

FOS Inhibits the Differentiation of Intramuscular Adipocytes in Goats
 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. Construction of Overexpression Vector, siRNA Synthesis, Cell Transfection
2.3. Oil Red O Staining
2.4. Bodipy Staining and DAPI Staining
2.5. Cell RNA Extraction and Quantitative Real-Time PCR
2.6. Measurement of Cellular Triglyceride Levels
2.7. Western Blotting
2.8. Statistical Analysis
3. Results
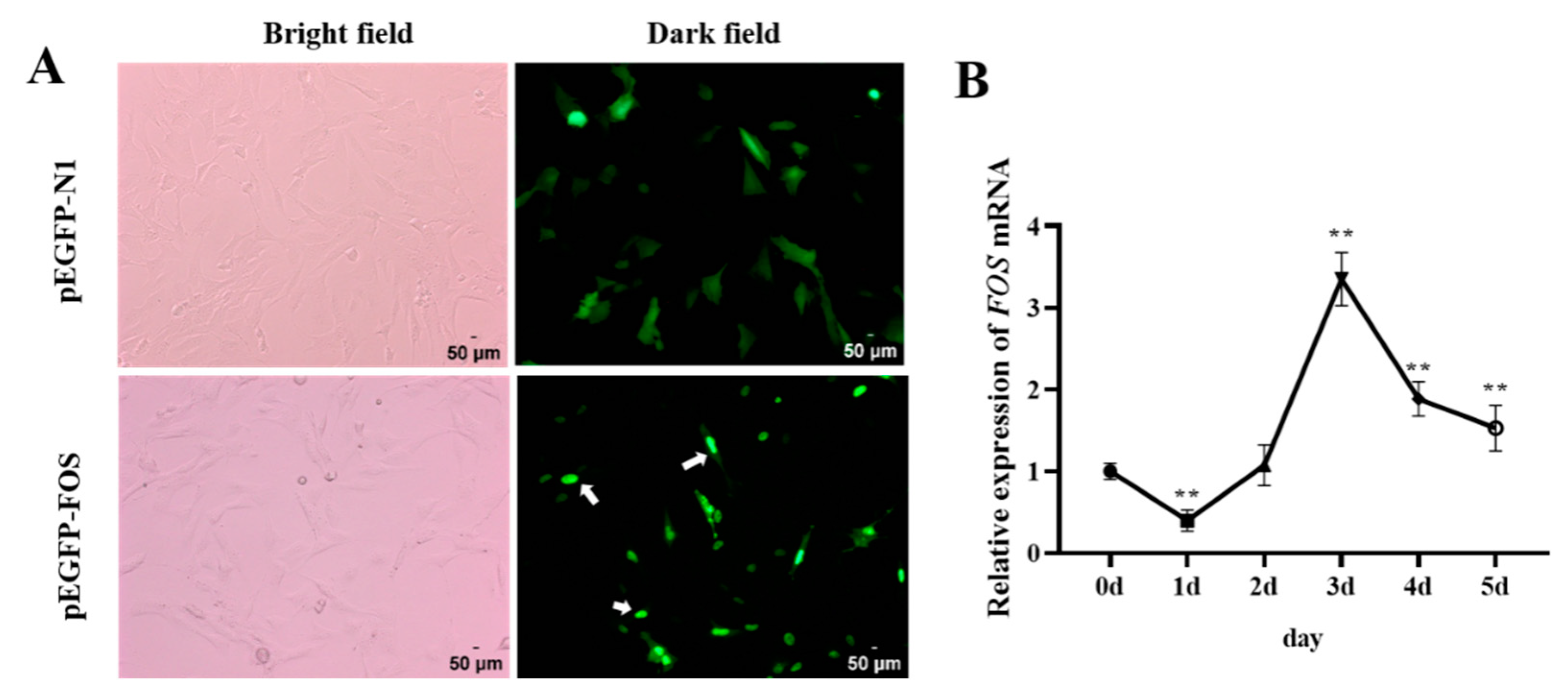
3.1. Subcellular Localization and Expression Patterns of FOS during Differentiation of Goat Intramuscular Adipocytes
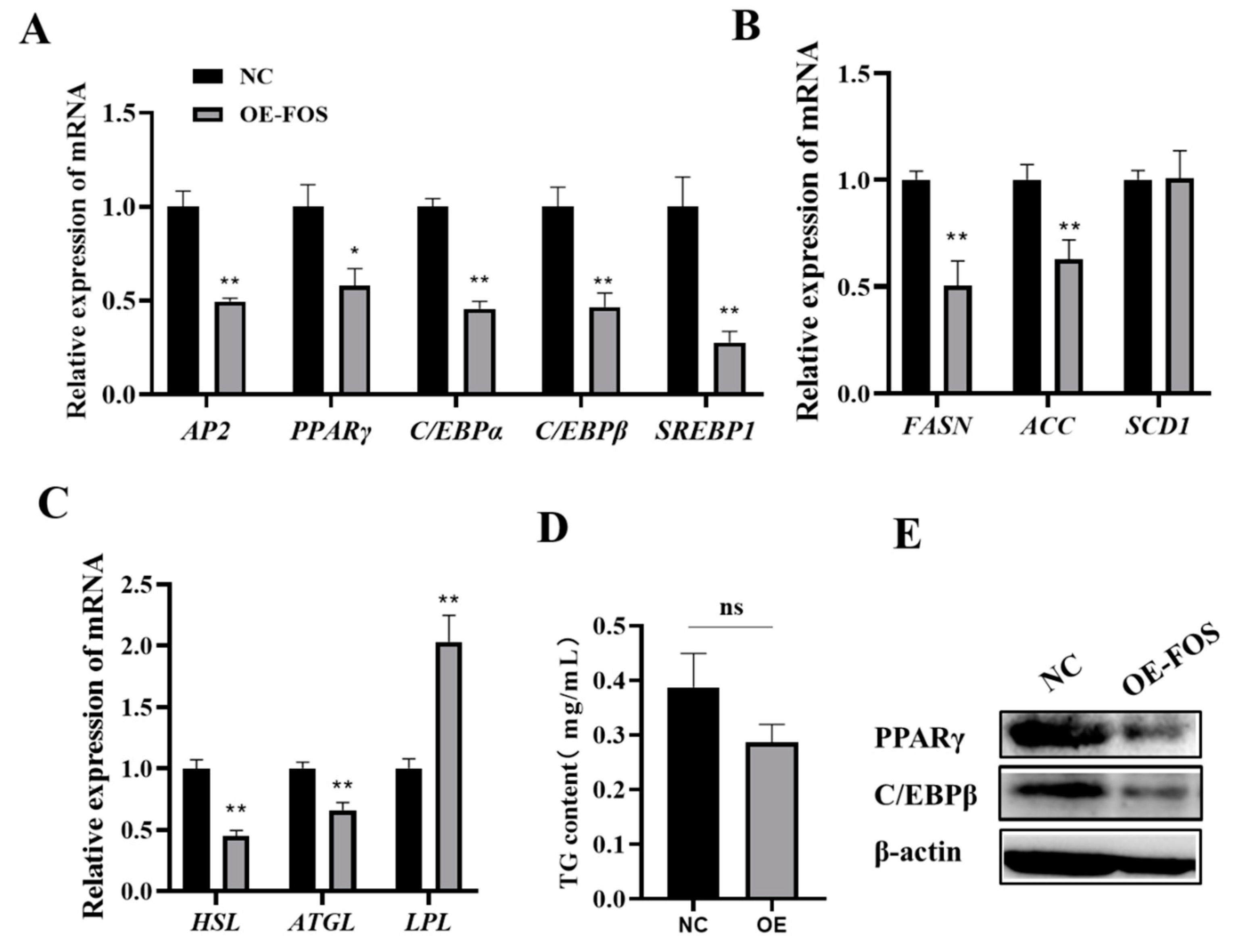
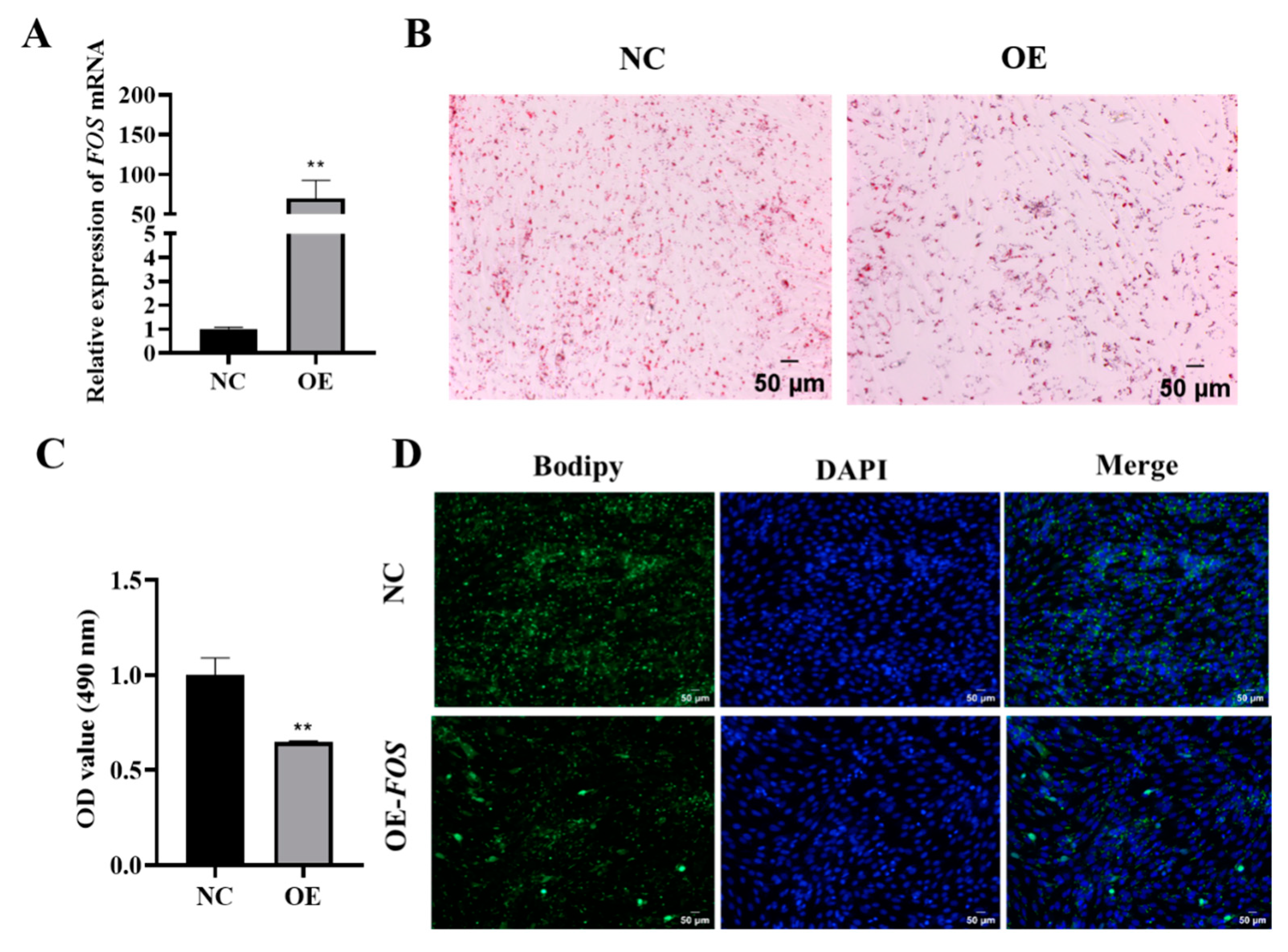
3.2. Overexpression of FOS Inhibits Goat Intramuscular Preadipocyte Differentiation
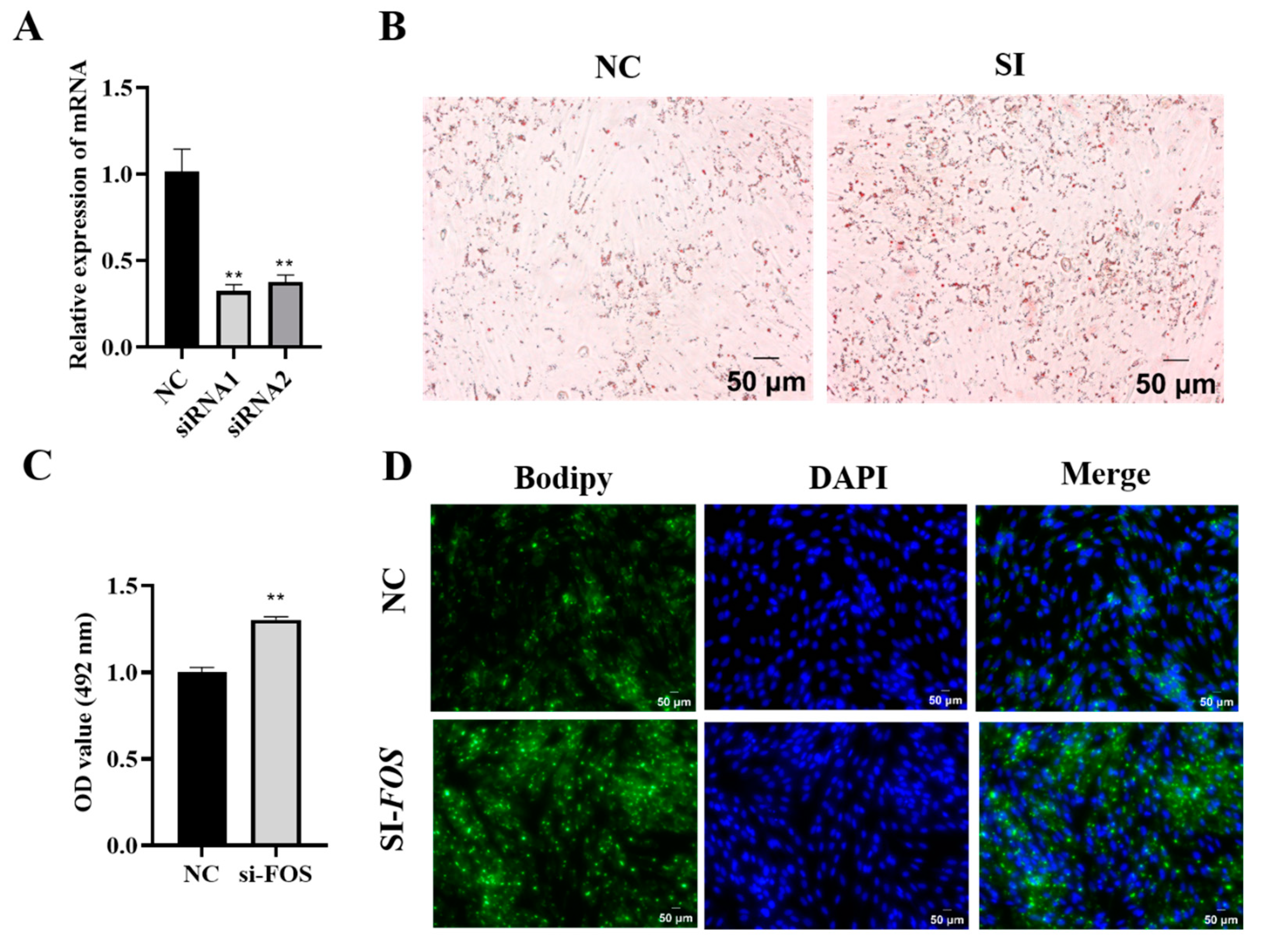
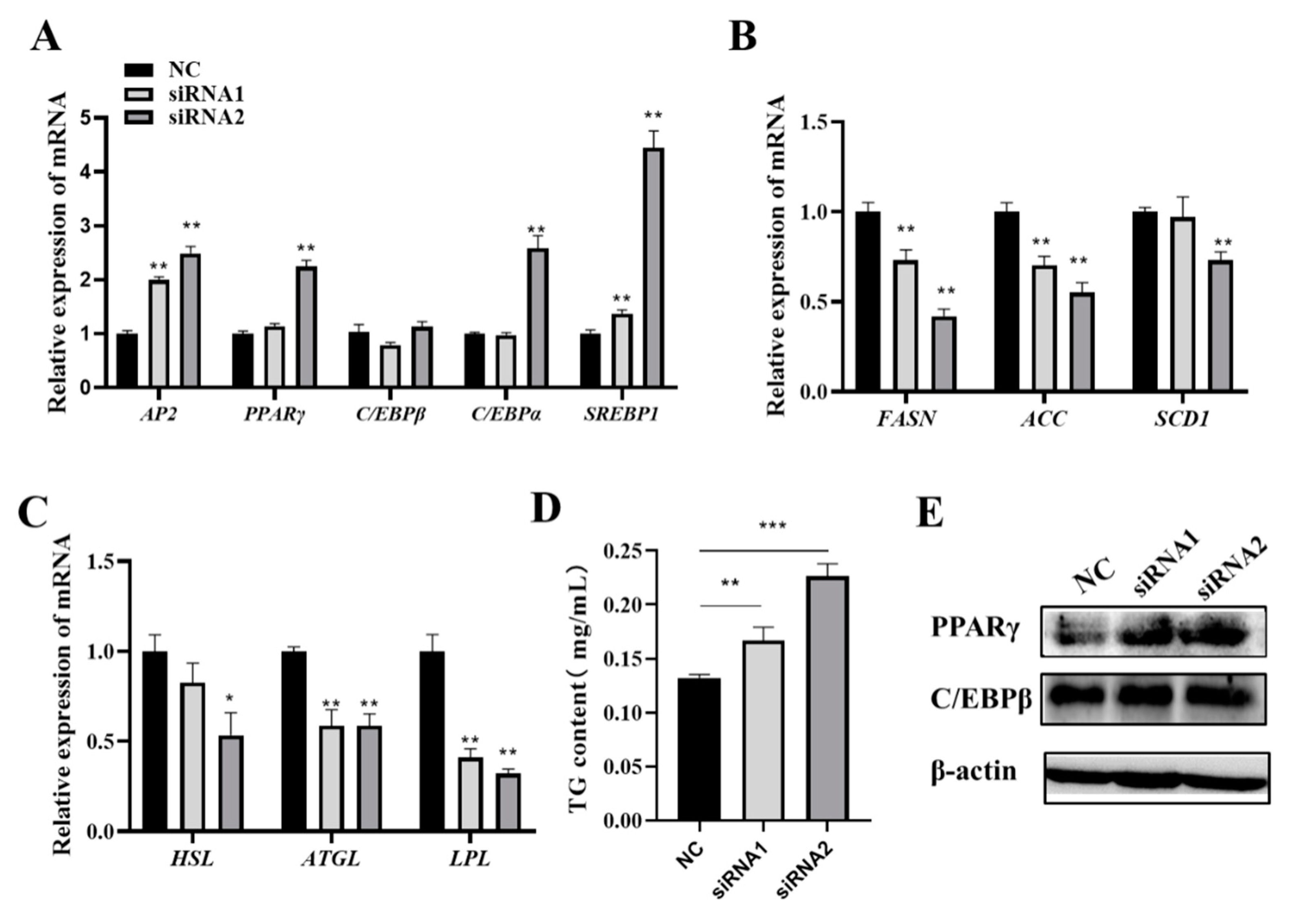
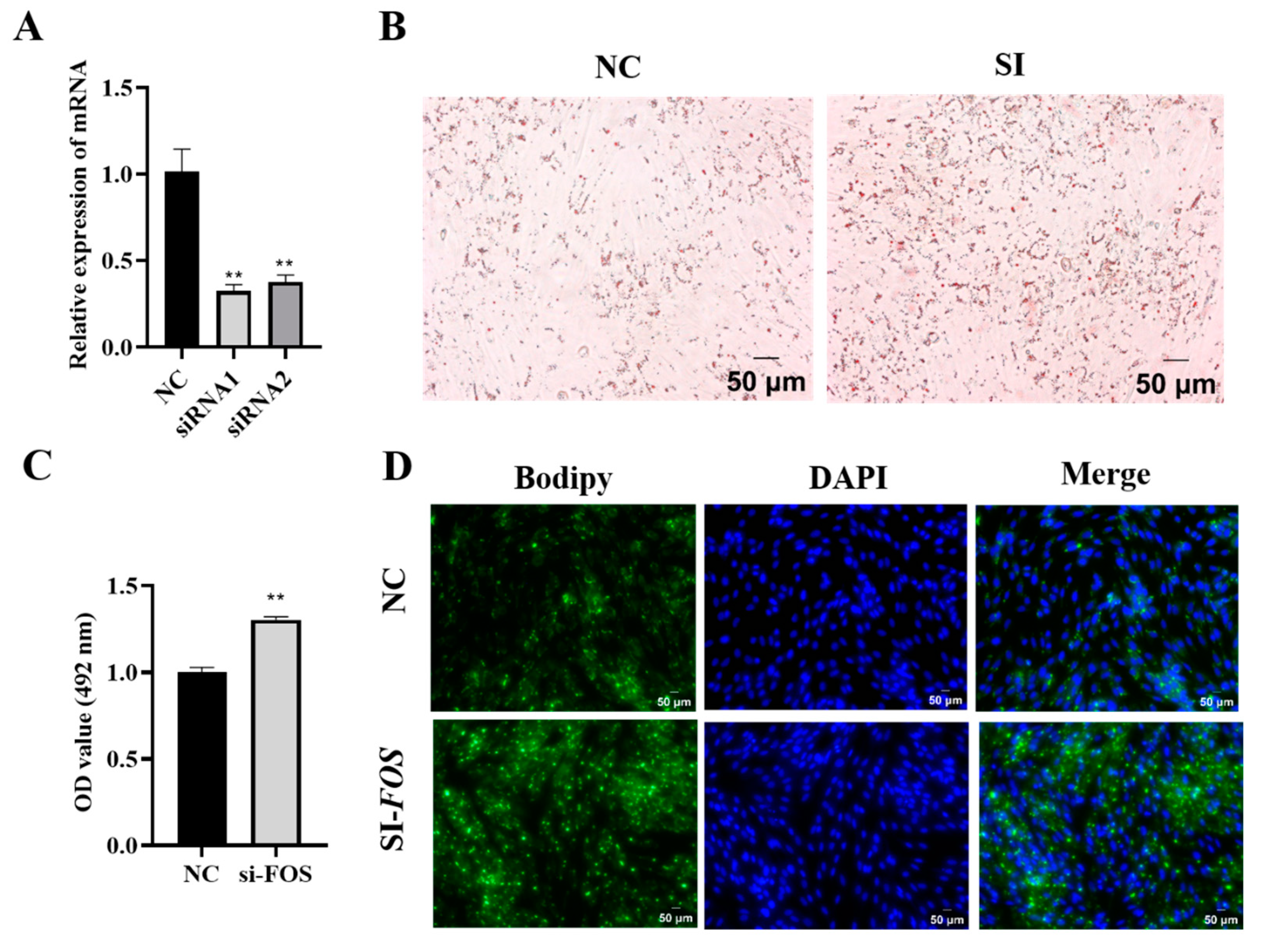
3.3. Knockdown of FOS Promotes Goat Intramuscular Preadipocyte Differentiation
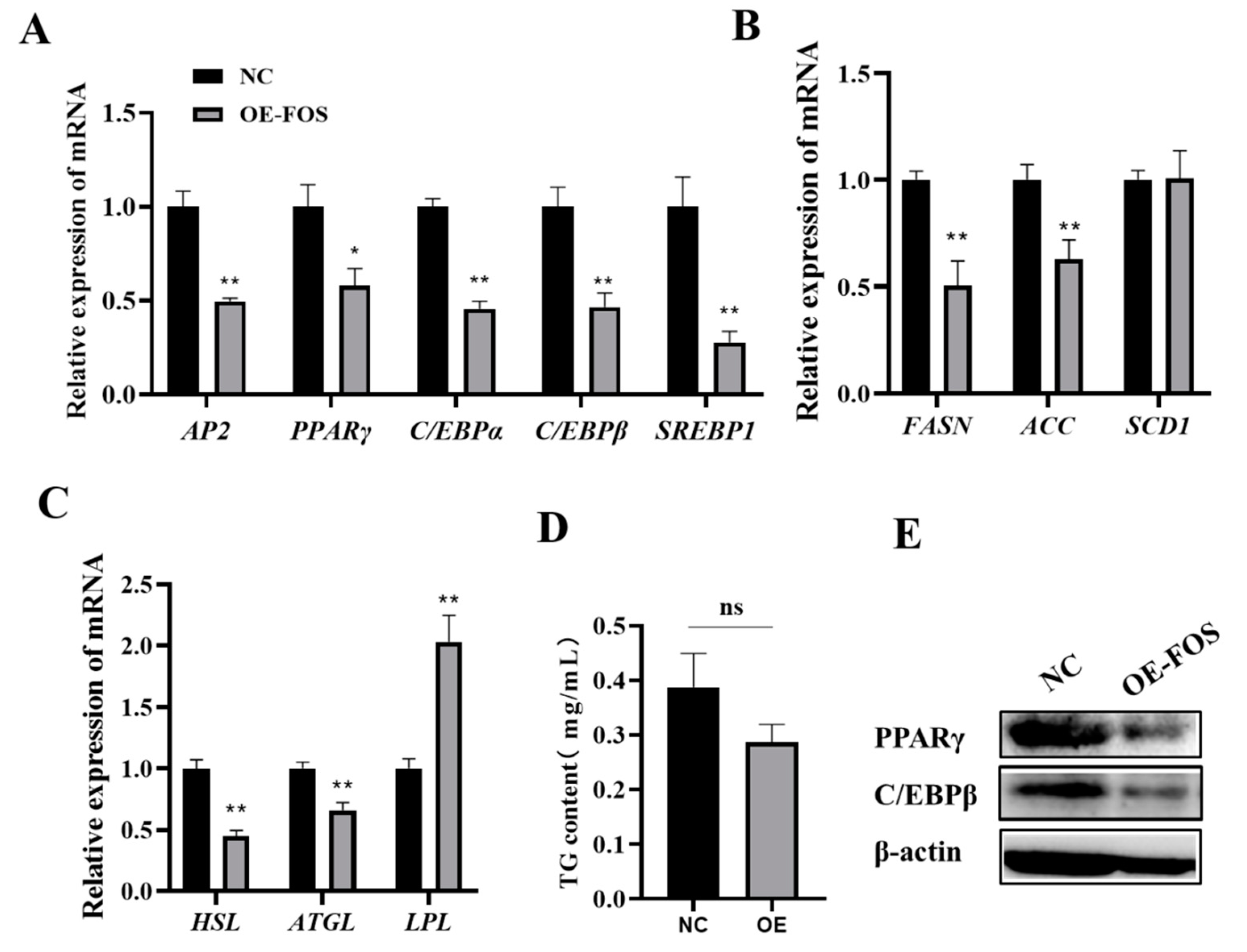
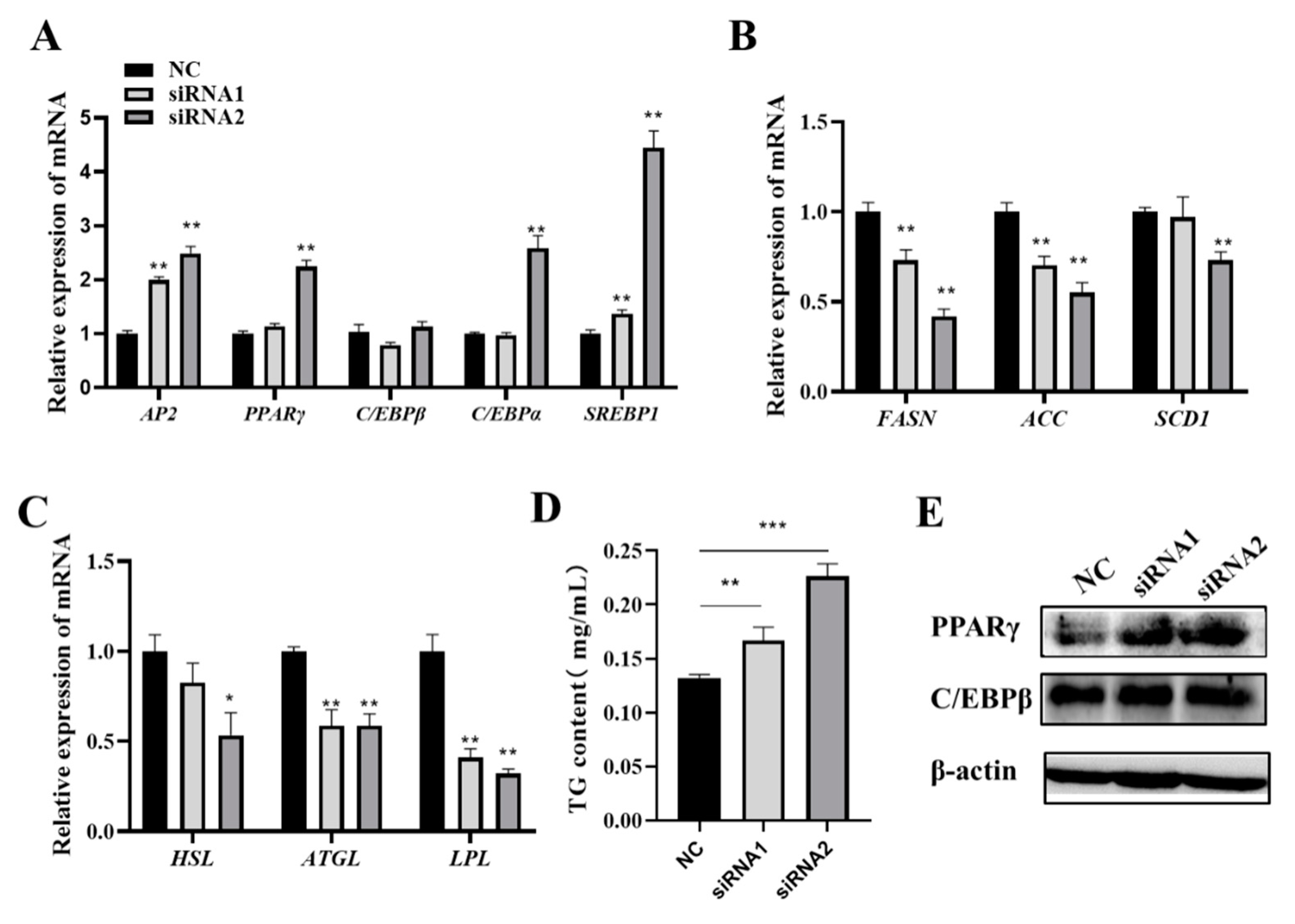
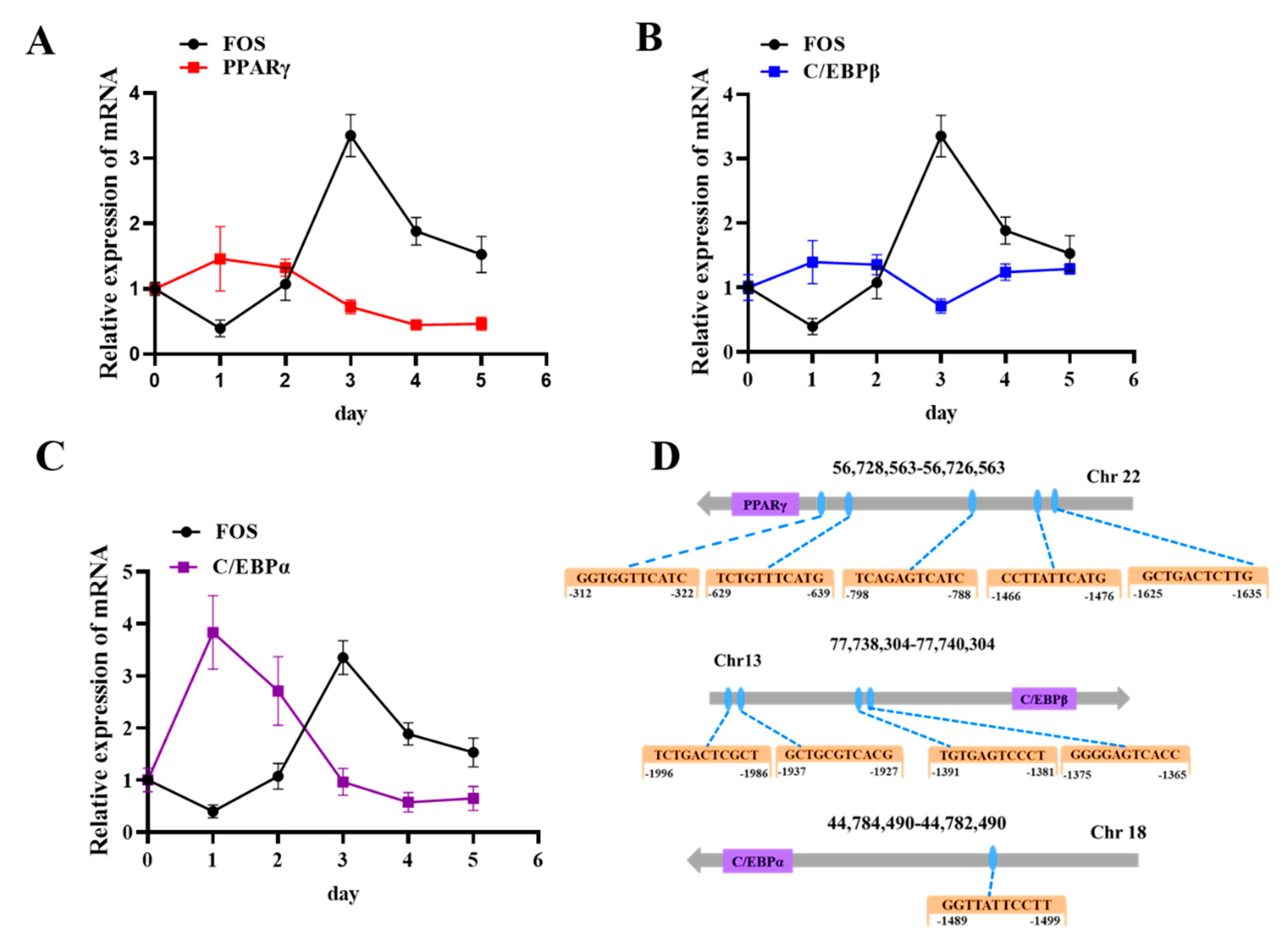
3.4. FOS Affects Intramuscular Preadipocyte Differentiation by Targeting PPARγ, C/EBPβ, and C/EBPα
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| IMF | Intramuscular fat |
| AP-1 | Activator protein-1 |
| PPARγ | Peroxisome proliferator-activated receptor γ |
| C/EBPβ | CCAAT enhancer binding protein β |
| C/EBPα | CCAAT enhancer binding protein α |
| AP2 | Fatty-acid binding protein |
| SREBP1 | Sterol regulatory element binding protein-1 |
| FASN | Fatty-acid synthase |
| ACC | Acetyl-coenzyme A carboxylase |
| SCD1 | Stearoyl-coenzyme A desaturase 1 |
| HSL | Hormone-sensitive lipase |
| ATGL | Adipose triglyceride lipase |
| LPL | Lipoprotein lipase |
| UXT | Ubiquitously expressed transcript gene |
| NC | Negative control |
| OE | Overexpression |
