weight and Egg-Laying Trait
3.2. Genome Resequencing and Whole Genome Resequencing
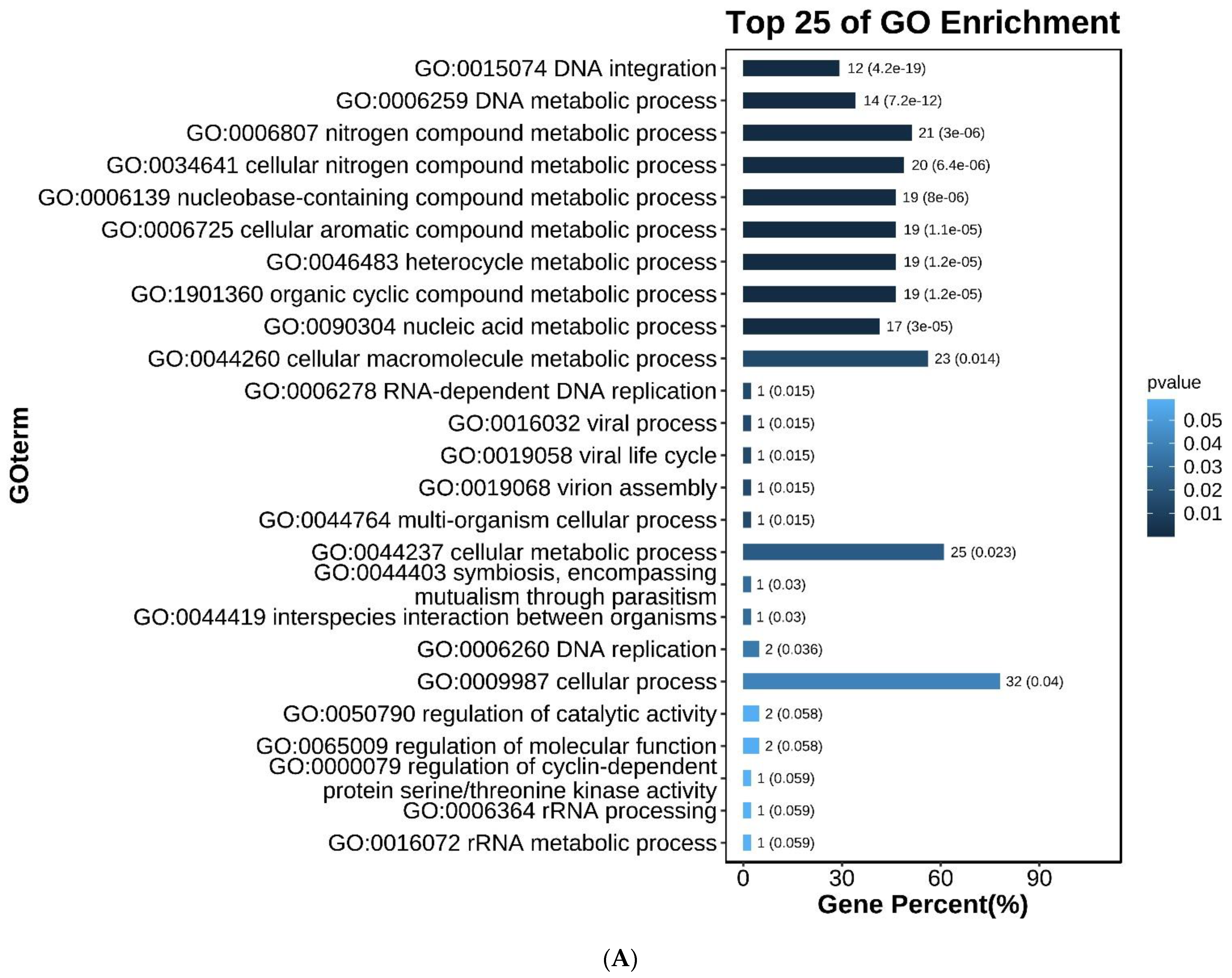
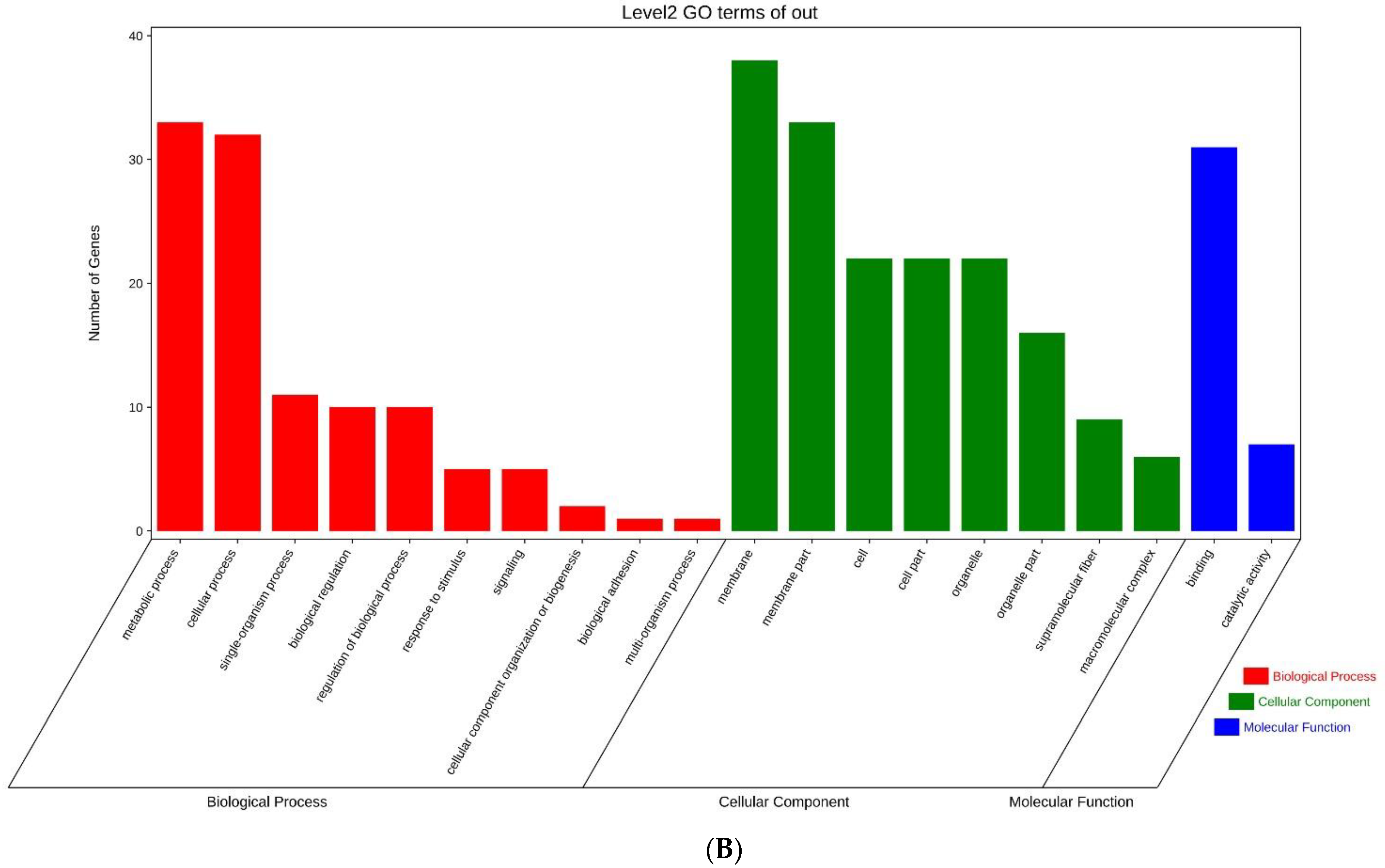
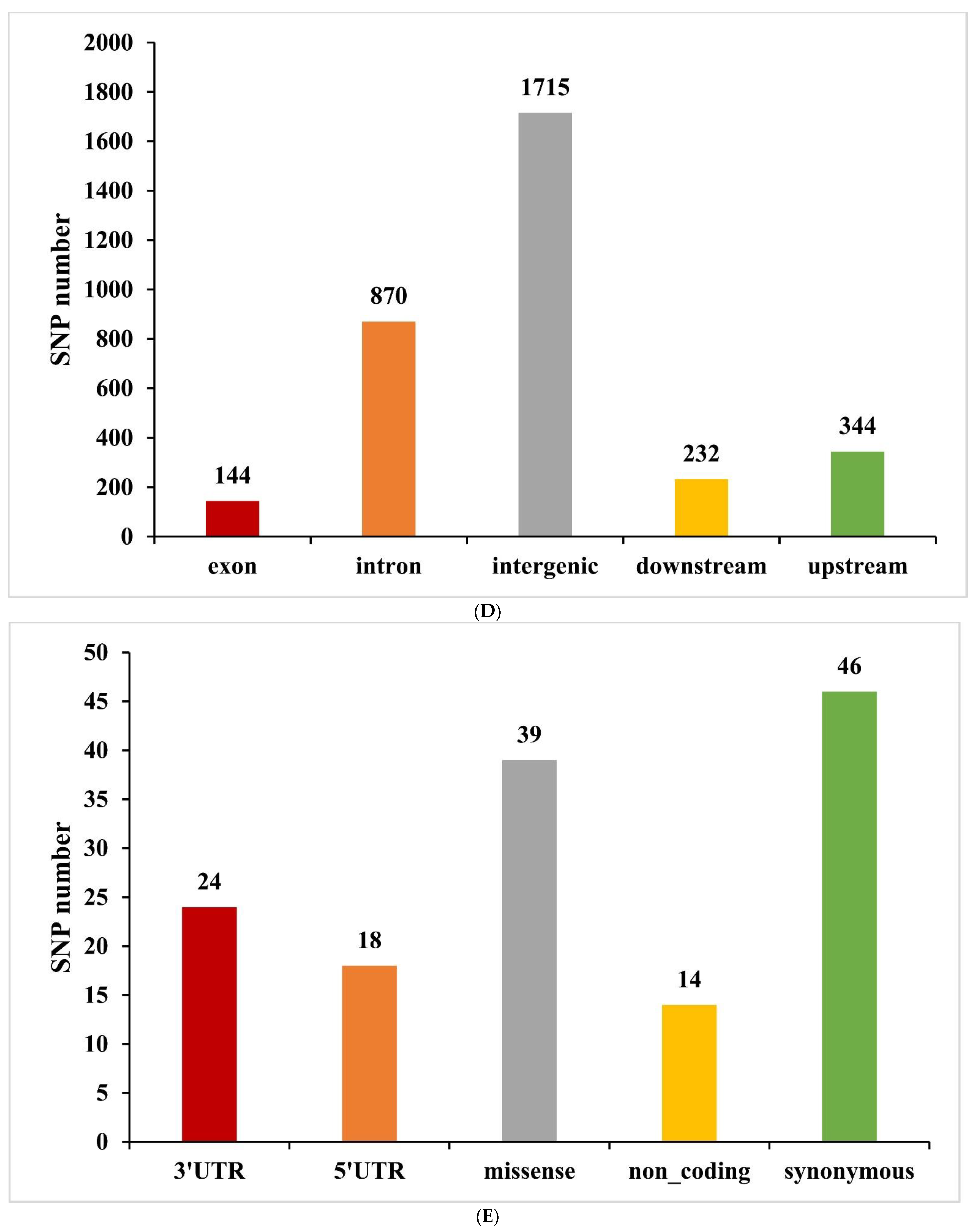
3.3. Screening and Analysis of Candidate SNPs
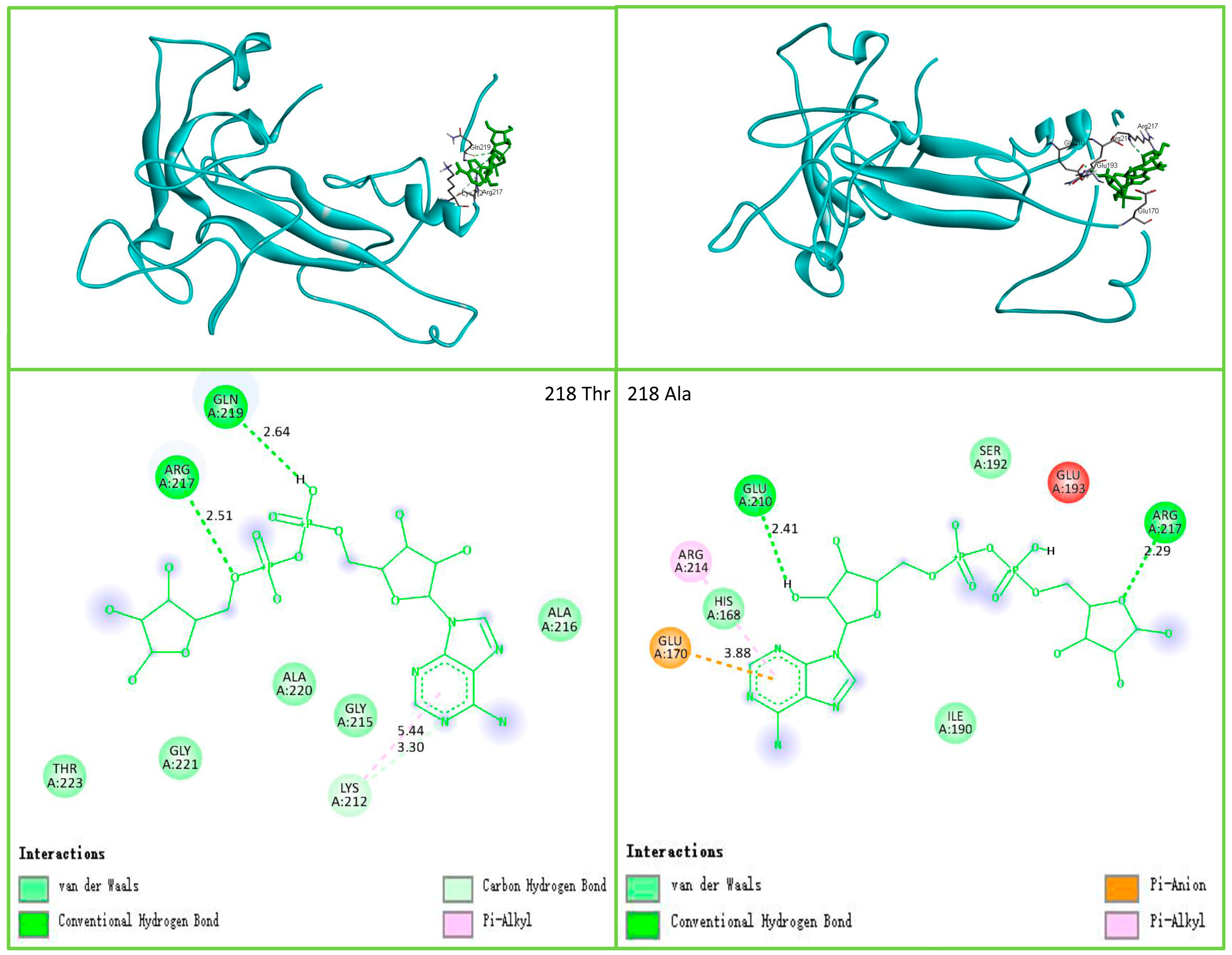
3.4. Candidate Genes’ 3D Structure Prediction and Molecular Docking
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yu, J.; Guo, C. Mitochondrial dysfunction in follicles is associated with broodiness in Zhedong white goose. Anim. Reprod. Sci. 2022, 243, 107032. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; Yang, Y.Z.; Gu, T.T.; Cao, Z.F.; Zhao, W.M.; Qin, H.R.; Xu, Q.; Chen, G.H. Comparison of the broody behavior characteristics of different breeds of geese. Poult. Sci. 2019, 98, 5226–5233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, Z.Y.; An, C.; Weng, K.Q.; Cao, Z.F.; Xu, Q.; Chen, G.H. Effect of active immunization with recombinant-derived goose INH-alpha, AMH, and PRL fusion protein on broodiness onset and egg production in geese (Anser cygnoides). Poult. Sci. 2021, 100, 101452. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, L.; Zhang, Y.; Yao, Y.; Zhao, W.; Xu, Q.; Chen, G. Characterization of ovarian morphology and reproductive hormones in Zhedong white geese (Anser cygnoides domesticus) during the reproductive cycle. J. Anim. Physiol. Anim. Nutr. 2021, 105, 938–945. [Google Scholar] [CrossRef]
- Liu, G.J.; Chen, Z.F.; Zhao, X.H.; Li, M.Y.; Guo, Z.H. Meta-analysis: Supplementary artificial light and goose reproduction. Anim. Reprod. Sci. 2020, 214, 106278. [Google Scholar] [CrossRef]
- Zhu, H.X.; Liu, X.Q.; Hu, M.D.; Lei, M.M.; Chen, Z.; Ying, S.J.; Yu, J.N.; Dai, Z.C.; Shi, Z.D. Endocrine and molecular regulation mechanisms of the reproductive system of Hungarian White geese investigated under two artificial photoperiodic programs. Theriogenology 2019, 123, 167–176. [Google Scholar] [CrossRef]
- Zhu, H.X.; Hu, M.D.; Guo, B.B.; Qu, X.L.; Lei, M.M.; Chen, R.; Chen, Z.; Shi, Z.D. Effect and molecular regulatory mechanism of monochromatic light colors on the egg-laying performance of Yangzhou geese. Anim. Reprod. Sci. 2019, 204, 131–139. [Google Scholar] [CrossRef]
