
Global Frequency Analyses of Canine Progressive Rod-Cone Degeneration–Progressive Retinal Atrophy and Collie Eye Anomaly Using Commercial Genetic Testing Data
 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Samples
2.2. Statistical Analyses
3. Results
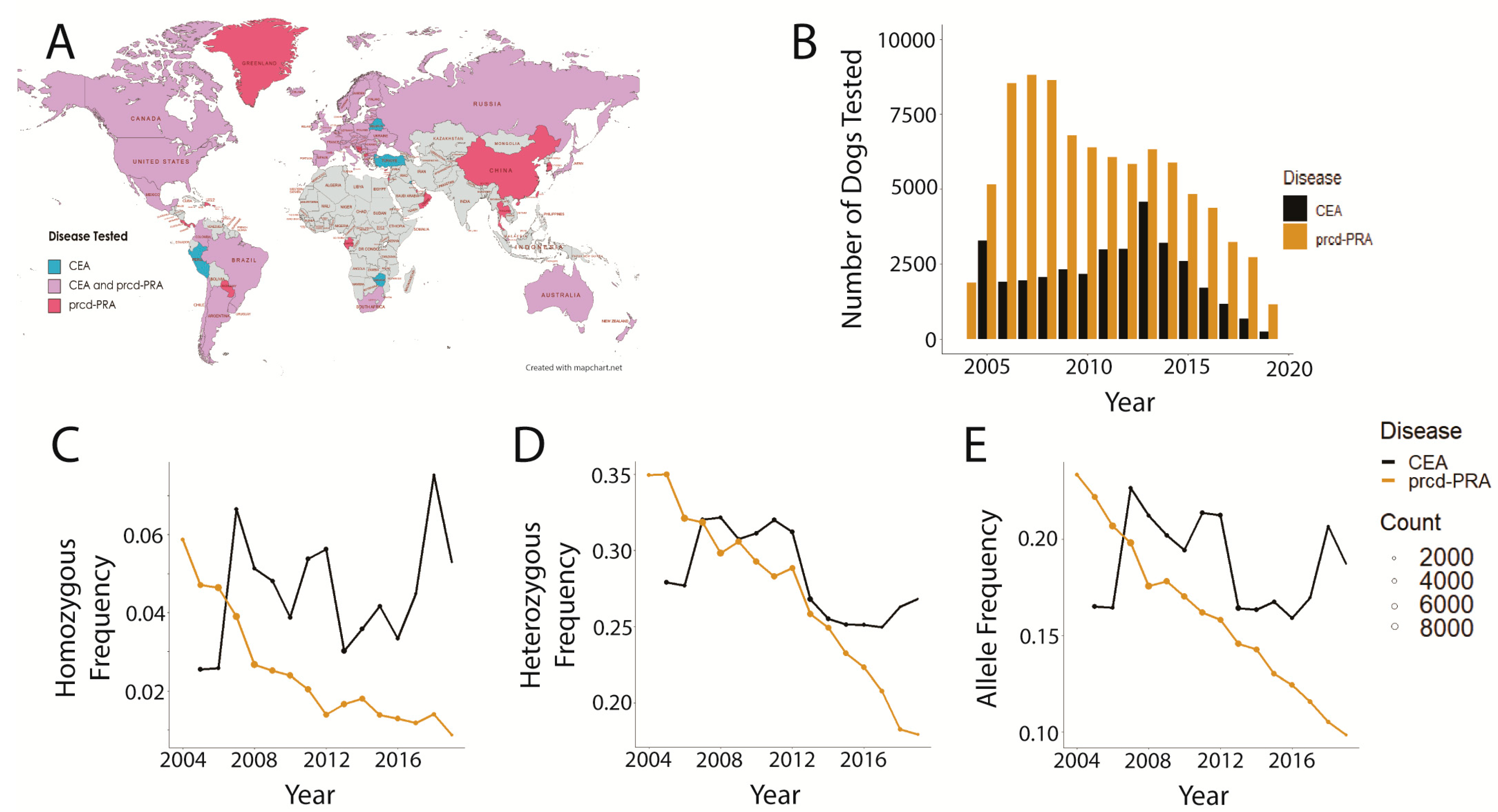
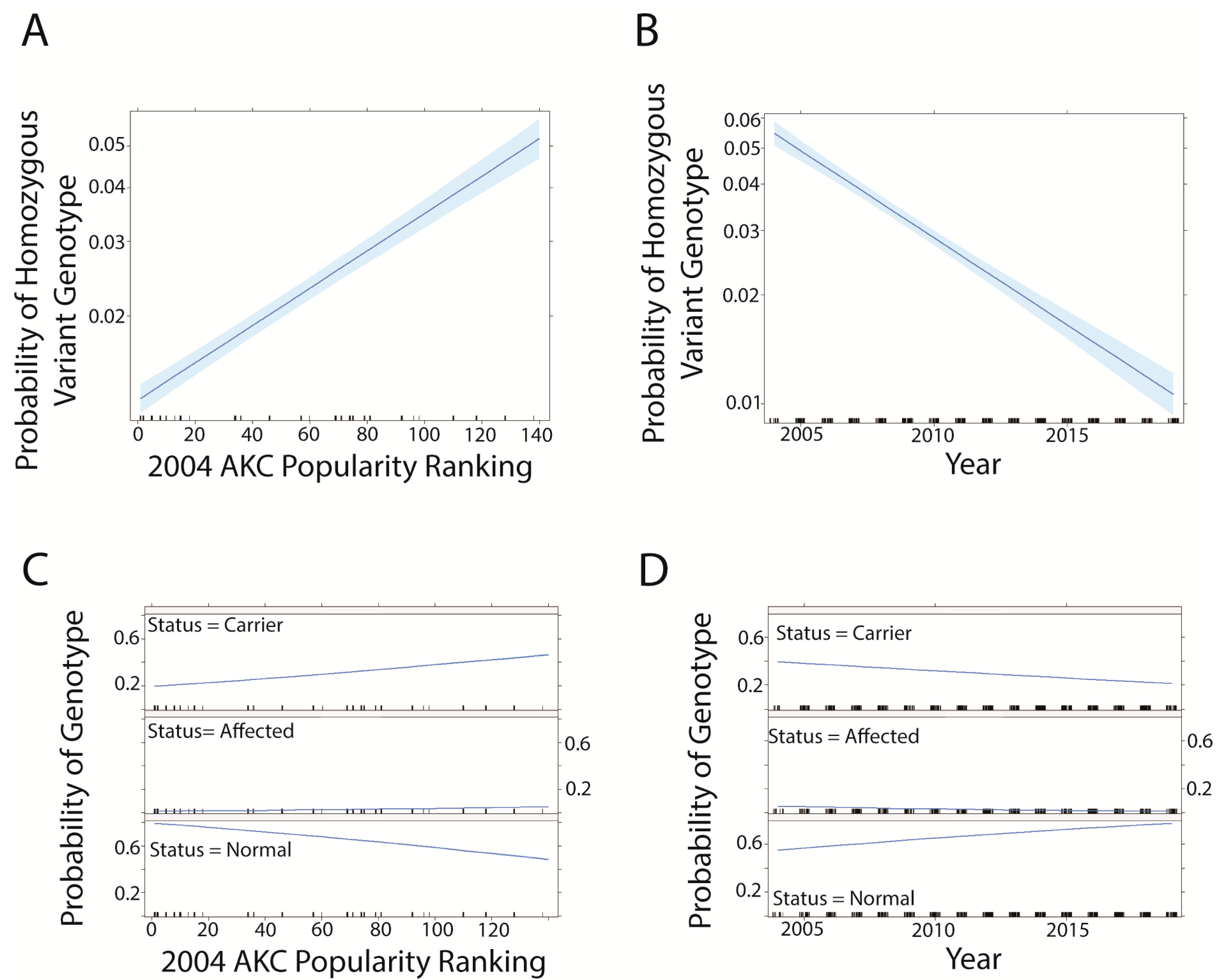
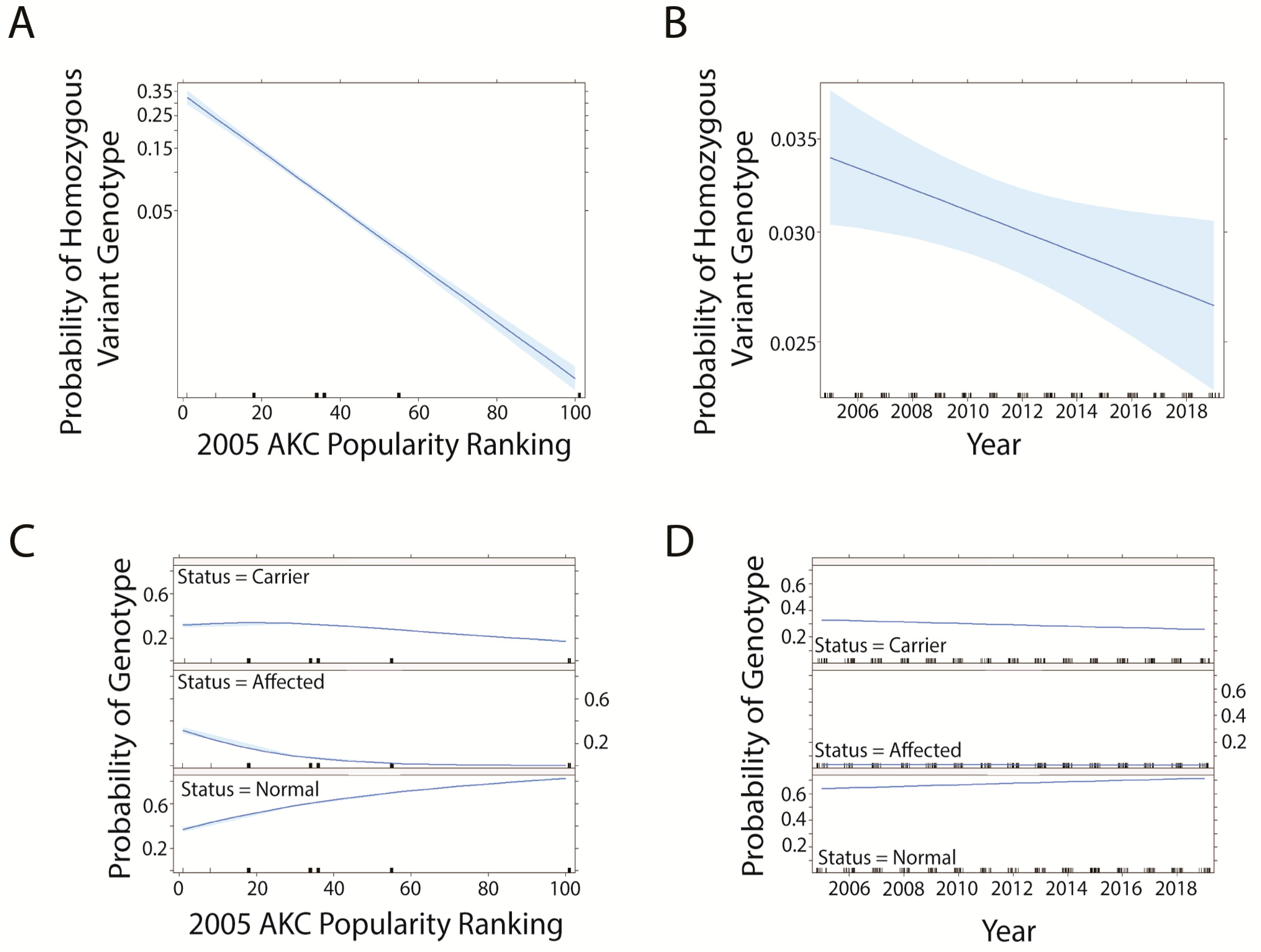
