
Advancements and Challenges in Ancient DNA Research: Bridging the Global North–South Divide
 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
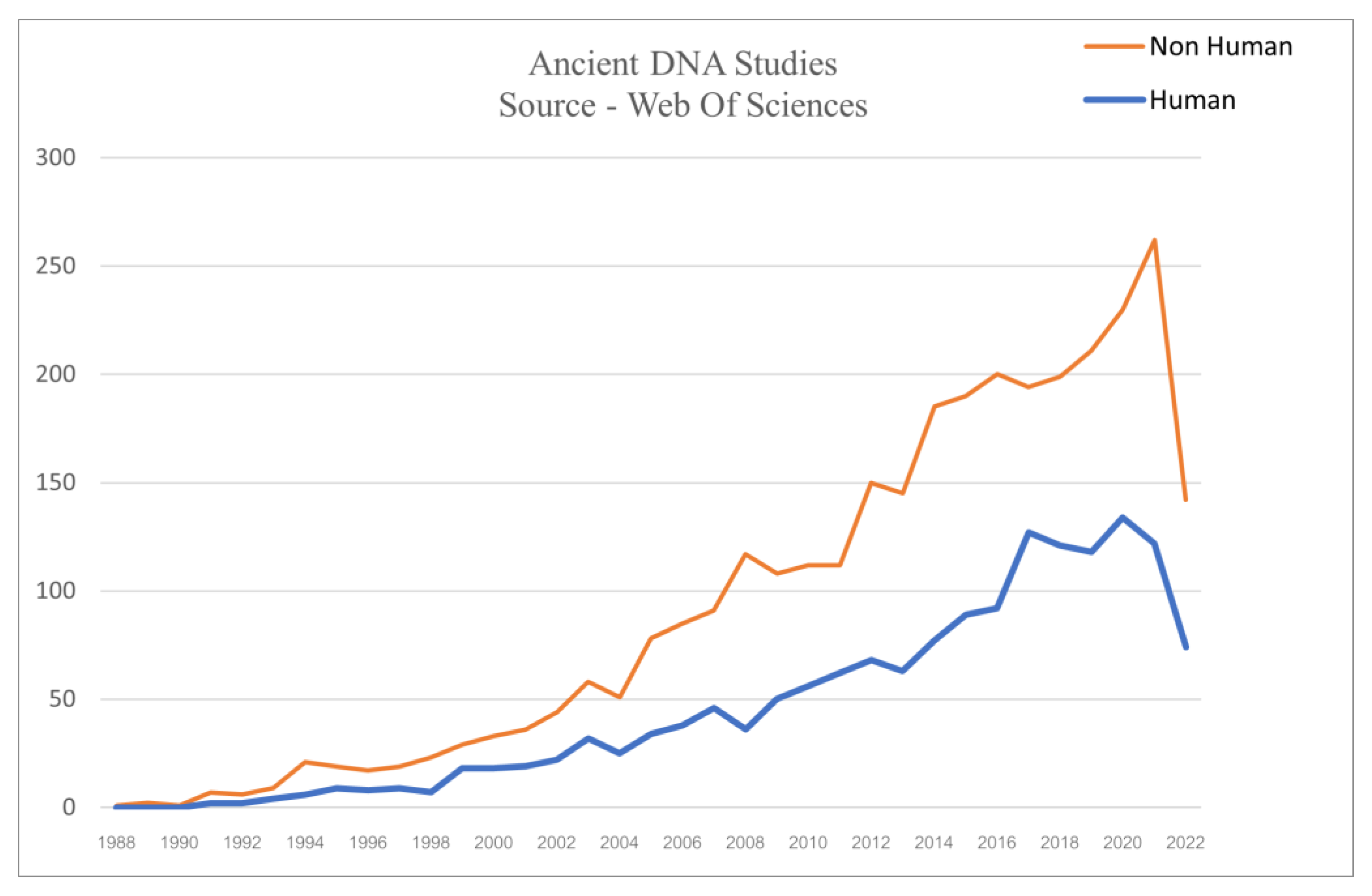
2. Rise of aDNA Field
2.1. Species Phylogenies
2.2. Migration and Phylogeography
2.3. Archaic Hominis
2.4. Paleopathology
2.5. Adaptation
3. Technological Advancements
3.1. Revolutionized Extraction Methodologies
3.2. Evolution in Sequencing Technology
3.3. Advancements in Data Analyzing Tools
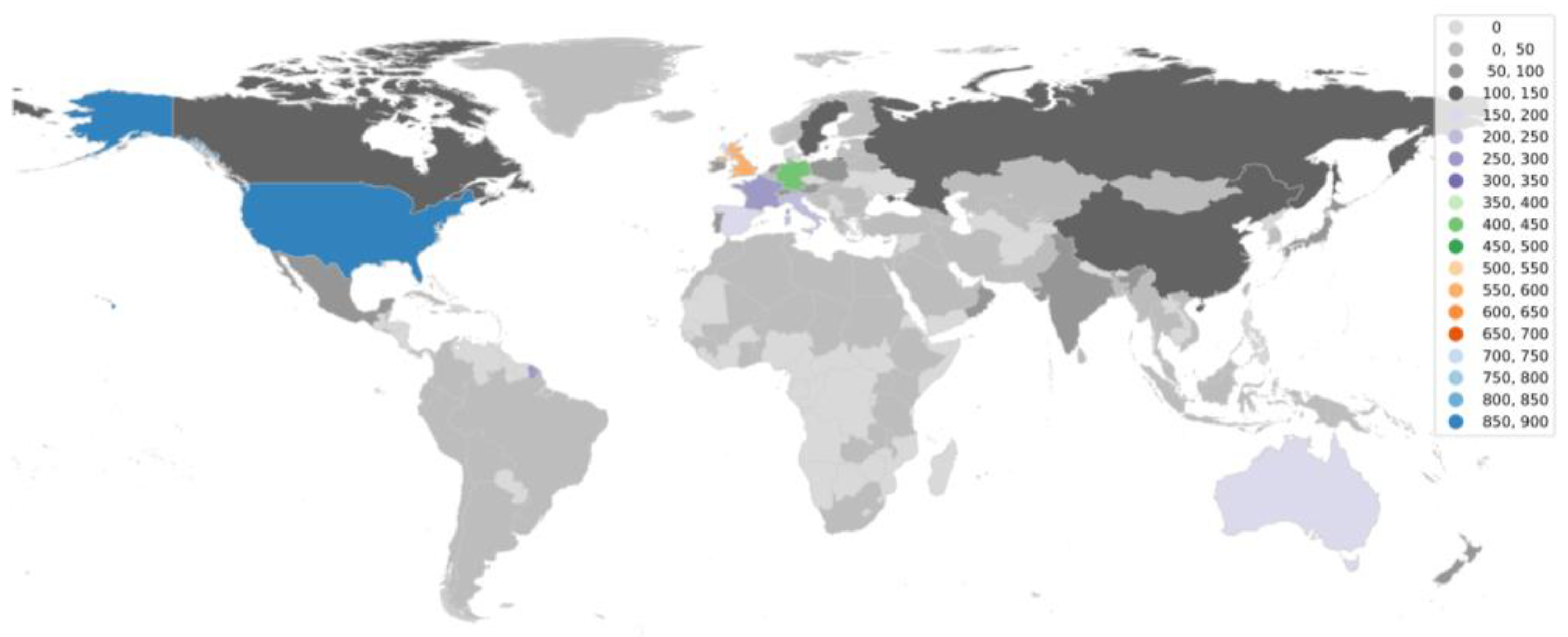
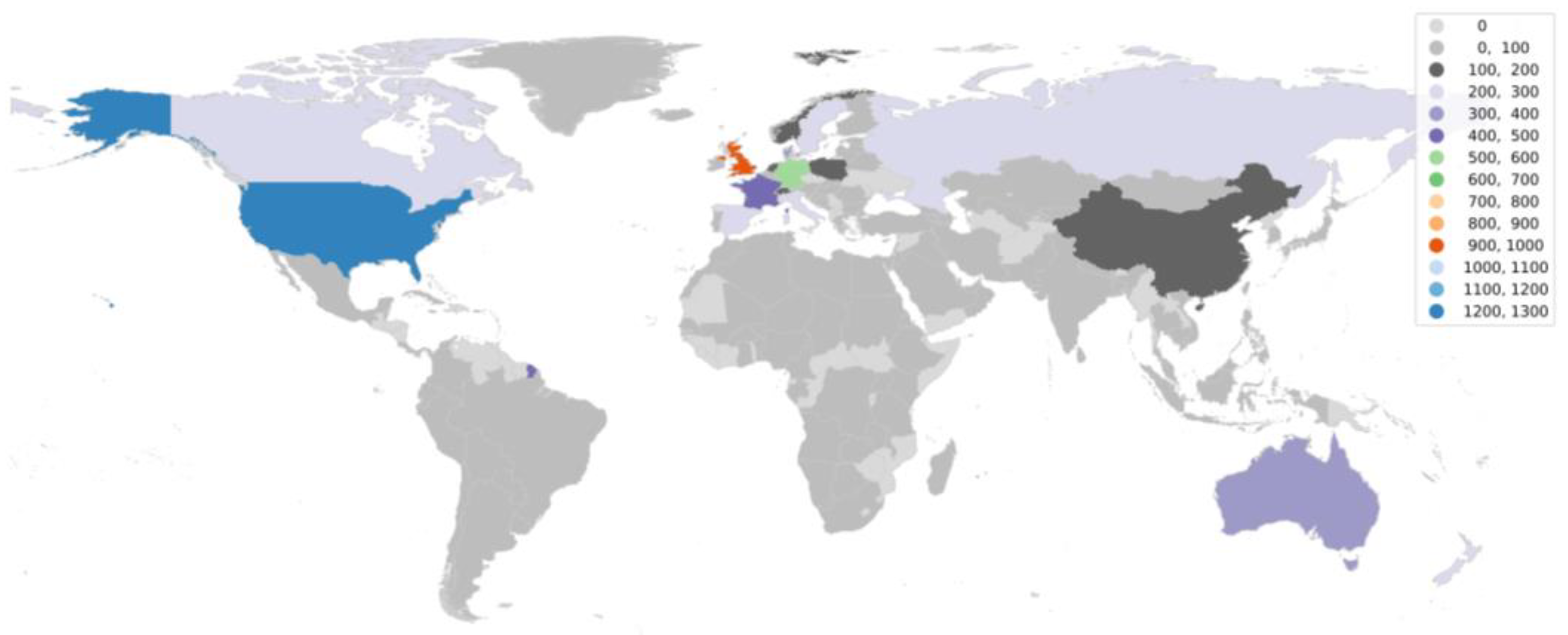
4. Dominance of Global North in aDNA Research
5. Challenges to Overcome
5.1. Availability of aDNA from the Ancient Biological Samples
5.2. Contamination and Authenticity of aDNA
5.3. Ethical Concerns
6. Mapping out the Future Direction
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Darwin, C. On the Origin of the Species; Routledge: London, UK, 1859. [Google Scholar]
- Higuchi, R.; Bowman, B.; Freiberger, M.; Ryder, O.A.; Wilson, A.C. DNA sequences from the quagga, an extinct member of the horse family. Nature 1984, 312, 282–284. [Google Scholar] [CrossRef] [PubMed]
- Pääbo, S. Molecular cloning of Ancient Egyptian mummy DNA. Nature 1985, 314, 644–645. [Google Scholar] [CrossRef] [PubMed]
- Saiki, R.K.; Scharf, S.; Faloona, F.; Mullis, K.B.; Horn, G.T.; Erlich, H.A.; Arnheim, N. Enzymatic Amplification of β-Globin Genomic Sequences and Restriction Site Analysis for Diagnosis of Sickle Cell Anemia. Science 1985, 230, 1350–1354. [Google Scholar] [CrossRef] [PubMed]
- Saiki, R.K.; Gelfand, D.H.; Stoffel, S.; Scharf, S.J.; Higuchi, R.; Horn, G.T.; Mullis, K.B.; Erlich, H.A. Primer-Directed Enzymatic Amplification of DNA with a Thermostable DNA Polymerase. Science 1988, 239, 487–491. [Google Scholar] [CrossRef]
- Spyrou, M.A.; Musralina, L.; Ruscone, G.A.; Kocher, A.; Borbone, P.; Khartanovich, V.I.; Buzhilova, A.; Djansugurova, L.; Bos, D.K.; Kühnert, D.; et al. The source of the Black Death in fourteenth-century central Eurasia. Nature 2022, 606, 718–724. [Google Scholar] [CrossRef]
- Lindo, J.; Rosa, R.; Santos, A.; Sans, M.; DeGiorgio, M.; Figueiro, G. The genomic prehistory of the Indigenous peoples of Uruguay. PNAS Nexus 2022, 1, pgac047. [Google Scholar] [CrossRef]
- Reich, D.; Green, R.E.; Kircher, M.; Krause, J.; Patterson, N.; Durand, E.Y.; Viola, B.; Briggs, A.W.; Udo, S.; Johnson, P.L.F.; et al. Genetic history of an archaic hominin group from Denisova Cave in Siberia. Nature 2010, 468, 1053–1060. [Google Scholar] [CrossRef]
- Rasmussen, M.; Li, Y.; Lindgreen, S.; Pedersen, J.S.; Albrechtsen, A.; Moltke, I.; Metspalu, M.; Metspalu, E.; Kivisild, T.; Gupta, R.; et al. Ancient human genome sequence of an extinct Palaeo-Eskimo. Nature 2010, 463, 757–762. [Google Scholar] [CrossRef]
- Green, R.E.; Krause, J.; Briggs, A.W.; Maricic, T.; Stenzel, U.; Kircher, M.; Patterson, N.; Li, H.; Zhai, W.; Fritz, M.; et al. A draft sequence of the neandertal genome. Science 2010, 328, 710–722. [Google Scholar] [CrossRef]
- Margaryan, A.; Lawson, D.J.; Sikora, M.; Racimo, F.; Rasmussen, S.; Mooltke, I.; Cassidy, L.M.; Jorsboe, E.; Ingason, A.; Pedersen, M.K.; et al. Population genomics of the Viking world. Nature 2020, 585, 390–396. [Google Scholar] [CrossRef]
- Barlow, A.; Cahill, J.A.; Hartmann, S.; Theunert, C.; Xenikoudakis, G.; Fortes, G.G.; Pajimans, J.L.A.; Rabeder, G.; Frischauf, C.; Grandal-d’Anglade, A.; et al. Partial genomic survival of cave bears in living brown bears. Nat. Ecol. Evol. 2018, 2, 1563–1570. [Google Scholar] [CrossRef]
- Palkopoulou, E.; Mallick, S.; Skoglund, P.; Enk, J.; Rohland, N.; Li, H.; Omrak, A.; Vartanyan, S.; Poinar, H.; Götherström, A.; et al. Complete Genomes Reveal Signatures of Demographic and Genetic Declines in the Woolly Mammoth. Curr. Biol. 2015, 25, 1395–1400. [Google Scholar] [CrossRef] [PubMed]