- Ouyang, Q.; Hu, S.; Wang, G.; Hu, J.; Zhang, J.; Li, L.; Hu, B.; He, H.; Liu, H.; Xia, L.; et al. Comparative Transcriptome Analysis Suggests Key Roles for 5-Hydroxytryptamlne Receptors in Control of Goose Egg Production. Genes 2020, 11, 455. [Google Scholar] [CrossRef]
- Zhao, Q.; Chen, J.; Zhang, X.; Xu, Z.; Lin, Z.; Li, H.; Lin, W.; Xie, Q. Genome-Wide Association Analysis Reveals Key Genes Responsible for Egg Production of Lion Head Goose. Front. Genet. 2019, 10, 1391. [Google Scholar] [CrossRef]
- Ye, M.; Sun, L.; Yang, R.; Wang, Z.; Qi, K. The optimization of fermentation conditions for producing cellulase of Bacillus amyloliquefaciens and its application to goose feed. R. Soc. Open Sci. 2017, 4, 171012. [Google Scholar] [CrossRef]
- Melak, S.; Wang, Q.; Tian, Y.; Wei, W.; Zhang, L.; Elbeltagy, A.; Chen, J. Identification and Validation of Marketing Weight-Related SNP Markers Using SLAF Sequencing in Male Yangzhou Geese. Genes 2021, 12, 1203. [Google Scholar] [CrossRef] [PubMed]
- Abdel Moniem, H.; Yusuf, M.S.; Chen, G. Ecology and population structure of some indigenous geese breeds and the impact of four GH and Pit-1 SNPs on their body weights. Environ. Sci. Pollut. Res. Int. 2021, 28, 37603–37615. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Chen, P.; Zhou, C.; Zhao, X.; Zhang, K.; Wu, R.; Zhang, C.; Wang, Y.; Xie, Y.; Wang, Q. Genome-wide association study for reproduction-related traits in Chinese domestic goose. Br. Poult. Sci 2022, 63, 754–760. [Google Scholar] [CrossRef]
- Souza, C.J.; McNeilly, A.S.; Benavides, M.V.; Melo, E.O.; Moraes, J.C. Mutation in the protease cleavage site of GDF9 increases ovulation rate and litter size in heterozygous ewes and causes infertility in homozygous ewes. Anim. Genet. 2014, 45, 732–739. [Google Scholar] [CrossRef]
- Lee, S.H.; Lee, S.H.; Park, H.B.; Kim, J.M. Identification of key adipogenic transcription factors for the pork belly parameters via the association weight matrix. Meat Sci. 2023, 195, 109015. [Google Scholar] [CrossRef]