| HFD-fed | High-fat-diet feed |
| NASH | Nonalcoholic steatohepatitis |
| SPARC | Secreted protein acidic and rich in cysteine |
References
- Jia, W.; Di, C.; Shi, L. Applications of lipidomics in goat meat products: Biomarkers, structure, nutrition interface and future perspectives. J. Proteom. 2023, 6, 104753. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wei, T.; Liu, L.X.; Liu, J.Q.; Wang, C.X.; Yuan, Z.Y.; Ma, H.H.; Jin, H.G.; Zhang, L.C.; Cao, Y. Whole-Transcriptome Analysis of Preadipocyte and Adipocyte and Construction of Regulatory Networks to Investigate Lipid Metabolism in Sheep. Front. Genet. 2021, 29, 662143. [Google Scholar] [CrossRef] [PubMed]
- Hocquette, J.F.; Gondret, F.; Baéza, E.; Médale, F.; Jurie, C.; Pethick, D.W. Intramuscular fat content in meat-producing animals: Development, genetic and nutritional control, and identification of putative markers. Animal 2010, 4, 303–319. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Huang, J.; Wang, X.; Ma, Y. Transcription factors regulate adipocyte differentiation in beef cattle. Anim. Genet. 2020, 51, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Milde-Langosch, K. The Fos family of transcription factors and their role in tumourigenesis. Eur. J. Cancer 2005, 41, 2449–2461. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.; Rosewell, K.L.; Brännström, M.; Akin, J.W.; Curry, T.E.; Jo, M. FOS, a Critical Downstream Mediator of PGR and EGF Signaling Necessary for Ovulatory Prostaglandins in the Human Ovary. J. Clin. Endocrinol. Metab. 2018, 103, 4241–4252. [Google Scholar] [CrossRef] [PubMed]
- Dalhäusser, A.K.; Rössler, O.G.; Thiel, G. Regulation of c-Fos gene transcription by stimulus-responsive protein kinases. Gene 2022, 5, 146284. [Google Scholar] [CrossRef]
- Iwatsuki, M.; Matsuoka, M. Fluoride-induced c-Fos expression in MC3T3-E1 osteoblastic cells. Toxicol. Mech. Methods 2016, 26, 132–138. [Google Scholar] [CrossRef]