- Orlando, L.; Allaby, R.; Skoglund, P.; Sarkissian, C.; Stockhammer, P.W.; Ávila-Arcos, M.C.; Fu, Q.; Krause, J.; Willerslev, E.; Stone, A.C.; et al. Ancient DNA analysis. Nat. Rev. Methods Primers 2021, 1, 14. [Google Scholar] [CrossRef]
- Spyrou, M.A.; Bos, K.I.; Herbig, A.; Krause, J. Ancient pathogen genomics as an emerging tool for infectious disease research. Nat. Rev. Genet. 2019, 20, 323–340. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, R.; Akey, J.M.; Jakobsson, M.; Pritchard, J.K.; Tishkoff, S.; Willerslev, E. Tracing the peopling of the world through genomics. Nature 2017, 541, 302–310. [Google Scholar] [CrossRef]
- Rai, N.; Taher, N.; Singh, M.; Chaubey, G.; Jha, A.N.; Singh, L.; Thangaraj, K. Relic excavated in western India is probably of Georgian Queen Ketevan. Mitochondrion 2014, 14, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, M.L.; Buenrostro, J.D.; Valdiosera, C.; Schroeder, H.; Allentoft, M.E.; Sikora, M.; Rasmussen, M.; Gravel, S.; Guillén, S.; Nekhrizov, G.; et al. Pulling out the 1%: Whole-genome capture for the targeted enrichment of ancient DNA sequencing libraries. Am. J. Hum. Genet. 2013, 93, 852–864. [Google Scholar] [CrossRef]
- Rohland, N.; Siedel, H.; Hofreiter, M. A rapid column-based ancient DNA extraction method for increased sample throughput. Mol. Ecol. Resour. 2010, 10, 677–683. [Google Scholar] [CrossRef]
- Dabney, J.; Knapp, M.; Glocke, I.; Gansauge, M.; Weihmann, A.; Nickel, B.; Valdiosera, C.; García, N.; Pääbo, S.; Arsuage, J.; et al. Complete mitochondrial genome sequence of a Middle Pleistocene cave bear reconstructed from ultrashort DNA fragments. Proc. Natl. Acad. Sci. USA 2013, 110, 15758–15763. [Google Scholar] [CrossRef]
- Adler, C.J.; Haak, W.; Donlon, D.; Cooper, A. Survival and recovery of DNA from ancient teeth and bones. J. Archaeol. Sci. 2011, 38, 956–964. [Google Scholar] [CrossRef]
- Hansen, H.B.; Damgaard, P.B.; Margaryan, A.; Stenderup, J.; Lynnerup, N.; Willerslev, E.; Allentoft, M.E. Comparing Ancient DNA Preservation in Petrous Bone and Tooth Cementum. PLoS ONE 2017, 12, e0170940. [Google Scholar] [CrossRef]
- Pinhasi, R.; Fernandes, D.; Sirak, K.; Novak, M.; Connell, S.; Alpaslan-Roodenberg, S.; Gerritsen, F.; Moiseyev, V.; Gromov, A.; Raczky, P.; et al. Optimal Ancient DNA Yields from the Inner Ear Part of the Human Petrous Bone. PLoS ONE 2015, 10, e0129102. [Google Scholar] [CrossRef]
- Burbano, H.A.; Hodges, E.; Green, R.E.; Briggs, A.W.; Krause, J.; Meyer, M.; Good, J.M.; Maricic, T.; Johnson, P.L.F.; Xuan, Z.; et al. Targeted investigation of the Neandertal genome by array-based sequence capture. Science 2010, 328, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Krajewski, C.; Buckley, L.; Westerman, M. DNA phylogeny of the marsupial wolf resolved. Proc. R. Soc. Lond. B Biol. Sci. 1997, 264, 911–917. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.; Lalueza-Fox, C.; Anderson, S.; Rambaut, A.; Austin, J.; Ward, R. Complete mitochondrial genome sequences of two extinct moas clarify ratite evolution. Nature 2001, 409, 704–707. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, A.D.; Castresana, J.; Feldmaier-Fuchs, G.; Pääbo, S. A Molecular Phylogeny of Two Extinct Sloths. Mol. Phylogenet. Evol. 2001, 18, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, E.E.; James, H.F.; Olson, S.L.; Sorenson, M.D.; Jackson, J.; Fleischer, R.C. mtDNA from fossils reveals a radiation of Hawaiian geese recently derived from the Canada goose (Branta canadensis). Proc. Natl. Acad. Sci. USA 2002, 99, 1399–1404. [Google Scholar] [CrossRef]
- Bert, D.G. Determining the Spatial Scale of Species Response Habitat. Bioscience 2004, 54, 227–233. [Google Scholar] [CrossRef]