- Yuan, J.; Zhou, X.; Xu, G.; Xu, S.; Liu, B. Genetic diversity and population structure of Tongcheng pigs in China using whole-genome SNP chip. Front. Genet. 2022, 13, 910521. [Google Scholar] [CrossRef] [PubMed]
- Mesbah-Uddin, M.; Guldbrandtsen, B.; Capitan, A.; Lund, M.S.; Boichard, D.; Sahana, G. Genome-wide association study with imputed whole-genome sequence variants including large deletions for female fertility in 3 Nordic dairy cattle breeds. J. Dairy Sci. 2022, 105, 1298–1313. [Google Scholar] [CrossRef]
- Guo, Y.; Bai, F.; Wang, J.; Fu, S.; Zhang, Y.; Liu, X.; Zhang, Z.; Shao, J.; Li, R.; Wang, F.; et al. Design and characterization of a high-resolution multiple-SNP capture array by target sequencing for sheep. J. Anim. Sci. 2022, 101, skac383. [Google Scholar] [CrossRef]
- Moaeen-Ud-Din, M.; Danish Muner, R.; Khan, M.S. Genome wide association study identifies novel candidate genes for growth and body conformation traits in goats. Sci. Rep. 2022, 12, 9891. [Google Scholar] [CrossRef]
- Li, J.; Wang, Z.; Lubritz, D.; Arango, J.; Fulton, J.; Settar, P.; Rowland, K.; Cheng, H.; Wolc, A. Genome-wide association studies for egg quality traits in White Leghorn layers using low-pass sequencing and SNP chip data. J. Anim. Breed. Genet. 2022, 139, 380–397. [Google Scholar] [CrossRef]
- Yuan, X.; Lan, G.; Li, L.; He, H.; Wang, J.; Hu, S. Differential gene expression profiling of the goose pineal gland. Br. Poult. Sci. 2020, 61, 200–208. [Google Scholar] [CrossRef] [PubMed]
- Gao, G.; Gao, D.; Zhao, X.; Xu, S.; Zhang, K.; Wu, R.; Yin, C.; Li, J.; Xie, Y.; Hu, S.; et al. Genome-Wide Association Study-Based Identification of SNPs and Haplotypes Associated With Goose Reproductive Performance and Egg Quality. Front. Genet. 2021, 12, 602583. [Google Scholar] [CrossRef] [PubMed]
- Farrar, V.S.; Flores, L.; Viernes, R.C.; Ornelas Pereira, L.; Mushtari, S.; Calisi, R.M. Prolactin promotes parental responses and alters reproductive axis gene expression, but not courtship behaviors, in both sexes of a biparental bird. Horm. Behav. 2022, 144, 105217. [Google Scholar] [CrossRef] [PubMed]