- Tong, X.; Chen, M.; Song, R.; Zhao, H.; Bian, J.; Gu, J.; Liu, Z. Overexpression of c-Fos reverses osteoprotegerin-mediated suppression of osteoclastogenesis by increasing the Beclin1-induced autophagy. J. Cell. Mol. Med. 2020, 25, 937–945. [Google Scholar] [CrossRef]
- Racca, A.C.; Prucca, C.G.; Caputto, B.L. Fra-1 and c-Fos N-Terminal Deletion Mutants Impair Breast Tumor Cell Proliferation by Blocking Lipid Synthesis Activation. Front. Oncol. 2019, 19, 544. [Google Scholar] [CrossRef]
- Distel, R.; Ro, H.; Rosen, B.; Groves, D.; Spiegelman, B. Nucleoprotein complexes that regulate gene expression in adipocyte differentiation: Direct participation of c-fos. Cell 1987, 49, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Xiao, H.; LeBlanc, S.E.; Wu, Q.; Konda, S.; Salma, N.; Marfella, C.G.A.; Ohkawa, Y.; Imbalzano, A.N. Chromatin accessibility and transcription factor binding at the PPARγ2 promoter during adipogenesis is protein kinase A-dependent. J. Cell. Physiol. 2010, 226, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Yang, P.; Liu, J.; Wu, H.; Yu, W.; Zhang, T.; Fu, H.; Liu, Y.; Hai, C. RARγ-C-Fos-PPARγ2 signaling rather than ROS generation is critical for all-trans retinoic acid-inhibited adipocyte differentiation. Biochimie 2014, 106, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Chen, K.; Wang, L.; Yang, W.; Wang, C.; Hu, G.; Mo, Z. Profiling of differentially expressed genes in adipose tissues of multiple symmetric lipomatosis. Mol. Med. Rep. 2017, 16, 6570–6579. [Google Scholar] [CrossRef] [PubMed]