- Goloubinoff, P.; Pääbo, S.; Wilson, A.C. Evolution of maize inferred from sequence diversity of an Adh2 gene segment from archaeological specimens. Proc. Natl. Acad. Sci. USA 1993, 90, 1997–2001. [Google Scholar] [CrossRef]
- Jaenicke-Després, V.; Buckler, E.S.; Smith, B.D.; Gilbert, M.T.P.; Cooper, A.; Doebley, J.; Pääbo, S. Early allelic selection in maize as revealed by ancient DNA. Science 2003, 302, 1206–1208. [Google Scholar] [CrossRef]
- Poinar, H.; Kuch, M.; McDonald, G.; Martin, P.; Pääbo, S. Nuclear gene sequences from agyptiannetocene sloth coprolite. Curr. Biol. 2003, 13, 1150–1152. [Google Scholar] [CrossRef] [PubMed]
- Veeramah, K.R. Primate paleogenomics. Paleogenomics Genome-Scale Anal. Anc. DNA 2018, 353–373. [Google Scholar] [CrossRef]
- Kaestle, F.A.; Horsburgh, K.A. Ancient DNA in anthropology: Methods, applications, and ethics. Am. J. Phys. Anthr. 2002, 45, 92–130. [Google Scholar] [CrossRef]
- Karanth, K.P.; Delefosse, T.; Rakotosamimanana, B.; Parsons, T.J.; Yoder, A.D. Ancient DNA from giant extinct lemurs confirms single origin of Malagasy primates. Proc. Natl. Acad. Sci. USA 2005, 102, 5090–5095. [Google Scholar] [CrossRef]
- Stringer, C.B.; Andrews, P. Genetic and Fossil Evidence for the Origin of Modern Humans. Science 1988, 239, 1263–1268. [Google Scholar] [CrossRef]
- Ingman, M.; Kaessmann, H.; Pääbo, S.; Gyllensten, U. Mitochondrial genome variation and the origin of modern humans. Nature 2000, 408, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Maca-Meyer, N.; González, A.M.; Larruga, J.M.; Flores, C.; Cabrera, V.M. Major genomic mitochondrial lineages delineate early human expansions. BMC Genet. 2001, 2, 13. [Google Scholar] [CrossRef]
- Ovchinnikov, I.v.; Götherström, A.; Romanova, G.P.; Kharitonov, V.M.; Lidén, K.; Goodwin, W. Molecular analysis of Neanderthal DNA from the northern Caucasus. Nature 2000, 404, 490–493. [Google Scholar] [CrossRef]
- Serre, D.; Langaney, A.; Chech, M.; Teschler-Nicola, M.; Paunovic, M.; Mennecier, P.; Horfreiter, M.; Possnert, G.; Pääbo, S. No Evidence of Neandertal mtDNA Contribution to Early Modern Humans. PLoS Biol. 2006, 491–503. [Google Scholar] [CrossRef]
- Lum, J.K.; Cann, R.L. mtDNA lineage analyses: Origins and migrations of Micronesians and Polynesians. Am. J. Phys. Anthr. 2000, 113, 151–168. [Google Scholar]
- Redd, A.J.; Stoneking, M. Peopling of Sahul: mtDNA Variation in Aboriginal Australian and Papua New Guinean Populations. Am. J. Hum. Genet. 1999, 65, 808–828. [Google Scholar] [CrossRef] [PubMed]
- Silva, W.A.; Bonatto, S.L.; Holanda, A.J.; Ribeiro-Dos-Santos, A.K.; Paixão, B.M.; Goldman, G.H.; Abe-Sandes, K.; Rodriguez-Delfin, L.; Barbosa, M.; Paçó-Larson, M.L.; et al. Mitochondrial Genome Diversity of Native Americans Supports a Single Early Entry of Founder Populations into America. Am. J. Hum. Genet. 2002, 71, 187–192. [Google Scholar] [CrossRef] [PubMed]
- Simoni, L.; Calafell, F.; Pettener, D.; Bertranpetit, J.; Barbujani, G. Geographic Patterns of mtDNA Diversity in Europe. Am. J. Hum. Genet. 2000, 66, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Gretzinger, J.; Sayer, D.; Justeau, P.; Altena, E.; Pala, M.; Dulias, K.; Edwards, C.J.; Jodoin, S.; Lacher, L.; Sabin, S.; et al. The Anglo-Saxon migration and the formation of the early English gene pool. Nature 2022, 610, 112–119. [Google Scholar] [CrossRef]
- Narasimhan, V.M.; Patterson, N.; Moorjani, P.; Rohland, N.; Bernardos, R.; Mallick, S.; Lazaridis, I.; Nakatsuka, N.; Olalde, I.; Lipson, M.; et al. The formation of human populations in South and Central Asia. Science 2019, 365, eaat7487. [Google Scholar] [CrossRef]
- Allentoft, M.E.; Sikora, M.; Sjögren, K.; Rasmussen, S.; Rasmussen, M.; Stenderup, J.; Damgaard, P.B.; Schroeder, H.; Ahlström, T.; Vinner, L.; et al. Population genomics of Bronze Age Eurasia. Nature 2015, 522, 167–172. [Google Scholar] [CrossRef]