- Norris, A.R.; Martin, K.; Cockle, K.L. Weather and nest cavity characteristics influence fecundity in mountain chickadees. PeerJ 2022, 10, e14327. [Google Scholar] [CrossRef] [PubMed]
- Morelli, F.; Benedetti, Y.; Pape Moller, A. Diet specialization and brood parasitism in cuckoo species. Ecol. Evol. 2020, 10, 5097–5105. [Google Scholar] [CrossRef]
- Chen, F.; Li, J.; Zhang, H.; Xu, J.; Tao, Z.; Shen, J.; Shen, J.; Lu, L.; Li, C. Identification of differentially expressed known and novel miRNAs in broodiness of goose. Mol. Biol. Rep. 2014, 41, 2767–2777. [Google Scholar] [CrossRef]
- Yu, J.; Lou, Y.; Zhao, A. Transcriptome analysis of follicles reveals the importance of autophagy and hormones in regulating broodiness of Zhedong white goose. Sci. Rep. 2016, 6, 36877. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Li, L.; Han, C.; He, H.; Xu, H. Transcriptome analysis revealed the possible regulatory pathways initiating female geese broodiness within the hypothalamic-pituitary-gonadal axis. PLoS ONE 2018, 13, e0191213. [Google Scholar] [CrossRef]
- Lou, Y.; Yu, W.; Han, L.; Yang, S.; Wang, Y.; Ren, T.; Yu, J.; Zhao, A. ROS activates autophagy in follicular granulosa cells via mTOR pathway to regulate broodiness in goose. Anim. Reprod. Sci. 2017, 185, 97–103. [Google Scholar] [CrossRef]
- Qin, H.; Li, X.; Wang, J.; Sun, G.; Mu, X.; Ji, R. Ovarian transcriptome profile from pre-laying period to broody period of Xupu goose. Poult. Sci. 2021, 100, 101403. [Google Scholar] [CrossRef]
- Hou, L.; Ji, W.; Gu, T.; Weng, K.; Liu, D.; Zhang, Y.; Zhang, Y.; Xu, Q.; Chen, G. MiR-34c-5p promotes granulosa cells apoptosis by targeting Bcl2 in broody goose (Anser cygnoides). Anim. Biotechnol. 2022, 33, 1280–1288. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Lou, Y.; He, K.; Yang, S.; Yu, W.; Han, L.; Zhao, A. Goose broodiness is involved in granulosa cell autophagy and homeostatic imbalance of follicular hormones. Poult. Sci. 2016, 95, 1156–1164. [Google Scholar] [CrossRef]