- Xiong, Y.; Wang, Y.; Xu, Q.; Li, A.; Yue, Y.; Ma, Y.; Lin, Y. LKB1 Regulates Goat Intramuscular Adipogenesis Through Focal Adhesion Pathway. Front. Physiol. 2021, 13, 755598. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Lin, Y.; Zhao, N.; Wang, Y.; Li, Y. Hoxa5 Inhibits the Proliferation and Induces Adipogenic Differentiation of Subcutaneous Preadipocytes in Goats. Animals 2022, 21, 1859. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Li, Y.; Hu, T.; Gong, C.; Lu, G.; Ma, X.; Wang, Y.; Wang, Y.; Lin, Y. PDZK1-Interacting Protein 1(PDZKIP1) Inhibits Goat Subcutaneous Preadipocyte Differentiation through Promoting Autophagy. Animals 2023, 14, 1046. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Li, Y.; Lin, S.; Wang, Y.; Zhu, J.; Lin, Y. KLF4 Inhibits the Differentiation of Goat Intramuscular Preadipocytes Through Targeting C/EBPβ Directly. Front. Genet. 2021, 4, 663759. [Google Scholar] [CrossRef]
- Campos, M.; Kroon, E.; Gentz, R.; Ferreira, P. Protein domains involved in nuclear transport of Fos. Cell Biol. Int. 1999, 23, 81–88. [Google Scholar] [CrossRef]
- White, U.A.; Stephens, J.M. Transcriptional factors that promote formation of white adipose tissue. Mol. Cell. Endocrinol. 2010, 318, 10–14. [Google Scholar] [CrossRef]
- Lefterova, M.I.; Lazar, M.A. New developments in adipogenesis. Trends Endocrinol. Metab. 2009, 20, 107–114. [Google Scholar] [CrossRef]
- Barutcu, A.R.; Elizalde, G.; Gonzalez, A.E.; Soni, K.; Rinn, J.L.; Wagers, A.J.; Almada, A.E. Prolonged FOS activity disrupts a global myogenic transcriptional program by altering 3D chromatin architecture in primary muscle progenitor cells. Skelet. Muscle 2022, 15, 20. [Google Scholar] [CrossRef]
- Jing, Y.; Mu, F.; Xing, X.; Huang, J.; Lou, M.; Xu, H.; Ning, B.; Lou, Y.; Gao, Z.; Luo, H.; et al. Knockout and Restoration Reveal Differential Functional Roles of PPARγ1 and PPARγ2 in Chicken Adipogenesis. J. Agric. Food Chem. 2022, 70, 14959–14973. [Google Scholar] [CrossRef] [PubMed]