- Siska, V.; Jones, E.R.; Jeon, S.; Bhak, Y.; Kim, H.; Cho, Y.S.; Kim, H.; Lee, K.; Veselovskaya, E.; Balueva, T.; et al. Genome-wide data from two early Neolithic East Asian individuals dating to 7700 years ago. Sci. Adv. 2017, 3, e1601877. [Google Scholar] [CrossRef]
- Narasimhan, V.M.; Patterson, N.; Moorjani, P.; Lazaridis, I.; Lipson, M.; Mallick, S.; Rohland, N.; Bernardos, R.; Kim, A.M.; Nakatsuka, N.; et al. The Genomic Formation of South and Central Asia. bioRxiv 2018, 1–32. [Google Scholar] [CrossRef]
- Moorjani, P.; Thangaraj, K.; Patterson, N.; Lipson, M.; Loh, P.; Govindaraj, P.; Berger, B.; Reich, D.; Singh, L. Genetic Evidence for Recent Population Mixture in India. Am. J. Hum. Genet. 2013, 93, 422–438. [Google Scholar] [CrossRef]
- Reich, D.; Thangaraj, K.; Patterson, N.; Price, A.L.; Singh, L. Reconstructing Indian Population History. Nature 2009, 461, 489. [Google Scholar] [CrossRef]
- Shinde, V.; Narasimhan, V.M.; Rohland, N.; Mallick, S.; Mah, M.; Lipson, M.; Nakatsuka, N.; Adamski, N.; Broomandkhoshbacht, N.; Ferry, M.; et al. An Ancient Harappan Genome Lacks Ancestry from Steppe Pastoralists or Iranian Farmers. Cell 2019, 179, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Chaubey, G.; Metspalu, M.; Choi, Y.; Mägi, R.; Romero, I.G.; Soares, P.; Oven, M.; Behar, D.M.; Rootsi, S.; Hudjashov, G.; et al. Population Genetic Structure in Indian Austroasiatic Speakers: The Role of Landscape Barriers and Sex-Specific Admixture. Mol. Biol. Evol. 2011, 28, 1013–1024. [Google Scholar] [CrossRef] [PubMed]
- Harney, É.; Nayak, A.; Patterson, N.; Joglekar, P.; Mushrif-Tripathy, V.; Mallick, S.; Rohland, N.; Sedig, J.; Adamski, N.; Bernardos, R.; et al. Ancient DNA from the skeletons of Roopkund Lake reveals Mediterranean migrants in India. Nat. Commun. 2019, 10, 3670. [Google Scholar] [CrossRef] [PubMed]
- Sehrawat, J.S.; Agrawal, S.; Sankhyan, D.; Singh, M.; Kumar, S.; Prakash, S.; Rajpal, R.; Chaubey, G.; Thangaraj, K.; Rai, N. Pinpointing the Geographic Origin of 165-Year-Old Human Skeletal Remains Found in Punjab, India: Evidence from Mitochondrial DNA and Stable Isotope Analysis. Front. Genet. 2022, 13, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Prufer, K.; Racimo, F.; Patterson, N.; Jay, F.; Sankararaman, S.; Sawyer, S.; Heinze, A.; Renaud, G.; Sudmant, P.H.; de Filippo, C.; et al. The complete genome sequence of a Neanderthal from the Altai Mountains. Nature 2014, 505, 43–49. [Google Scholar] [CrossRef]
- Wall, J.D.; Yang, M.A.; Jay, F.; Kim, S.K.; Durand, E.Y.; Stevison, L.S.; Gignoux, C.; Woerner, A.; Hammer, M.F.; Slatkin, M. Higher levels of neanderthal ancestry in East Asians than in Europeans. Genetics 2013, 194, 199–209. [Google Scholar] [CrossRef]
- Vernot, B.; Akey, J.M. Complex history of admixture between modern humans and Neanderthals. Am. J. Hum. Genet. 2015, 94, 448–453. [Google Scholar] [CrossRef]
- Sanskararaman, S.; Mallick, S.; Dannemann, M.; Prüfer, K.; Kelso, J.; Pääbo, S.; Patterson, N.; Reich, D. The genomic landscape of Neanderthal ancestry in present-day humans. Nature 2014, 507, 354–357. [Google Scholar] [CrossRef]
- Sankararaman, S.; Patterson, N.; Li, H.; Pääbo, S.; Reich, D. The date of interbreeding between Neandertals and modern humans. PLoS Genet. 2012, 8, e1002947. [Google Scholar] [CrossRef]
- Vernot, B.; Akey, J.M. Resurrecting surviving Neandertal lineages from modern human genomes. Science 2014, 343, 1017–1021. [Google Scholar] [CrossRef]
- Kim, B.Y.; Lohmueller, K.E. Selection and reduced population size cannot explain higher amounts of Neandertal ancestry in East Asian that in European human population. Am. J. Hum. Genet. 2015, 96, 454–461. [Google Scholar] [CrossRef] [PubMed]