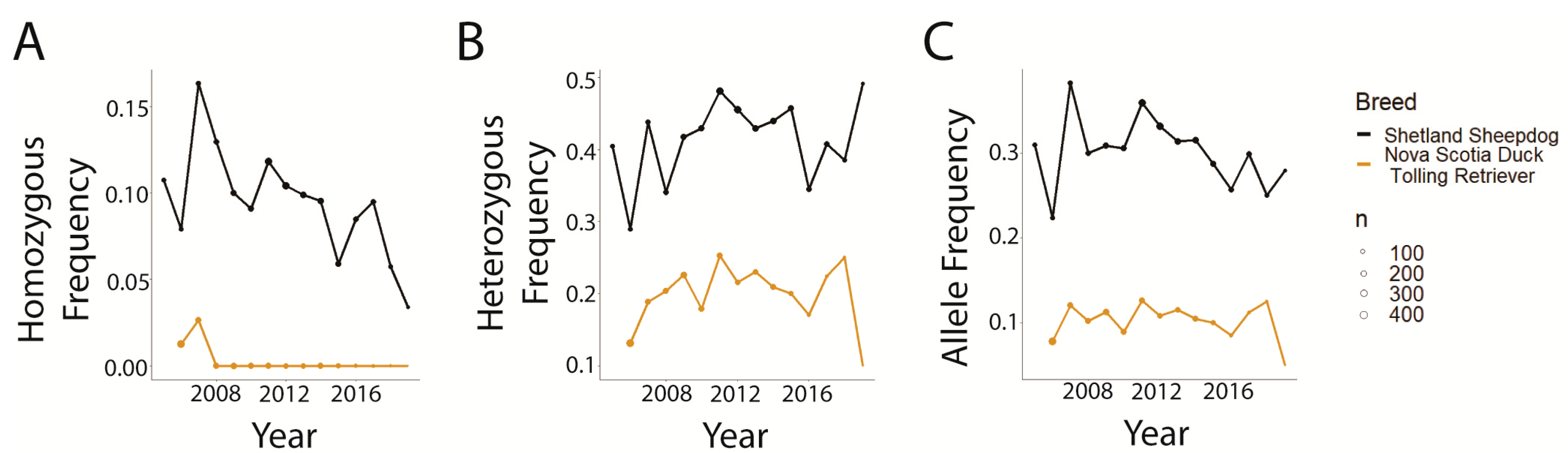
3.1. Data Distributions and Disease Comparisons
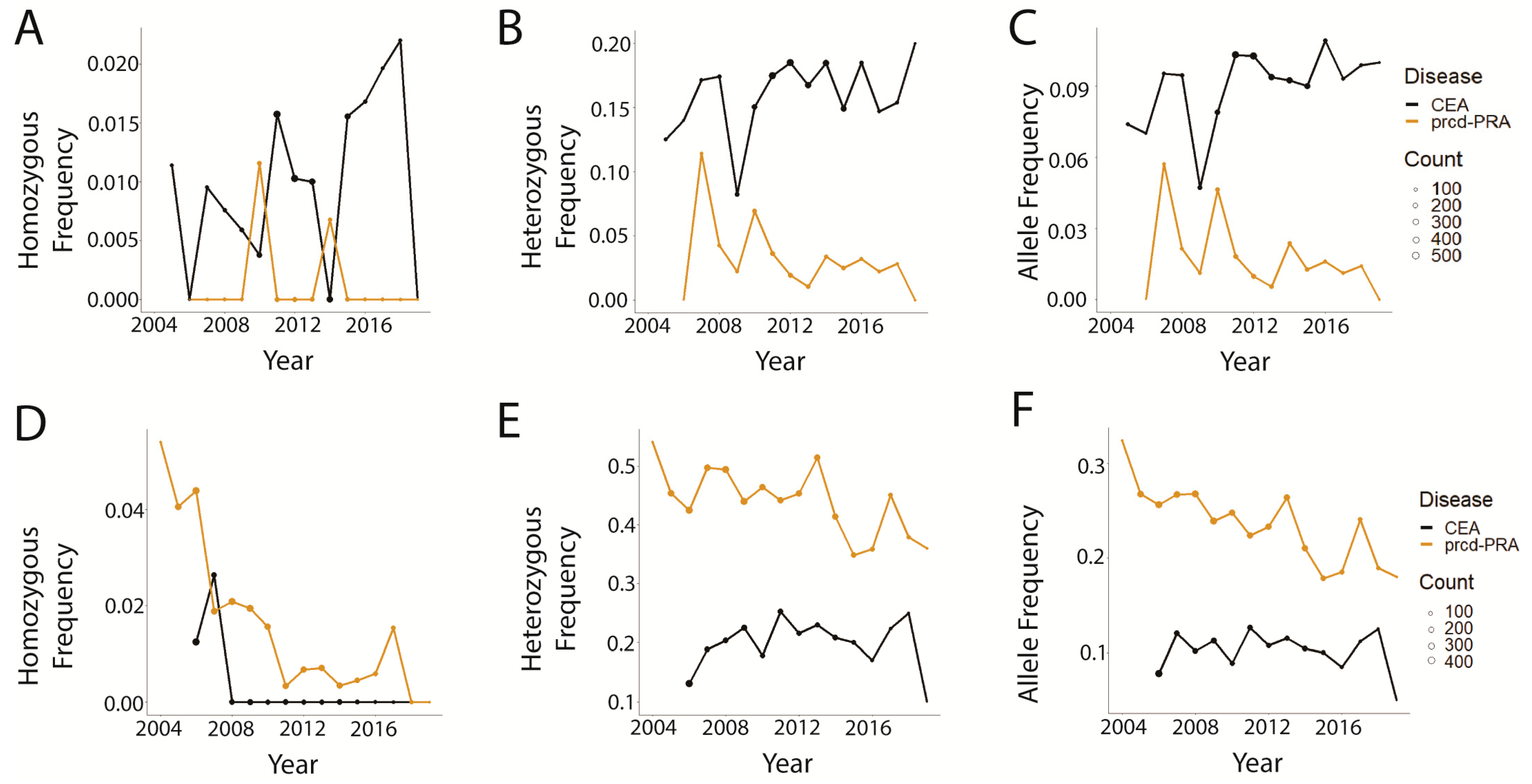
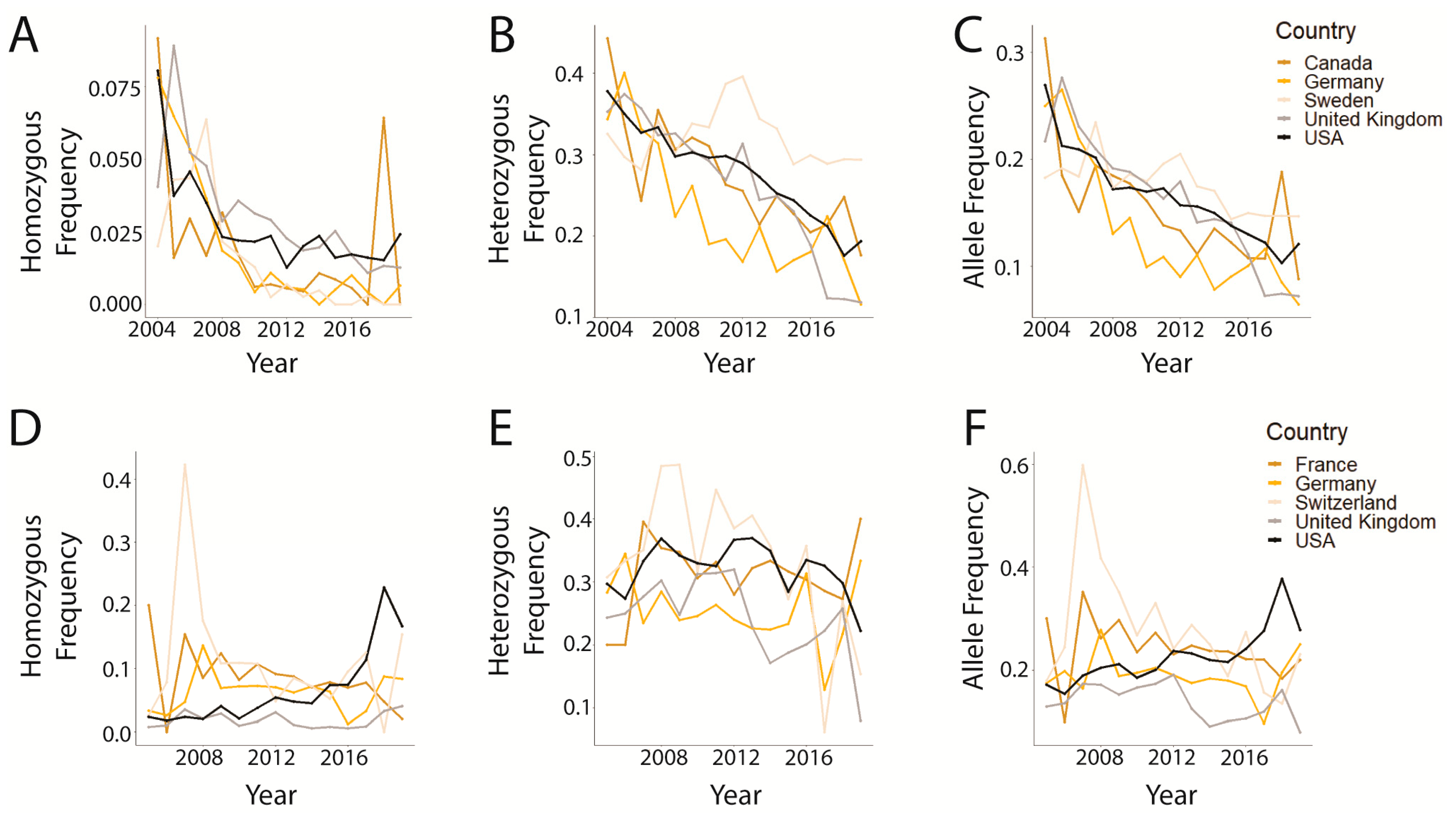
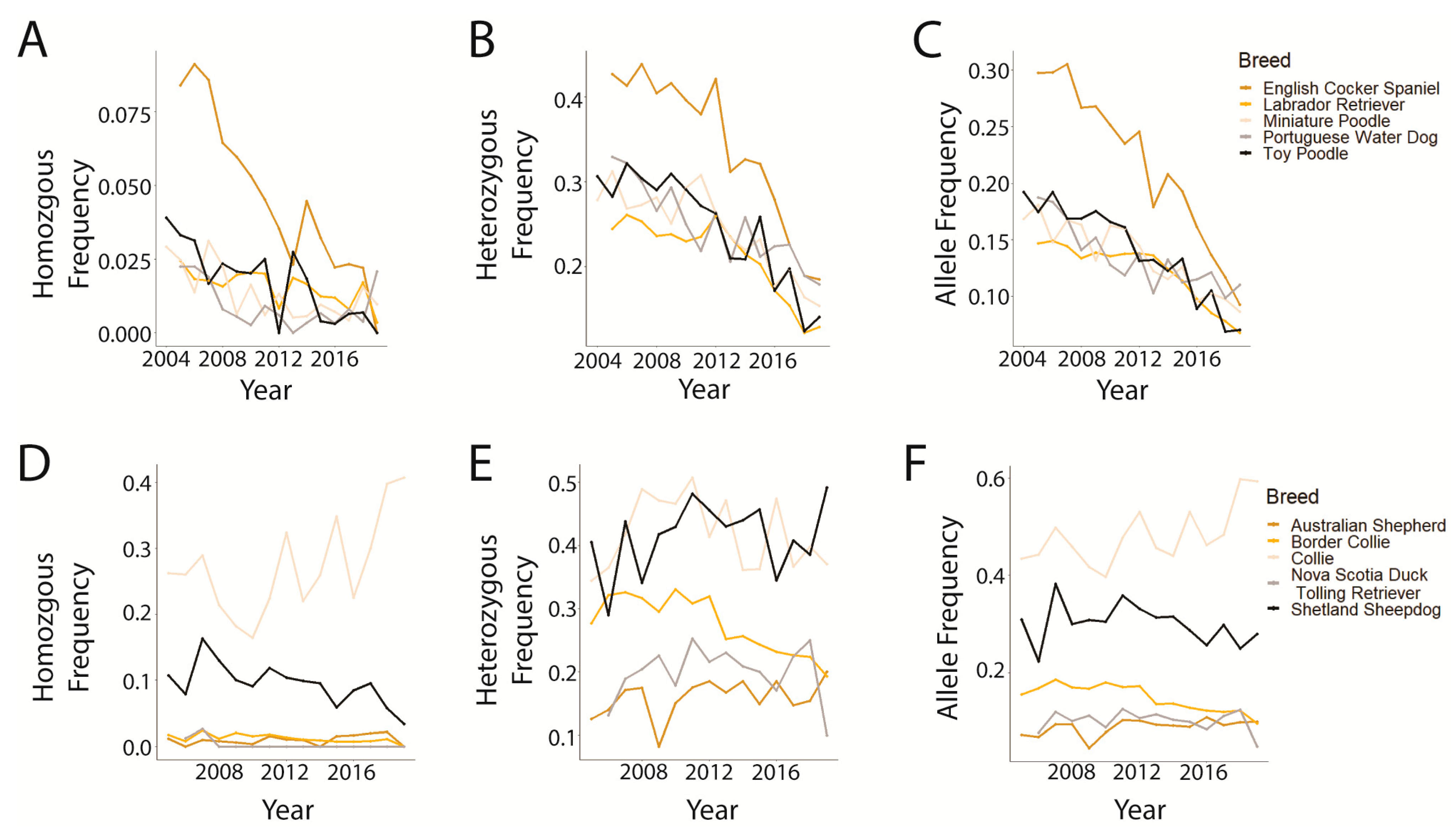