- Vidal-Puig, A.; Jimenez-Liñan, M.; Lowell, B.; Hamann, A.; Hu, E.; Spiegelman, B.; Flier, J.; Moller, D. Regulation of PPAR gamma gene expression by nutrition and obesity in rodents. J. Clin. Investig. 1996, 97, 2553–2561. [Google Scholar] [CrossRef] [PubMed]
- Gu, H.; Zhou, Y.; Yang, J.; Li, J.; Peng, Y.; Zhang, X.; Miao, Y.; Jiang, W.; Bu, G.; Hou, L.; et al. Targeted overexpression of PPARγ in skeletal muscle by random insertion and CRISPR/Cas9 transgenic pig cloning enhances oxidative fiber formation and intramuscular fat deposition. FASEB J. 2021, 35, e21308. [Google Scholar] [CrossRef] [PubMed]
- Ghaben, A.; Scherer, P. Adipogenesis and metabolic health. Nat. Rev. Mol. Cell Biol. 2019, 20, 242–258. [Google Scholar] [CrossRef] [PubMed]
- Moseti, D.; Regassa, A.; Kim, W.-K. Molecular Regulation of Adipogenesis and Potential Anti-Adipogenic Bioactive Molecules. Int. J. Mol. Sci. 2016, 19, 124. [Google Scholar] [CrossRef]
- Cao, H.; Zhang, S.; Shan, S.; Sun, C.; Li, Y.; Wang, H.; Yu, S.; Liu, Y.; Guo, F.; Zhai, Q.; et al. Ligand-dependent corepressor (LCoR) represses the transcription factor C/EBPβ during early adipocyte differentiation. J. Biol. Chem. 2017, 292, 18973–18987. [Google Scholar] [CrossRef] [PubMed]
- Davies, B.S.J.; Beigneux, A.P.; Fong, L.G.; Young, S.G. New wrinkles in lipoprotein lipase biology. Curr. Opin. Lipidol. 2012, 23, 35–42. [Google Scholar] [CrossRef]
- Liu, G.; Xu, J.-N.; Liu, D.; Ding, Q.; Liu, M.-N.; Chen, R.; Fan, M.; Zhang, Y.; Zheng, C.; Zou, D.-J.; et al. Regulation of plasma lipid homeostasis by hepatic lipoprotein lipase in adult mice. J. Lipid Res. 2016, 57, 1155–1161. [Google Scholar] [CrossRef]
- Bader, M.; Shimizu, K.; Nishimuta, S.; Fukumura, Y.; Michinaga, S.; Egusa, Y.; Hase, T.; Terada, T.; Sakurai, F.; Mizuguchi, H.; et al. Liver-specific overexpression of lipoprotein lipase improves glucose metabolism in high-fat diet-fed mice. PLoS ONE 2022, 13, e0274297. [Google Scholar]