- Meyer, M.; Kircher, M.; Gansauge, M.; Li, H.; Racimo, F.; Mallick, S.; Schraiber, J.G.; Jay, F.; Prüfer, K.; Filippo, C.; et al. A high-coverage genome sequence from an archaic Denisovan individual. Science 2012, 338, 222–226. [Google Scholar] [CrossRef] [PubMed]
- Slatkin, M.; Racimo, F. Ancient DNA and human history. Proc. Natl. Acad. Sci. USA 2016, 113, 6380–6387. [Google Scholar] [CrossRef] [PubMed]
- Sawyer, S.; Renaud, G.; Viola, B.; Hublin, J.; Gansauge, M.; Shunkov, M.; Derevianko, A.P.; Prüfer, K.; Kelso, J.; Pääbo, S. Nuclear and mitochondrial DNA sequences from two Denisovan individuals. Proc. Natl. Acad. Sci. USA 2015, 112, 15696–15700. [Google Scholar] [CrossRef]
- Warinner, C.; Herbig, A.; Mann, A.; Yates, J.A.F.; Weiß, C.L.; Burbano, H.A.; Orlando, L.; Krause, J. A Robust Framework for Microbial Archaeology. Annu. Rev. Genom. Hum. Genet. 2017, 18, 321–356. [Google Scholar] [CrossRef]
- Mafart, B.; Kéfi, R.; Béraud-Colomb, E. Palaeopathological and palaeogenetic study of 13 cases of developmental dysplasia of the hip with dislocation in a historical population from southern France. Int. J. Osteoarchaeol. 2007, 17, 26–38. [Google Scholar] [CrossRef]
- Robbins, G.; Tripathy, V.M.; Misra, V.N.; Mohanty, R.K.; Shinde, V.S.; Gray, K.M.; Schug, M.D. Ancient Skeletal Evidence for Leprosy in India (2000 B.c.). PLoS ONE 2009, 4, e5669. [Google Scholar] [CrossRef]
- Salo, W.L.; Aufderheide, A.C.; Buikstra, J.; Holcomb, T.A. Identification of Mycobacterium tuberculosis DNA in a pre-Columbian Peruvian mummy. Proc. Natl. Acad. Sci. USA 1994, 91, 2091–2094. [Google Scholar] [CrossRef]
- Haas, C.J.; Zink, A.; Molnar, E.; Szeimies, U.; Reischl, U.; Marcsik, A. Molecular Evidence for Different Stages of Tuberculosis in Ancient Bone Samples from Hungary. Am. J. Phys. Anthropol. 2000, 113, 293–304. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1002/1096-8644(200011)113:3%3C293::AID-AJPA2%3E3.0.CO;2-6 (accessed on 4 April 2022). [CrossRef]
- Mays, S.; Taylor, G.M.; Legge, A.J.; Young, D.B.; Turner-Walker, G. Paleopathological and biomolecular study of tuberculosis in a medieval skeletal collection from England. Am. J. Phys. Anthropol. 2001, 114, 298–311. [Google Scholar] [CrossRef]
- Zink, A.; Haas, C.J.; Reischl, U.; Szeimies, U.; Nerlich, A.G. Molecular analysis of skeletal tuberculosis in an angyptianyptian population. J. Med. Microbiol. 2001, 50, 355–366. [Google Scholar] [CrossRef]
- Donoghue, H.D.; Spigelman, M.; Greenblatt, C.L.; Lev-Maor, G.; Bar-Gal, G.K.; Matheson, C.; Vernon, K.; Nerlich, A.G.; Zink, A.R. Tuberculosis: From prehistory to Robert Koch, as revealed by ancient DNA. Lancet Infect. Dis. 2004, 4, 584–592. [Google Scholar] [CrossRef]
- Drancourt, M.; Aboudharam, G.; Signoli, M.; Dutour, O.; Raoult, D. Detection of 400-year-old Yersinia pestis DNA in human dental pulp: An approach to the diagnosis of ancient septicemia. Proc. Natl. Acad. Sci. USA 1998, 95, 12637–12640. [Google Scholar] [CrossRef]
- Bos, K.I.; Schuenemann, V.J.; Golding, G.B.; Burbano, H.A.; Waglechner, N.; Coombes, B.K.; Mcphee, J.B.; DeWitte, S.N.; Meyer, M.; Schmedes, S.; et al. A draft genome of Yersinia pestis from victims of the Black Death. Nature 2011, 478, 506–510. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.M.; Klumk, J.; Harbeck, M.; Devault, D.; Waglechner, N.; Sahl, J.W.; Enk, J.; Birdsell, D.N.; Kuch, M.; Lumibao, C.; et al. Yersinia pestis and the Plague of Justinian 541–543 AD: A genomic analysis. Lancet Infect. Dis. 2014, 14, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Kim, Y.J.; Bisht, R.S.; Dangi, V.; Shirvalkar, P.; Jadhav, N.; Oh, C.S.; Hong, J.H.; Chai, J.Y.; Seo, M.; et al. Archaeoparasitological strategy based on the microscopic examinations of prehistoric samples and the recent report on the difference in the prevalence of soil transmitted helminthic infections in the Indian subcontinent. Anc. Asia 2018, 9, 1–6. [Google Scholar] [CrossRef]