- Li, K.; Liu, T. Evaluation of Oncogene NUP37 as a Potential Novel Biomarker in Breast Cancer. Front. Oncol. 2021, 11, 669655. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Liu, Y.; Feng, W.; Lei, L.; Du, Y.; Wu, J.; Wang, S. NUP37, a positive regulator of YAP/TEAD signaling, promotes the progression of hepatocellular carcinoma. Oncotarget 2017, 8, 98004–98013. [Google Scholar] [CrossRef] [PubMed]
- Esteves de Lima, J.; Bonnin, M.A.; Birchmeier, C.; Duprez, D. Muscle contraction is required to maintain the pool of muscle progenitors via YAP and NOTCH during fetal myogenesis. Elife 2016, 5, e15593. [Google Scholar] [CrossRef]
- Cao, X.; Pfaff, S.L.; Gage, F.H. YAP regulates neural progenitor cell number via the TEA domain transcription factor. Genes Dev. 2008, 22, 3320–3334. [Google Scholar] [CrossRef]
- Rudolf, M.A.; Andreeva, A.; Kozlowski, M.M.; Kim, C.E.; Moskowitz, B.A.; Anaya-Rocha, A.; Kelley, M.W.; Corwin, J.T. YAP Mediates Hair Cell Regeneration in Balance Organs of Chickens, But LATS Kinases Suppress Its Activity in Mice. J. Neurosci. 2020, 40, 3915–3932. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.Y.; Chen, T.H.; Wen, P.Y.; Chou, C.H.; Ying, T.H.; Chang, S.P.; Ma, G.C.; Chen, M. Differential expression of NUDT9 at different phases of the menstrual cycle and in different components of normal and neoplastic human endometrium. Taiwan J. Obstet. Gynecol. 2009, 48, 96–107. [Google Scholar] [CrossRef]
- Wright, R.H.G.; Beato, M. Role of the NUDT Enzymes in Breast Cancer. Int. J. Mol. Sci. 2021, 22, 2267. [Google Scholar] [CrossRef]
- Gattkowski, E.; Rutherford, T.J.; Mockl, F.; Bauche, A.; Sander, S.; Fliegert, R.; Tidow, H. Analysis of ligand binding and resulting conformational changes in pyrophosphatase. NUDT9. FEBS J. 2021, 288, 6769–6782. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Content | 0–4 Week | 5–10 Week | 11–28 Week | Adult Goose (Winter) | Egg-Laying Goose |
|---|---|---|---|---|---|
| Energy MJ/kg | 11.7 | 11.7 | 11.1 | 10.7 | 11.3 |
| Crude protein % | 18 | 15.5 | 12 | 8 | 15.5 |
| Ca% | 1 | 1.2 | 1.2 | 1.2 | 2.35 |
| Lysine % | 1 | 1 | 0.7 | 0.7 | 0.75 |
| Sample | Total Reads | Unaligned Reads | Uniquely Aligned Reads | Clean_Paired_Reads | GC (%) | Q30 (%) |