- Zhao, L.; Li, F.; Liu, T.; Yuan, L.; Zhang, X.; Zhang, D.; Li, X.; Zhang, Y.; Zhao, Y.; Song, Q.; et al. Ovine ELOVL5 and FASN genes polymorphisms and their correlations with sheep tail fat deposition. Gene 2022, 10, 145954. [Google Scholar] [CrossRef] [PubMed]
- Hu, G.; Wang, S.; Tian, J.; Chu, L.; Li, H. Epistatic effect between ACACA and FABP2 gene on abdominal fat traits in broilers. J. Genet. Genom. 2010, 37, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Heier, C.; Kien, B.; Vesely, P.W.; Tang, Z.; Sexl, V.; Schoiswohl, G.; Strießnig-Bina, I.; Hoefler, G.; Zechner, R.; et al. Adipose triglyceride lipase activity regulates cancer cell proliferation via AMP-kinase and mTOR signaling. Biochim. Et Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2020, 1865, 158737. [Google Scholar] [CrossRef] [PubMed]
- Pajed, L.; Taschler, U.; Tilp, A.; Hofer, P.; Kotzbeck, P.; Kolleritsch, S.; Radner, F.P.W.; Pototschnig, I.; Wagner, C.; Schratter, M.; et al. Advanced lipodystrophy reverses fatty liver in mice lacking adipocyte hormone-sensitive lipase. Commun. Biol. 2021, 10, 323. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.; Xiaoli, A.; Yang, F. Regulation and Metabolic Significance of De Novo Lipogenesis in Adipose Tissues. Nutrients 2018, 29, 1383. [Google Scholar] [CrossRef] [PubMed]
- Cai, C.; Chen, D.-Z.; Tu, H.-X.; Chen, W.-K.; Ge, L.-C.; Fu, T.-T.; Tao, Y.; Ye, S.-S.; Li, J.; Lin, Z.; et al. MicroRNA-29c Acting on FOS Plays a Significant Role in Nonalcoholic Steatohepatitis Through the Interleukin-17 Signaling Pathway. Front. Physiol. 2021, 1, 597449. [Google Scholar] [CrossRef]
- Hatori, T.; Maeda, T.; Suzuki, A.; Takahashi, K.; Kato, Y. SPARC is a decoy counterpart for c-Fos and is associated with osteoblastic differentiation of bone marrow stromal cells by inhibiting adipogenesis. Mol. Med. Rep. 2023, 27, 50. [Google Scholar] [CrossRef]