- Pimenoff, V.N.; Houldcroft, C.J.; Rifkin, R.F.; Underdown, S. The Role of aDNA in Understanding the Coevolutionary Patterns of Human Sexually Transmitted Infections. Genes 2018, 9, 317. [Google Scholar] [CrossRef]
- Orlando, L.; Cooper, A. Using Ancient DNA to Understand Evolutionary and Ecological Processes. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 573–598. [Google Scholar] [CrossRef]
- Llamas, B.; Holland, M.L.; Chen, K.; Cropley, J.E.; Cooper, A.; Suter, C.M. High-Resolution Analysis of Cytosine Methylation in Ancient dNA. PLoS ONE 2012, 7, e30226. [Google Scholar] [CrossRef]
- Orlando, L.; Willerslev, E. An epigenetic window into the past? Science 2014, 345, 511–512. [Google Scholar] [CrossRef]
- Reich, D. Who We Are and How We Got Here Ancient DNA and the New Science of the Human Past; Oxford University Press: Oxford, UK, 2018. [Google Scholar]
- Gamba, C.; Hanghøj, K.; Gaunitz, C.; Alfarhan, A.H.; Alquraishi, S.A.; Al-Rasheid, K.A.; Bradley, D.G.; Orlando, L. Comparing the performance of three ancient DNA extraction methods for high-throughput sequencing. Mol. Ecol. Resour. 2016, 16, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Yang, D.Y.; Eng, B.; Waye, J.S.; Dudas, C.J.; Saunders, S.R. Improved ancient DNA purification for PCR using ion-exchange columns. Am. J. Phys. Anthropol. 1998, 136, 114–121. [Google Scholar] [CrossRef]
- Kistler, L. Ancient DNA extraction from plants. Methods Mol. Biol. 2012, 840, 71–79. [Google Scholar] [CrossRef] [PubMed]
- Warinner, C.; Rodrigues, J.F.M.; Vyas, R.; Trachsel, C.; Shved, N.; Grossmann, J.; Radini, A.; Hancock, Y.; Tito, R.Y.; Fiddyment, S.; et al. Pathogens and host immunity in the ancient human oral cavity. Nat. Genet. 2014, 46, 336–344. [Google Scholar] [CrossRef]
- Lech, T. Ancient DNA in historical parchments—Identifying a procedure for extraction and amplification of genetic material. Genet. Mol. Res. 2016, 15, 1–9. [Google Scholar] [CrossRef]
- Wood, J.R.; Wilmshurst, J.M. A protocol for subsampling Late Quaternary coprolites for multi-proxy analysis. Quat. Sci. Rev. 2016, 138, 1–5. [Google Scholar] [CrossRef]
- Green, E.J.; Speller, C.F. Novel Substrates as Sources of Ancient DNA: Prospects and Hurdles. Genes 2017, 8, 1–26. [Google Scholar] [CrossRef]
- Wandeler, P.; Hoeck, P.E.A.; Keller, L.F. Back to the future: Museum specimens in population genetics. Trends Ecol. Evol. 2007, 22, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C. Museums: The endangered dead. Nature 2015, 518, 292–294. [Google Scholar] [CrossRef]
- Pawłowska, J.; Lejzerowicz, F.; Esling, P.; Szczuciński, W.; Zajaogonekczkowski, M.; Pawlowski, J. Ancient DNA sheds new light on the Svalbard foraminiferal fossil record of the last millennium. Geobiology 2014, 12, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Fiddyment, S.; Holsinger, B.; Ruzzier, C.; Devine, A.; Binois, A.; Albarella, U.; Fischer, R.; Nichols, E.; Curtis, A.; Cheese, E.; et al. Animal origin of 13th-century uterine vellum revealed using noninvasive peptide fingerprinting. Proc. Natl. Acad. Sci. USA 2015, 112, 15066–15071. [Google Scholar] [CrossRef]
- Gomes, C.; Palomo-Díez, S.; Roig, J.; López-Parra, A.M.; Baeza-Richer, C.; Esparza-Arroyo, A.; Gibaja, J.; Arroyo-Pardo, E. Nondestructive extraction DNA method from bones or teeth, true or false? Forensic. Sci. Int. Genet. Suppl. Ser. 2015, 5, 279–282. [Google Scholar] [CrossRef]
- Andersen, J.C.; Mills, N.J. DNA extraction from museum specimens of parasitic Hymenoptera. PLoS ONE 2012, 7, e45549. [Google Scholar] [CrossRef] [PubMed]
- Grigorenko, A.P.; Borinskaya, S.A.; Yankovsky, N.K.; Rogaev, E.I. Achievements and Peculiarities in Studies of Ancient DNA and DNA from Complicated Forensic Specimens. Acta Nat. 2009, 1, 58–69. Available online: https://pmc/articles/PMC3347533/ (accessed on 27 June 2022). [CrossRef]