|---|---|---|---|---|---|---|
| B311 | 73,532,158 | 7,269,362 (9.88596%) | 63,142,228 (85.8702%) | 36,787,767 | 42% | 95.67% |
| B314 | 72,163,278 | 6,729,461 (9.32533%) | 62,789,055 (87.0097%) | 36,081,639 | 42% | 95.68% |
| B316 | 73,688,664 | 6,925,219 (9.39794%) | 63,955,897 (86.792%) | 36,844,332 | 42% | 95.66% |
| B319 | 73,511,268 | 6,445,136 (8.76755%) | 64,116,644 (87.2202%) | 36,755,634 | 42% | 96.24% |
| B320 | 73,829,012 | 6,502,035 (8.80688%) | 64,631,140 (87.5417%) | 36,914,506 | 42% | 95.57% |
| C401 | 73,517,946 | 6,853,528 (9.32225%) | 63,752,893 (86.7175%) | 36,758,973 | 42% | 96.40% |
| C402 | 73,428,654 | 7,519,679 (10.2408%) | 62,603,346 (85.2574%) | 36,714,327 | 42% | 96.79% |
| C407 | 73,539,408 | 6,771,803 (9.2084%) | 63,947,268 (86.9565%) | 36,769,704 | 42% | 96.35% |
| C411 | 65,064,624 | 6,636,293 (10.1995%) | 55,704,018 (85.6134%) | 32,532,312 | 42% | 91.20% |
| C412 | 73,430,222 | 6,913,259 (9.41473%) | 63,126,236 (85.9676%) | 36,715,111 | 42% | 96.10% |
| O1102 | 73,749,910 | 6,707,802 (9.09534%) | 64,160,474 (86.9974%) | 36,874,955 | 42% | 95.32% |
| O1112 | 73,671,940 | 6,572,557 (8.92138%) | 64,163,844 (87.094%) | 36,835,970 | 42% | 95.45% |
| O1114 | 73,888,590 | 6,678,570 (9.0387%) | 64,367,737 (87.1146%) | 36,944,295 | 42% | 95.67% |
| O1116 | 73,820,700 | 6,682,537 (9.05239%) | 64,336,901 (87.1529%) | 36,910,350 | 42% | 95.52% |
| O1125 | 73,684,588 | 6,743,275 (9.15154%) | 64,208,342 (87.1394%) | 36,842,294 | 42% | 95.68% |
| CHROM | POS | Gene_ID | Transcript_ID | Codan_Mutate | AA_Mutate | |
|---|---|---|---|---|---|---|
| SNP 1 | NW_013185657.1 | 483,398 | 106040530 | XM_013188477.1 | c.479G > A | Arg160Gln |
| SNP 2 | NW_013185677.1 | 5,511,334 | 106033575 | XM_013177200.1 | c.3523C > A | Gln1175Lys |
| SNP 3 | NW_013185696.1 | 595 | 106035773 | XM_013180816.1 | c.912G > A | Met304Ile |
| SNP 4 | NW_013185696.1 | 615 | 106035773 | XM_013180816.1 | c.892T > G | Phe298Val |
| SNP 5 | NW_013185720.1 | 3,081,938 | 106038401 | XM_013184886.1 | c.977G > A | Cys326Tyr |
| SNP 6 | NW_013185720.1 | 3,082,790 | 106038401 | XM_013184889.1 | c.245G > A | Arg82Gln |