- Diehl, A.M.; Yang, S.Q. Regenerative changes in C/EBPα and C/EBPß expression modulate binding to the C/EBP site in the c-fos promoter. Hepatology 1994, 19, 447–456. [Google Scholar] [CrossRef]
- Cui, T.X.; Kwok, R.; Schwartz, J. Cooperative regulation of endogenous cAMP-response element binding protein and CCAAT/enhancer-binding protein β in GH-stimulated c-fos expression. J. Endocrinol. 2007, 196, 89–100. [Google Scholar] [CrossRef]
- Hai, T.; Curran, T. Cross-family dimerization of transcription factors Fos/Jun and ATF/CREB alters DNA binding specificity. Proc. Natl. Acad. Sci. USA 1991, 88, 3720–3724. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.; Eferl, R. Fos/AP-1 proteins in bone and the immune system. Immunol. Rev. 2005, 208, 126–140. [Google Scholar] [CrossRef] [PubMed]
- Hasenfuss Sebastian, C.; Bakiri, L.; Thomsen Martin, K.; Williams Evan, G.; Auwerx, J.; Wagner Erwin, F. Regulation of Steatohepatitis and PPARγ Signaling by Distinct AP-1 Dimers. Cell Metab. 2014, 19, 84–95. [Google Scholar] [CrossRef] [PubMed]
- Cai, D.H.; Wang, D.; Keefer, J.; Yeamans, C.; Hensley, K.; Friedman, A.D. C/EBPα:AP-1 leucine zipper heterodimers bind novel DNA elements, activate the PU.1 promoter and direct monocyte lineage commitment more potently than C/EBPα homodimers or AP-1. Oncogene 2007, 27, 2772–2779. [Google Scholar] [CrossRef] [PubMed]
- Kveiborg, M.; Sabatakos, G.; Chiusaroli, R.; Wu, M.; Philbrick, W.M.; Horne, W.C.; Baron, R. ΔFosB Induces Osteosclerosis and Decreases Adipogenesis by Two Independent Cell-Autonomous Mechanisms. Mol. Cell. Biol. 2023, 24, 2820–2830. [Google Scholar] [CrossRef] [PubMed]
- Luther, J.; Driessler, F.; Megges, M.; Hess, A.; Herbort, B.; Mandic, V.; Zaiss, M.M.; Reichardt, A.; Zech, C.; Tuckermann, J.P.; et al. Elevated Fra-1 expression causes severe lipodystrophy. J. Cell Sci. 2011, 124, 1465–1476. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|