- Margulies, M.; Egholm, M.; Altman, W.E.; Attiya, S.; Bader, J.S.; Bemben, L.A.; Berka, J.; Braverman, M.S.; Chen, Y.J.; Chen, Z.; et al. Genome sequencing in microfabricated high-density picolitre reactors. Nature 2005, 437, 376–380. [Google Scholar] [CrossRef]
- Mamanova, L.; Coffey, A.J.; Scott, C.E.; Kozarewa, I.; Turner, E.H.; Kumar, A.; Howard, E.; Shendure, J.; Turner, D.J. Target-enrichment strategies for next-generation sequencing. Nat. Methods 2010, 7, 111–118. [Google Scholar] [CrossRef] [PubMed]
- Hofreiter, M.; Paijmans, J.L.; Goodchild, H.; Speller, C.F.; Barlow, A.; Fortes, G.G.; Thomas, J.A.; Ludwig, A.; Collins, M.J. The future of ancient DNA: Technical advances and conceptual shifts. BioEssays 2015, 37, 284–293. [Google Scholar] [CrossRef] [PubMed]
- Pickrell, J.K.; Reich, D. Toward a new history and geography of human genes informed by ancient DNA. Trends Genet. 2014, 30, 377–389. [Google Scholar] [CrossRef]
- Fu, Q.; Meyer, M.; Gao, X.; Stenzel, U.; Burbano, H.A.; Kelso, J.; Pääbo, S. DNA analysis of an early modern human from Tianyuan Cave, China. Proc. Natl. Acad. Sci. USA 2013, 110, 2223–2227. [Google Scholar] [CrossRef]
- der Sarkissian, C.; Balanovsky, O.; Brandt, G.; Khartanovich, V.; Buzhilova, A.; Koshel, S.; Zaporozhchenko, V.; Gronenborn, D.; Moiseyev, V.; Kolpakov, E.; et al. Ancient DNA Reveals Prehistoric Gene-Flow from Siberia in the Complex Human Population History of North East Europe. PLoS Genet. 2013, 9, e1003296. [Google Scholar] [CrossRef]
- Rivollat, M.; Mendisco, F.; Pemonge, M.H.; Safi, A.; Saint-Marc, D.; Brémond, A.; Couture-Veschambre, C.; Rottier, S.; Deguilloux, M.F. When the Waves of European Neolithization Met: First Paleogenetic Evidence from Early Farmers in the Southern Paris Basin. PLoS ONE 2015, 10, e0125521. [Google Scholar] [CrossRef]
- Cerqueira, C.C.S.; Paixão-Côrtes, V.R.; Zambra, F.M.B.; Salzano, F.M.; Hünemeier, T.; Bortolini, M.C. Predicting homo pigmentation phenotype through genomic data: From neanderthal to James Watson. Am. J. Hum. Biol. 2012, 24, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, P.; Storå, J.; Götherström, A.; Jakobsson, M. Accurate sex identification of ancient human remains using DNA shotgun sequencing. J. Archaeol. Sci. 2013, 40, 4477–4482. [Google Scholar] [CrossRef]
- Morozova, I.; Flegontov, P.; Mikheyev, A.S.; Bruskin, S.; Asgharian, H.; Ponomarenko, P.; Klyuchnikov, V.; ArunKumar, G.; Prokhortchouk, E.; Gankin, Y.; et al. Toward high-resolution population genomics using archaeological samples. DNA Res. 2016, 23, 295–310. [Google Scholar] [CrossRef]
- McVean, G. A Genealogical Interpretation of Principal Components Analysis. PLoS Genet. 2009, 5, e1000686. [Google Scholar] [CrossRef]
- Patterson, N.; Price, A.L.; Reich, D. Population Structure and Eigenanalysis. PLoS Genet. 2006, 2, e190. [Google Scholar] [CrossRef]
- Maixner, F.; Krause-Kyora, B.; Turaev, D.; Herbig, A.; Hoopmann, M.R.; Hallows, J.L.; Kusebauch, U.; Vigl, E.E.; Malfertheiner, P.; Megraud, F.; et al. The 5300-year-old Helicobacter pylori genome of the Iceman. Science 2016, 351, 162–165. [Google Scholar] [CrossRef] [PubMed]
- Ávila-Arcos, M.C.; Castro, C.d.; Nieves-Colón, M.A.; Raghavan, M. Recommendations for Sustainable Ancient DNA Research in the Global South: Voices from a New Generation of Paleogenomicists. Front. Genet. 2022, 13, 880170. [Google Scholar] [CrossRef] [PubMed]
- Dados, N.; Connell, R. The global south. Contexts 2012, 11, 12–13. [Google Scholar] [CrossRef]
- Minasny, B.; Fiantis, D. “Helicopter Research”: Who Benefits from International Studies in Indonesia? Available online: https://theconversation.com/helicopter-research-who-benefits-from-international-studies-in-indonesia-102165 (accessed on 18 August 2022).