| SNP 7 | NW_013185721.1 | 2,609,554 | 106038498 | XM_013185039.1 | c.1637A > G | Gln546Arg |
| SNP 8 | NW_013185748.1 | 3,578,791 | 106040672 | XM_013188695.1 | c.522C > G | Phe174Leu |
| SNP 9 | NW_013185750.1 | 69,680 | 106040872 | XM_013189032.1 | c.1442T > G | Ile481Ser |
| SNP 10 | NW_013185760.1 | 14,289 | 106041536 | XM_013190069.1 | c.3349G > T | Val1117Phe |
| SNP 11 | NW_013185766.1 | 956,541 | 106041864 | XM_013190505.1 | c.652A > G | Thr218Ala |
| SNP 12 | NW_013185766.1 | 956,750 | 106041864 | XM_013190505.1 | c.861A > G | Ter287Ter |
| SNP 13 | NW_013185800.1 | 639,495 | 106043465 | XM_013192926.1 | c.290G > A | Arg97Lys |
| SNP 14 | NW_013185811.1 | 2,202,032 | 106043881 | XM_013193607.1 | c.509G > C | Arg170Thr |
| SNP 15 | NW_013185811.1 | 2,202,034 | 106043881 | XM_013193607.1 | c.507G > C | Glu169Asp |
| SNP 16 | NW_013185864.1 | 1,205,891 | 106045958 | XM_013196755.1 | c.298C > T | Leu100Phe |
| SNP 17 | NW_013185870.1 | 1,328,057 | 106046235 | XM_013197144.1 | c.353C > T | Ala118Val |
| SNP 18 | NW_013185914.1 | 62,943 | 106047476 | XM_013198970.1 | c.561A > C | Arg187Ser |
| SNP 19 | NW_013185914.1 | 63,115 | 106047476 | XM_013198970.1 | c.389A > G | Glu130Gly |
| SNP 20 | NW_013185915.1 | 942,834 | 106047517 | XM_013199056.1 | c.163A > G | Met55Val |
| SNP 21 | NW_013185915.1 | 942,936 | 106047517 | XM_013199056.1 | c.265A > G | Thr89Ala |
| SNP 22 | NW_013185925.1 | 822,558 | 106047720 | XM_013199342.1 | c.512G > A | Arg171His |
| SNP 23 | NW_013186010.1 | 166,638 | 106049132 | XM_013201311.1 | c.92G > C | Gly31Ala |
| SNP 24 | NW_013186010.1 | 166,674 | 106049132 | XM_013201311.1 | c.56G > A | Arg19His |
| SNP 25 | NW_013186010.1 | 261,219 | 106049145 | XM_013201321.1 | c.25G > A | Ala9Thr |
| SNP 26 | NW_013186017.1 | 261,566 | 106049239 | XM_013201442.1 | c.95A > C | Tyr32Ser |
| SNP 27 | NW_013186061.1 | 144,964 | 106049600 | XM_013201966.1 | c.134A > C | Asn45Thr |
| SNP 28 | NW_013186074.1 | 48,559 | 106049689 | XM_013202086.1 | c.620G > A | Arg207His |
| SNP 29 | NW_013186089.1 | 144,070 | 106049779 | XM_013202193.1 | c.13A > T | Thr5Ser |