| FOS | GCTTCAACGCCGACTACGAG | AAGGAGTCTGCCGGTGAGTG |
| AP2 | TGAAGTCACTCCAGATGACAGG | TGACACATTCCAGCACCAGC |
| PPARγ | AAGCGTCAGGGTTCCACTATG | GAACCTGATGGCGTTATGAGAC |
| C/EBPβ | CAAGAAGACGGTGGACAAGC | AACAAGTTCCGCAGGGTG |
| C/EBPα | CCGTGGACAAGAACAGCAAC | AGGCGGTCATTGTCACTGGT |
| SREBP1 | AAGTGGTGGGCCTCTCTGA | GCAGGGGTTTCTCGGACT |
| FASN | TGTGCAACTGTGCCCTAG | GTCCTCTGAGCAGCGTGT |
| ACC | GGAGACAAACAGGGACCATT | ATCAGGGACTGCCGAAAC |
| SCD1 | TCGTGCCGTGGTATCTATGG | GGGGTTGATGGTCTTGTCGT |
| HSL | AGGGTCATTGCCGACTTCC | GTCTCGTTGCGTTTGTAGTGC |
| ATGL | GGAGCTTATCCAGGCCAATG | TGCGGGCAGATGTCACTCT |
| LPL | GAGGCCTTGGAGATGTGGAC | AATTGCACCGGTACGCCTTA |
| UXT | GCAAGTGGATTTGGGCTGTAAC | ATGGAGTCCTTGGTGAGGTTGT |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, T.; Li, Z.; Gong, C.; Xiong, Y.; Sun, S.; Xing, J.; Li, Y.; Li, R.; Wang, Y.; Wang, Y.; et al. FOS Inhibits the Differentiation of Intramuscular Adipocytes in Goats. Genes 2023, 14, 2088. https://doi.org/10.3390/genes14112088
Hu T, Li Z, Gong C, Xiong Y, Sun S, Xing J, Li Y, Li R, Wang Y, Wang Y, et al. FOS Inhibits the Differentiation of Intramuscular Adipocytes in Goats. Genes. 2023; 14(11):2088. https://doi.org/10.3390/genes14112088
Chicago/Turabian StyleHu, Tingting, Zhibin Li, Chengsi Gong, Yan Xiong, Shiyu Sun, Jiani Xing, Yanyan Li, Ruiwen Li, Youli Wang, Yong Wang, and et al. 2023. "FOS Inhibits the Differentiation of Intramuscular Adipocytes in Goats" Genes 14, no. 11: 2088. https://doi.org/10.3390/genes14112088
APA StyleHu, T., Li, Z., Gong, C., Xiong, Y., Sun, S., Xing, J., Li, Y., Li, R., Wang, Y., Wang, Y., & Lin, Y. (2023). FOS Inhibits the Differentiation of Intramuscular Adipocytes in Goats. Genes, 14(11), 2088. https://doi.org/10.3390/genes14112088