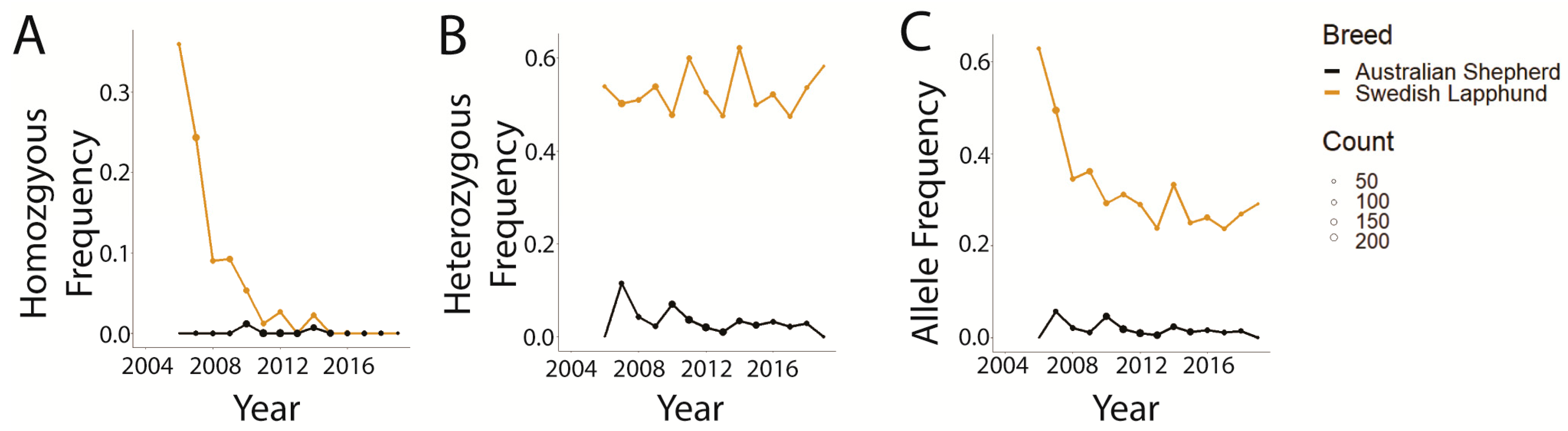
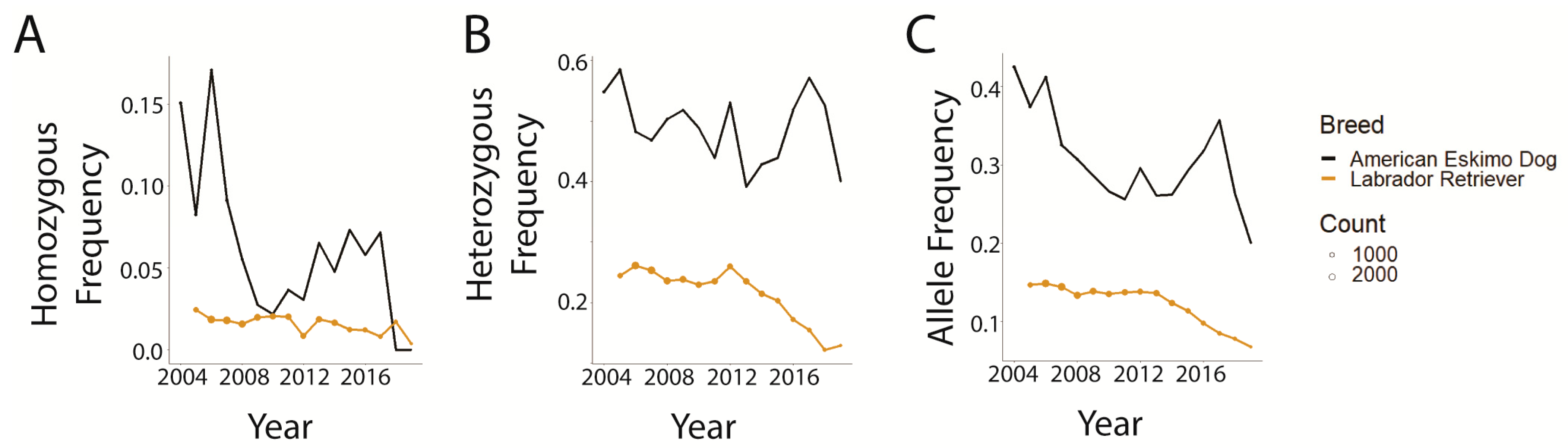
3.2. Trends in prcd-PRA
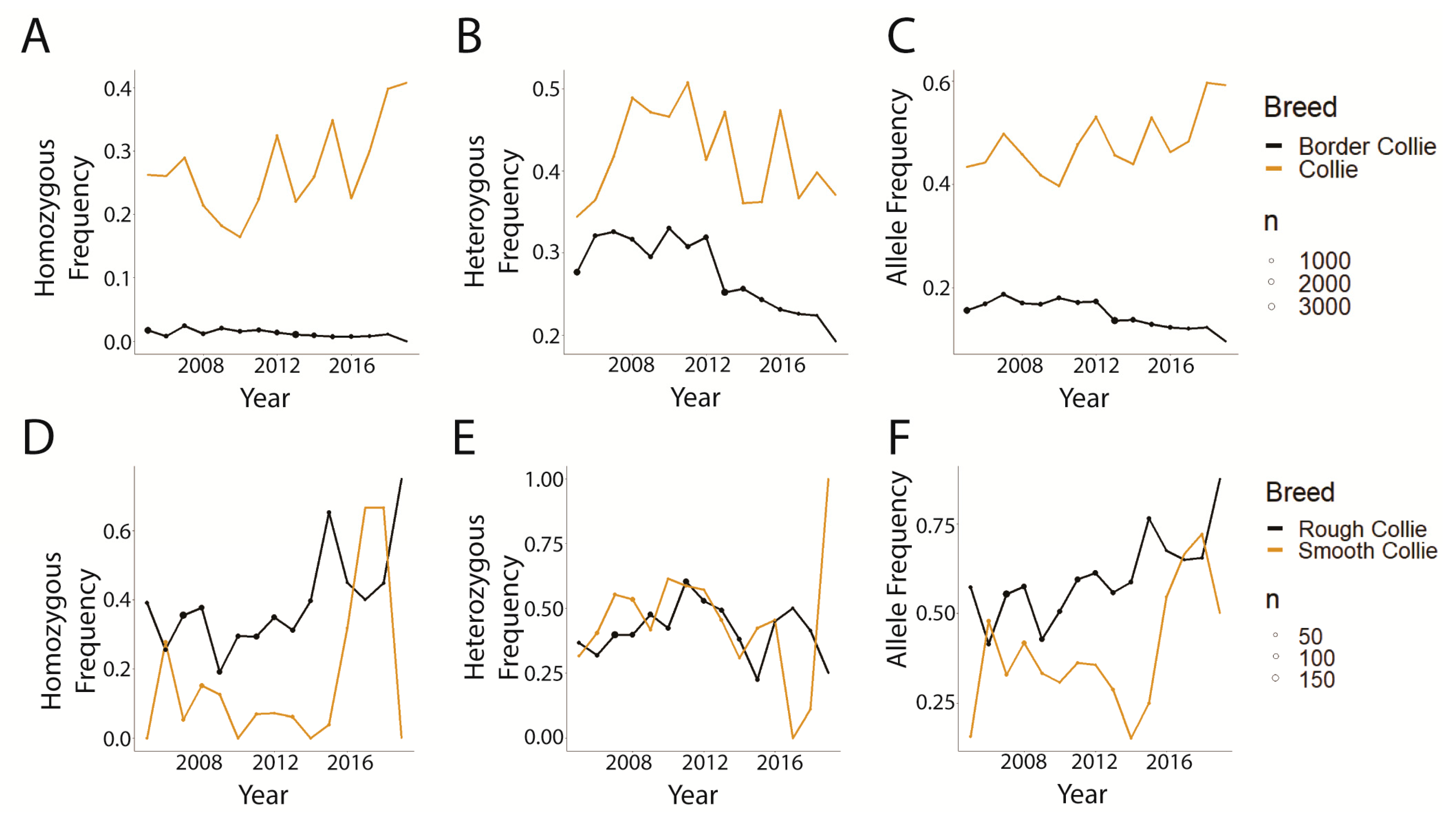
3.3. Trends in CEA
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Online Mendelian Inheritance in Animals (OMIA). Faculty of Veterinary Science, University of Sydney. Available online: http://omia.angis.org.au/ (accessed on 19 May 2022).