- Haelewaters, D.; Hofmann, T.A.; Romero-Olivares, A.L. Ten simple rules for Global North researchers to stop perpetuating helicopter research in the Global South. PLoS Comput. Biol. 2021, 17, e1009277. [Google Scholar] [CrossRef]
- Thangaraj, K.; Rai, N. Peopling of India: Ancient DNA perspectives. J. Biosci. 2019, 44, 70. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.S.; Nasidze, I.; Walimbe, S.R.; Stoneking, M. Brief communication: Discouraging prospects for ancient DNA from India. Am. J. Phys. Anthropol. 2000, 113, 129–133. [Google Scholar] [CrossRef] [PubMed]
- Argüelles, J.M.; Fuentes, A.; Yáñez, B. Analyzing asymmetries and praxis in aDNA research: A bioanthropological critique. Am Anthropol. 2022, 124, 130–140. [Google Scholar] [CrossRef]
- Lindahl, T. Instability and decay of the primary structure of DNA. Nature 1993, 362, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Smith, C.I.; Chamberlain, A.T.; Riley, M.S.; Stringer, C.; Collins, M.J. The thermal history of human fossils and the likelihood of successful DNA amplification. J. Hum. Evol. 2003, 45, 203–217. [Google Scholar] [CrossRef]
- Molak, M.; Ho, S.Y.W. Evaluating the impact of post-mortem damage in ancient DNA: A theoretical approach. J. Mol. Evol. 2011, 73, 244–255. [Google Scholar] [CrossRef]
- Dabney, J.; Meyer, M.; Pääbo, S. Ancient DNA Damage. Cold Spring Harb. Perspect. Biol. 2013, 5, a012567. [Google Scholar] [CrossRef]
- Paijmans, J.L.A.; Gilbert, M.T.P.; Hofreiter, M. Mitogenomic analyses from ancient DNA. Mol. Phylogenet. Evol. 2013, 69, 404–416. [Google Scholar] [CrossRef]
- Bollongino, R.; Tresset, A.; Vigne, J.D. Environment and excavation: Pre-lab impacts on ancient DNA analyses. C. R. Palevol. 2008, 7, 91–98. [Google Scholar] [CrossRef]
- Green, R.E.; Briggs, A.W.; Krause, J.; Prüfer, K.; Burbano, H.A.; Siebauer, M.; Lachmann, M.; Pääbo, S. The Neandertal genome and ancient DNA authenticity. EMBO J. 2009, 28, 2494–2502. [Google Scholar] [CrossRef]
- Renaud, G.; Slon, V.; Duggan, A.T.; Kelso, J. Schmutzi: Estimation of contamination and endogenous mitochondrial consensus calling for ancient DNA. Genome Biol. 2015, 16, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Skoglund, P.; Northoff, B.H.; Shunkov, M.V.; Derevianko, A.P.; Pääbo, S.; Krause, J.; Jakobsson, M. Separating endogenous ancient DNA from modern day contamination in a Siberian Neandertal. Proc. Natl. Acad. Sci. USA 2014, 111, 2229–2234. [Google Scholar] [CrossRef] [PubMed]
- Renaud, G.; Hanghøj, K.; Willerslev, E.; Orlando, L. gargammel: A sequence simulator for ancient DNA. Bioinformatics 2017, 33, 577–579. [Google Scholar] [CrossRef] [PubMed]
- Heidt, A. Ancient DNA Boom Underlines a Need for Ethical Frameworks. Science 2022, 1–10. Available online: https://www.the-scientist.com/news-opinion/ancient-dna-boom-underlines-a-need-for-ethical-frameworks-69645 (accessed on 18 August 2022).
- Scheib, C.L.; Li, H.; Desai, T.; Link, V.; Kendall, C.; Dewar, G.; Griffith, P.W.; Mörseburg, A.; Johnson, J.R.; Potter, A.; et al. Ancient human parallel lineages within North America contributed to a coastal expansion. Science 2018, 360, 1024–1027. [Google Scholar] [CrossRef]
- Nakatsuka, N.; Lazaridis, I.; Barbieri, C.; Skoglund, P.; Rohland, N.; Mallick, S.; Posth, C.; Harkins-Kinkaid, K.; Ferry, M.; Harney, É.; et al. A Paleogenomic Reconstruction of the Deep Population History of the Andes. Cell 2020, 181, 1131–1145.e21. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.K.; Colwell, C.; Claw, K.G.; Stone, A.C.; Bolnick, D.A.; Hawks, J.; Brothers, K.B.; Nanibaa’A, G. Fostering Responsible Research on Ancient DNA. Am. J. Hum. Genet. 2020, 107, 183–195. [Google Scholar] [CrossRef]
- Alpaslan-Roodenberg, S.; Anthony, D.; Babiker, H.; Bánffy, E.; Booth, T.; Capone, P.; Deshpande-Mukherjee, A.; Eisenmann, S.; Fehren-Schmitz, L.; Frachetti, M.; et al. Ethics of DNA research on human remains: Five globally applicable guidelines. Nature 2021, 599, 41–46. [Google Scholar] [CrossRef]
- Cooper, A.; Poinar, H.N. Ancient DNA: Do it right or not at all. Science 2000, 289, 1139. [Google Scholar] [CrossRef]
- Chaubey, G.; Ayub, Q.; Rai, N.; Prakash, S.; Mushrif-Tripathy, V.; Mezzavilla, M.; Pathak, A.K.; Tamang, R.; Firasat, S.; Reidla, M.; et al. “Like sugar in milk”: Reconstructing the genetic history of the Parsi population. Genome Biol. 2017, 18, 1–9. [Google Scholar] [CrossRef]
- Pasupuleti, N.; Mustak, M.S. Ancient DNA: Revisiting past through population genomics. Polymorphism 2020, 5, 45–52. Available online: https://www.peerpublishers.com/index.php/snp/article/view/62 (accessed on 18 August 2022).
- Thangaraj, K.; Chaubey, G.; Kivisild, T.; Reddy, A.G.; Singh, V.K.; Rasalkar, A.A.; Singh, L. Reconstructing the origin of Andaman Islanders. Science 2005, 308, 996. [Google Scholar] [CrossRef] [PubMed]
- Tamang, R.; Thangaraj, K. Genomic view on the peopling of India. Investig. Genet. 2012, 3, 1–9. [Google Scholar] [CrossRef]
- Tamang, R.; Singh, L.; Thangaraj, K. Complex genetic origin of Indian populations and its implications. J. Biosci. 2012, 37, 911–919. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dalal, V.; Pasupuleti, N.; Chaubey, G.; Rai, N.; Shinde, V. Advancements and Challenges in Ancient DNA Research: Bridging the Global North–South Divide. Genes 2023, 14, 479. https://doi.org/10.3390/genes14020479
Dalal V, Pasupuleti N, Chaubey G, Rai N, Shinde V. Advancements and Challenges in Ancient DNA Research: Bridging the Global North–South Divide. Genes. 2023; 14(2):479. https://doi.org/10.3390/genes14020479
Chicago/Turabian StyleDalal, Vasundhra, Nagarjuna Pasupuleti, Gyaneshwer Chaubey, Niraj Rai, and Vasant Shinde. 2023. "Advancements and Challenges in Ancient DNA Research: Bridging the Global North–South Divide" Genes 14, no. 2: 479. https://doi.org/10.3390/genes14020479
APA StyleDalal, V., Pasupuleti, N., Chaubey, G., Rai, N., & Shinde, V. (2023). Advancements and Challenges in Ancient DNA Research: Bridging the Global North–South Divide. Genes, 14(2), 479. https://doi.org/10.3390/genes14020479