| SNP 30 | NW_013186111.1 | 71,554 | 106049894 | XM_013202325.1 | c.422C > A | Thr141Asn |
| SNP 31 | NW_013186111.1 | 71,555 | 106049894 | XM_013202325.1 | c.421A > C | Thr141Pro |
| SNP 32 | NW_013186150.1 | 35,471 | 106029399 | XM_013170682.1 | c.397G > A | Ala133Thr |
| SNP 33 | NW_013186231.1 | 6,643 | 106029521 | XM_013170813.1 | c.349G>T | Ala117Ser |
| SNP 34 | NW_013186264.1 | 9,000 | 106029548 | XM_013170832.1 | c.303G > C | Leu101Phe |
| SNP 35 | NW_013186857.1 | 861 | 106029642 | XM_013170927.1 | c.680T > C | Ile227Thr |
| SNP 36 | NW_013187000.1 | 470 | 106029654 | XM_013170934.1 | c.200T > C | Ile67Thr |
| SNP 37 | NW_013187000.1 | 667 | 106029654 | XM_013170934.1 | c.397A > G | Ile133Val |
| SNP 38 | NW_013187000.1 | 1,090 | 106029654 | XM_013170934.1 | c.820G > A | Val274Ile |
| SNP 39 | NW_013188772.1 | 217 | 106029704 | XM_013170974.1 | c.341G > A | Gly114Asp |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, G.; Guo, Z.; Zhao, X.; Sun, J.; Yue, S.; Li, M.; Chen, Z.; Ma, Z.; Zhao, H. Whole Genome Resequencing Identifies Single-Nucleotide Polymorphism Markers of Growth and Reproduction Traits in Zhedong and Zi Crossbred Geese. Genes 2023, 14, 487. https://doi.org/10.3390/genes14020487
Liu G, Guo Z, Zhao X, Sun J, Yue S, Li M, Chen Z, Ma Z, Zhao H. Whole Genome Resequencing Identifies Single-Nucleotide Polymorphism Markers of Growth and Reproduction Traits in Zhedong and Zi Crossbred Geese. Genes. 2023; 14(2):487. https://doi.org/10.3390/genes14020487
Chicago/Turabian StyleLiu, Guojun, Zhenhua Guo, Xiuhua Zhao, Jinyan Sun, Shan Yue, Manyu Li, Zhifeng Chen, Zhigang Ma, and Hui Zhao. 2023. "Whole Genome Resequencing Identifies Single-Nucleotide Polymorphism Markers of Growth and Reproduction Traits in Zhedong and Zi Crossbred Geese" Genes 14, no. 2: 487. https://doi.org/10.3390/genes14020487
APA StyleLiu, G., Guo, Z., Zhao, X., Sun, J., Yue, S., Li, M., Chen, Z., Ma, Z., & Zhao, H. (2023). Whole Genome Resequencing Identifies Single-Nucleotide Polymorphism Markers of Growth and Reproduction Traits in Zhedong and Zi Crossbred Geese. Genes, 14(2), 487. https://doi.org/10.3390/genes14020487