- Nicholas, F.W. Online Mendelian Inheritance in Animals (OMIA): A comparative knowledgebase of genetic disorders and other familial traits in non-laboratory animals. Nucleic Acids Res. 2003, 31, 275–277. [Google Scholar] [CrossRef]
- Bunel, M.; Chaudieu, G.; Hamel, C.; Lagoutte, L.; Manes, G.; Botherel, N.; Brabet, P.; Pilorge, P.; André, C.; Quignon, P. Natural models for retinitis pigmentosa: Progressive retinal atrophy in dog breeds. Hum. Genet. 2019, 138, 441–453. [Google Scholar] [CrossRef]
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef] [PubMed]
- Loukovitis, E.; Anastasia, S.; Tranos, P.; Balidis, M.; Asteriadis, S.; Thanos, V.; Thanos, S.; Anogeianakis, G. A review of recent developments in retinitis pigmentosa genetics, its clinical features, and natural course. Med. Hypothesis Discov. Innov. Ophthalmol. 2021, 9, 231–254. [Google Scholar] [CrossRef]
- Zangerl, B.; Goldstein, O.; Philp, A.R.; Lindauer, S.J.P.; Pearce-Kelling, S.E.; Mullins, R.F.; Graphodatsky, A.S.; Ripoll, D.; Felix, J.S.; Stone, E.M.; et al. Identical mutation in a novel retinal gene causes progressive rod-cone degeneration in dogs and retinitis pigmentosa in humans. Genomics 2006, 88, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Allon, G.; Mann, I.; Remez, L.; Sehn, E.; Rizel, L.; Nevet, M.J.; Perlman, I.; Wolfrum, U.; Ben-Yosef, T. PRCD is concentrated at the base of photoreceptor outer segments and is involved in outer segment disc formation. Hum. Mol. Genet. 2019, 28, 4078–4088. [Google Scholar] [CrossRef] [PubMed]
- Spencer, W.J.; Ding, J.D.; Lewis, T.R.; Yu, C.; Phan, S.; Pearring, J.N.; Kim, K.Y.; Thor, A.; Mathew, R.; Kalnitsky, J.; et al. PRCD is essential for high-fidelity photoreceptor disc formation. Proc. Natl. Acad. Sci. USA 2019, 116, 13087–13096. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, G.; Alligood, J.; O’Brien, P.; Buyukmihci, N. Pathogenesis of progressive rod-cone degeneration in miniature poodles. Investig. Ophthalmol. Vis. Sci. 1982, 23, 610–630. [Google Scholar]
- Dostal, J.; Hrdlicova, A.; Horak, P. Progressive rod-cone degeneration (PRCD) in selected dog breeds and variability in its phenotypic expression. Vet. Med. 2011, 56, 243–247. [Google Scholar] [CrossRef]
- Downs, L.M.; Hitti, R.; Pregnolato, S.; Mellersh, C.S. Genetic screening for PRA-associated mutations in multiple dog breeds shows that PRA is heterogeneous within and between breeds. Vet. Ophthalmol. 2014, 17, 126–130. [Google Scholar] [CrossRef]
- Donner, J.; Freyer, J.; Davison, S.; Anderson, H.; Blades, M.; Honkanen, L.; Inman, L.; Brookhart-Knox, C.A.; Louviere, A.; Forman, O.P.; et al. Genetic prevalence and clinical relevance of canine Mendelian disease variants in over one million dogs. PLoS Genet. 2023, 19, e1010651. [Google Scholar] [CrossRef] [PubMed]
- Donner, J.; Anderson, H.; Davison, S.; Hughes, A.M.; Bouirmane, J.; Lindqvist, J.; Lytle, K.M.; Ganesan, B.; Ottka, C.; Ruotanen, P.; et al. Frequency and distribution of 152 genetic disease variants in over 100,000 mixed breed and purebred dogs. PLoS Genet. 2018, 14, e1007361. [Google Scholar] [CrossRef] [PubMed]
- Kohyama, M.; Tada, N.; Mitsui, H.; Tomioka, H.; Tsutsui, T.; Yabuki, A.; Rahman, M.M.; Kushida, K.; Mizukami, K.; Yamato, O. Real-time PCR genotyping assay for canine progressive rod-cone degeneration and mutant allele frequency in Toy Poodles, Chihuahuas and Miniature Dachshunds in Japan. J. Vet. Med. Sci. 2016, 78, 481–484. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Miyadera, K.; Acland, G.M.; Aguirre, G.D. Genetic and phenotypic variations of inherited retinal diseases in dogs: The power of within- and across-breed studies. Mamm. Genome 2012, 23, 40–61. [Google Scholar] [CrossRef] [PubMed]
- Paw Print Genetics Progressive Retinal Atrophy, Progressive Rod-Cone Degeneration. Available online: https://www.pawprintgenetics.com/products/tests/details/89/ (accessed on 7 October 2023).
- All DNA Tests—OFA. Available online: https://ofa.org/all-dna-tests/?TEST=PROGRESSIVERETINALATROPHY (accessed on 13 February 2023).
- Lenffer, J. OMIA (Online Mendelian Inheritance in Animals): An enhanced platform and integration into the Entrez search interface at NCBI. Nucleic Acids Res. 2006, 34, D599–D601. [Google Scholar] [CrossRef]
- UCDavis. Progressive Rod-Cone Degeneration (PRA-prcd). Available online: https://vgl.ucdavis.edu/test/pra-prcd#:~:text=toearlyadulthood.[MostPRCD-affecteddogshavenoticeablevisualimpairmentby4,typicallyprogressingtocompleteblindness] (accessed on 7 October 2023).
- WisdomPanel, Mars Petcare Science & Diagnostics MyBreedData. Available online: https://mybreeddata.com/ (accessed on 7 October 2023).
- Barnett, K.C.; Stades, F.C. Collie eye anomaly in the Shetland Sheepdog in The Netherlands. J. Small Anim. Pract. 1979, 20, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Barnett, K.C. Collie eye anomaly (CEA). J. Small Anim. Pract. 1979, 20, 537–542. [Google Scholar] [CrossRef]
- Palanova, A. Collie eye anomaly: A review. Vet. Med. 2016, 60, 345–350. [Google Scholar] [CrossRef]
- Fredholm, M.; Larsen, R.C.; Jönsson, M.; Söderlund, M.A.; Hardon, T.; Proschowsky, H.F. Discrepancy in compliance between the clinical and genetic diagnosis of choroidal hypoplasia in Danish Rough Collies and Shetland Sheepdogs. Anim. Genet. 2016, 47, 250–252. [Google Scholar] [CrossRef]
- Lowe, J.K.; Kukekova, A.V.; Kirkness, E.F.; Langlois, M.C.; Aguirre, G.D.; Acland, G.M.; Ostrander, E.A. Linkage mapping of the primary disease locus for collie eye anomaly. Genomics 2003, 82, 86–95. [Google Scholar] [CrossRef]
- Sargan, D.R. Collie eye anomaly in the rough collie. J. Small Anim. Pract. 2001, 42, 204. [Google Scholar] [CrossRef][Green Version]
- Yakely, W.L.; Wyman, M.; Donovan, E.F.; Fechheimer, N.S. Genetic Transmission of an Ocular Fundus Anomaly in Collies. J. Am. Vet. Med. Assoc. 1968, 152, 457–461. [Google Scholar] [PubMed]
- Yakely, W.L. Collie Eye Anomaly: Decreased Prevalence Through Selective Breeding. J. Am. Vet. Med. Assoc. 1972, 161, 1103–1107. [Google Scholar] [PubMed]
- Parker, H.G.; Kukekova, A.V.; Akey, D.T.; Goldstein, O.; Kirkness, E.F.; Baysac, K.C.; Mosher, D.S.; Aguirre, G.D.; Acland, G.M.; Ostrander, E.A. Breed relationships facilitate fine-mapping studies: A 7.8-kb deletion cosegregates with Collie eye anomaly across multiple dog breeds. Genome Res. 2007, 17, 1562–1571. [Google Scholar] [CrossRef] [PubMed]
- Pagon, R.A. Ocular coloboma. Surv. Ophthalmol. 1981, 25, 223–236. [Google Scholar] [CrossRef] [PubMed]
- Woodbine, L.; Gennery, A.R.; Jeggo, P.A. The clinical impact of deficiency in DNA non-homologous end-joining. DNA Repair 2014, 16, 84–96. [Google Scholar] [CrossRef] [PubMed]
- Buck, D.; Malivert, L.; de Chasseval, R.; Barraud, A.; Fondanèche, M.-C.; Sanal, O.; Plebani, A.; Stéphan, J.-L.; Hufnagel, M.; le Deist, F.; et al. Cernunnos, a Novel Nonhomologous End-Joining Factor, Is Mutated in Human Immunodeficiency with Microcephaly. Cell 2006, 124, 287–299. [Google Scholar] [CrossRef]
- Thorsrud, J.A.; Huson, H.J. Description of breed ancestry and genetic health traits in arctic sled dog breeds. Canine Med. Genet. 2021, 8, 8. [Google Scholar] [CrossRef]
- Dostál, J.; Horák, P.; Hrdlicová, A.; Stratil, A. Simplified PCR analysis of a mutation in the NHEJ1 gene causing collie eye anomaly in some dog breeds. Czech J. Anim. Sci. 2010, 55, 346–350. [Google Scholar] [CrossRef]
- Mizukami, K.; Chang, H.-S.; Ota, M.; Yabuki, A.; Hossain, M.A.; Rahman, M.M.; Uddin, M.M.; Yamato, O. Collie eye anomaly in Hokkaido dogs: Case study. Vet. Ophthalmol. 2012, 15, 128–132. [Google Scholar] [CrossRef]
- Paw Print Genetics Collie Eye Anomaly. Available online: https://www.pawprintgenetics.com/products/tests/details/92/ (accessed on 7 October 2023).
- The Health of the Auvergne Shepherd—Chien Berger d’Auvergne (ASCBA). Available online: https://www.chienbergerdauvergne.org/la-sante/ (accessed on 22 February 2023).
- Brown, E.A.; Thomasy, S.M.; Murphy, C.J.; Bannasch, D.L. Genetic analysis of optic nerve head coloboma in the Nova Scotia Duck Tolling Retriever identifies discordance with the NHEJ1 intronic deletion (collie eye anomaly mutation). Vet. Ophthalmol. 2018, 21, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Wallin-Håkanson, B.; Wallin-Håkanson, N.; Hedhammar, Å. Collie eye anomaly in the rough collie in Sweden: Genetic transmission and influence on offspring vitality. J. Small Anim. Pract. 2000, 41, 254–258. [Google Scholar] [CrossRef] [PubMed]
- Sigdel, A.; Niggel, J.; Aguirre, G.D.; Murgiano, L. Whole-genome mapping unravels the genetic etiology associated with complex phenotypes of collie eye anomaly. Investig. Ophthalmol. Vis. Sci. 2023, 64, 2779. [Google Scholar]
- Marelli, S.P.; Rizzi, R.; Paganelli, A.; Bagardi, M.; Minozzi, G.; Brambilla, P.G.; Polli, M. Genotypic and allelic frequency of a mutation in the NHEJ1 gene associated with collie eye anomaly in dogs in Italy. Vet. Rec. Open 2022, 9, e26. [Google Scholar] [CrossRef] [PubMed]
- Grosås, S.; Lingaas, F.; Prestrud, K.W.; Ropstad, E.-O. Compliance between clinical and genetic diagnosis of choroidal hypoplasia in 103 Norwegian Border Collie puppies. Vet. Ophthalmol. 2018, 21, 371–375. [Google Scholar] [CrossRef] [PubMed]
- Mizukami, K.; Yabuki, A.; Kohyama, M.; Kushida, K.; Rahman, M.M.M.; Uddin, M.M.M.; Sawa, M.; Yamato, O. Molecular prevalence of multiple genetic disorders in Border collies in Japan and recommendations for genetic counselling. Vet. J. 2016, 214, 21–23. [Google Scholar] [CrossRef]
- Trecenti-Santana, A.S.; Guiraldelli, G.G.; Albertino, L.G.; Ferreira, J.F.; Andrade, F.M.; Borges, A.S.; Oliveira-Filho, J.P. Allele frequency of SLC4A3 (PRA1), TTC8 (PRA2), and PRA-prcd mutations in golden retrievers in Brazil. Front. Vet. Sci. 2022, 9, 973854. [Google Scholar] [CrossRef] [PubMed]
- Andrade, L.R.; Caceres, A.M.; Trecenti, A.S.; Brandão, C.V.S.; Gandolfi, M.G.; Aguiar, E.V.; Andrade, D.G.A.; Borges, A.S.; Oliveira-Filho, J.P. Allele Frequency of the C.5G>A Mutation in the PRCD Gene Responsible for Progressive Retinal Atrophy in English Cocker Spaniel Dogs. Animals 2019, 9, 844. [Google Scholar] [CrossRef]
- Lewis, T.W.; Mellersh, C.S. Changes in mutation frequency of eight Mendelian inherited disorders in eight pedigree dog populations following introduction of a commercial DNA test. PLoS ONE 2019, 14, e0209864. [Google Scholar] [CrossRef]
- Takanosu, M. Different allelic frequency of progressive rod-cone degeneration in two populations of Labrador Retrievers in Japan. J. Vet. Med. Sci. 2017, 79, 1746–1748. [Google Scholar] [CrossRef]
- Talenti, A.; Dreger, D.L.; Frattini, S.; Polli, M.; Marelli, S.; Harris, A.C.; Liotta, L.; Cocco, R.; Hogan, A.N.; Bigi, D.; et al. Studies of modern Italian dog populations reveal multiple patterns for domestic breed evolution. Ecol. Evol. 2018, 8, 2911–2925. [Google Scholar] [CrossRef]
- Parker, H.G.; Dreger, D.L.; Rimbault, M.; Davis, B.W.; Mullen, A.B.; Carpintero-Ramirez, G.; Ostrander, E.A. Genomic Analyses Reveal the Influence of Geographic Origin, Migration, and Hybridization on Modern Dog Breed Development. Cell Rep. 2017, 19, 697–708. [Google Scholar] [CrossRef]
- Kesselheim, A.S.; Cook-Deegan, R.M.; Winickoff, D.E.; Mello, M.M. Gene Patenting—The Supreme Court Finally Speaks. N. Engl. J. Med. 2013, 369, 869–875. [Google Scholar] [CrossRef]
- Genetic Veterinary Sciences, Inc. V. Canine EIC Genetics, LLC, 101 F. Supp. 3d 833 | Casetext Search + Citator. Available online: https://casetext.com/case/genetic-veterinary-scis-inc-v-canine-eic-genetics-llc (accessed on 15 February 2023).
- Dziuk, E. The Canine Health Information Center (CHIC). In Proceedings of the Tufts’ Canine and Feline Breeding and Genetics Conference, Sturbridge, MA, USA, 30 September–1 October 2005. [Google Scholar]
- OFA. Browse By Breed. Available online: https://ofa.org/chic-programs/browse-by-breed/ (accessed on 14 December 2022).
- Shan, S.; Xu, F.; Brenig, B. Genome-Wide Association Studies Reveal Neurological Genes for Dog Herding, Predation, Temperament, and Trainability Traits. Front. Vet. Sci. 2021, 8, 693290. [Google Scholar] [CrossRef]
- Jones, P.; Chase, K.; Martin, A.; Davern, P.; Ostrander, E.A.; Lark, K.G. Single-Nucleotide-Polymorphism-Based Association Mapping of Dog Stereotypes. Genetics 2008, 179, 1033–1044. [Google Scholar] [CrossRef]
- Campbell, C.L.; Bhérer, C.; Morrow, B.E.; Boyko, A.R.; Auton, A. A Pedigree-Based Map of Recombination in the Domestic Dog Genome. G3 Genes Genomes Genet. 2016, 6, 3517–3524. [Google Scholar] [CrossRef] [PubMed]
- Bell, J. Genetic Testing and Counseling: A Trojan Horse for Dog and Cat Breeds?—TUFTSBG2007—VIN. In Proceedings of the Tufts’ Canine and Feline Breeding and Genetics Conference, Sturbridge, MA, USA, 13–14 September 2007. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| X2 | p-Value | |
|---|---|---|
| prcd-PRA | 698.0194 | 2.7 × 10−152 |
| CEA | 7.52339 | 0.023244 |
| LRM for Affected Genotype (p-Value) | MRM for Genotype (SE) | MRM for Carrier Genotype (SE) | |
|---|---|---|---|
| prcd-PRA | −1.252 × 10−1 (<2 × 10−16) | −0.04864 (1.07 × 10−5) | −0.06331 (5.54 × 10−6) |
| CEA | −2.46 × 10−2 (0.00834) | −0.03438 (1.62 × 10−5) | −0.01948 (9.95 × 10−6) |
| Rank | LRM for Affected Genotype (p-Value *) | MRM for Affected Genotype (SE) | MRM for Carrier Genotype (SE) |
|---|---|---|---|
| 1 | Swedish Lapphund (4.65 × 10−5) | Swedish Lapphund (7.031 × 10−4) | Swedish Lapphund (4.300 × 10−3) |
| 2 | American Eskimo Dog (0.921564) | Entlebucher Mountain Dog (2.957 × 10−4) | American Eskimo Dog (3.057 × 10−5) |
| 3 | Entlebucher Mountain Dog (0.97598) | American Eskimo Dog (1.597 × 10−6) | Entlebucher Mountain Dog (1.865 × 10−3) |
| 4 | Australian Cattle Dog (0.955735) | Australian Cattle Dog (1.073 × 10−3) | Nova Scotia Duck Tolling Retriever (1.626 × 10−2) |
| 5 | English Cocker Spaniel (0.006659) | English Cocker Spaniel (2.299 × 10−3) | Australian Cattle Dog (6.342 × 10−3) |
| 6 | Spanish Water Dog (0.000362) | Nova Scotia Duck Tolling Retriever (6.288 × 10−4) | English Cocker Spaniel (2.005 × 10−2) |
| 7 | Finnish Lapphund (4.75 × 10−10) | Spanish Water Dog (2.990 × 10−5) | Labradoodle, Australian (1.363 × 10−3) |
| 8 | Nova Scotia Duck Tolling Retriever (<2 × 10−16) | Finnish Lapphund (7.442 × 10−5) | Dwarf Poodle (8.613 × 10−4) |
| 9 | Toy Poodle (<2 × 10−16) | Toy Poodle (1.058 × 10−3) | Finnish Lapphund (1.702 × 10−3) |
| 10 | Chesapeake Bay Retriever (4.31 × 10−8) | Chesapeake Bay Retriever (2.371 × 10−5) | Chesapeake Bay Retriever (6.065 × 10−4) |
| 11 | Labrador Retriever (<2 × 10−16) | Labrador Retriever (6.113 × 10−4) | Spanish Water Dog (6.533 × 10−4) |
| 12 | Miniature Poodle (<2 × 10−16) | Miniature Poodle (7.277 × 10−4) | Portuguese Water Dog (8.761 × 10−3) |
| 13 | Dwarf Poodle (3.16 × 10−6) | Dwarf Poodle (1.075 × 10−5) | Toy Poodle (2.562 × 10−2) |
| 14 | Portuguese Water Dog (<2 × 10−16) | Portuguese Water Dog (1.544 × 10−4) | Miniature Poodle (2.593 × 10−2) |
| 15 | Chinese Crested (<2 × 10−16) | Labradoodle, Australian (2.312 × 10−5) | Labrador Retriever (1.638 × 10−2) |
| 16 | Labradoodle, Australian (4.71 × 10−13) | Chinese Crested (1.439 × 10−5) | Golden Retriever (1.280 × 10−3) |
| 17 | Golden Retriever (<2 × 10−16) | Australian Shepherd (4.887 × 10−6) | Chinese Crested (5.514 × 10−4) |
| 18 | Australian Shepherd (1.64 × 10−9) | Golden Retriever (7.927 × 10−6) | Australian Shepherd (2.257 × 10−4) |
| Rank | LRM for Affected Genotype (p-Value *) | MRM for Affected Genotype (SE) | MRM for Carrier Genotype (SE) |
|---|---|---|---|
| 1 | Collie (<2 × 10−16) | Collie (1.633 × 10−2) | Collie (2.571 × 10−2) |
| 2 | Shetland Sheepdog (<2 × 10−16) | Shetland Sheepdog (8.510 × 10−3) | Shetland Sheepdog (2.396 × 10−2) |
| 3 | Border Collie (0.00067) | Border Collie (8.850 × 10−3) | Border Collie (2.293 × 10−2) |
| 4 | Australian Shepherd (0.96233) | Australian Shepherd (5.506 × 10−6) | Nova Scotia Duck Tolling Retriever (3.209 × 10−2) |
| 5 | Nova Scotia Duck Tolling Retriever (0.38093) | Nova Scotia Duck Tolling Retriever (3.634 × 10−4) | Australian Shepherd (3.693 × 10−5) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Clark, J.A.; Anderson, H.; Donner, J.; Pearce-Kelling, S.; Ekenstedt, K.J. Global Frequency Analyses of Canine Progressive Rod-Cone Degeneration–Progressive Retinal Atrophy and Collie Eye Anomaly Using Commercial Genetic Testing Data. Genes 2023, 14, 2093. https://doi.org/10.3390/genes14112093
Clark JA, Anderson H, Donner J, Pearce-Kelling S, Ekenstedt KJ. Global Frequency Analyses of Canine Progressive Rod-Cone Degeneration–Progressive Retinal Atrophy and Collie Eye Anomaly Using Commercial Genetic Testing Data. Genes. 2023; 14(11):2093. https://doi.org/10.3390/genes14112093
Chicago/Turabian StyleClark, Jessica A., Heidi Anderson, Jonas Donner, Susan Pearce-Kelling, and Kari J. Ekenstedt. 2023. "Global Frequency Analyses of Canine Progressive Rod-Cone Degeneration–Progressive Retinal Atrophy and Collie Eye Anomaly Using Commercial Genetic Testing Data" Genes 14, no. 11: 2093. https://doi.org/10.3390/genes14112093
APA StyleClark, J. A., Anderson, H., Donner, J., Pearce-Kelling, S., & Ekenstedt, K. J. (2023). Global Frequency Analyses of Canine Progressive Rod-Cone Degeneration–Progressive Retinal Atrophy and Collie Eye Anomaly Using Commercial Genetic Testing Data. Genes, 14(11), 2093. https://doi.org/10.3390/genes14112093