
Oncomeric Profiles of microRNAs as New Therapeutic Targets for Treatment of Ewing’s Sarcoma: A Composite Review


 and
and 
Abstract
:1. Introduction
2. Biosynthesis of miRs and Their Involvement in Cancers
3. miRs in ES Development and Progression
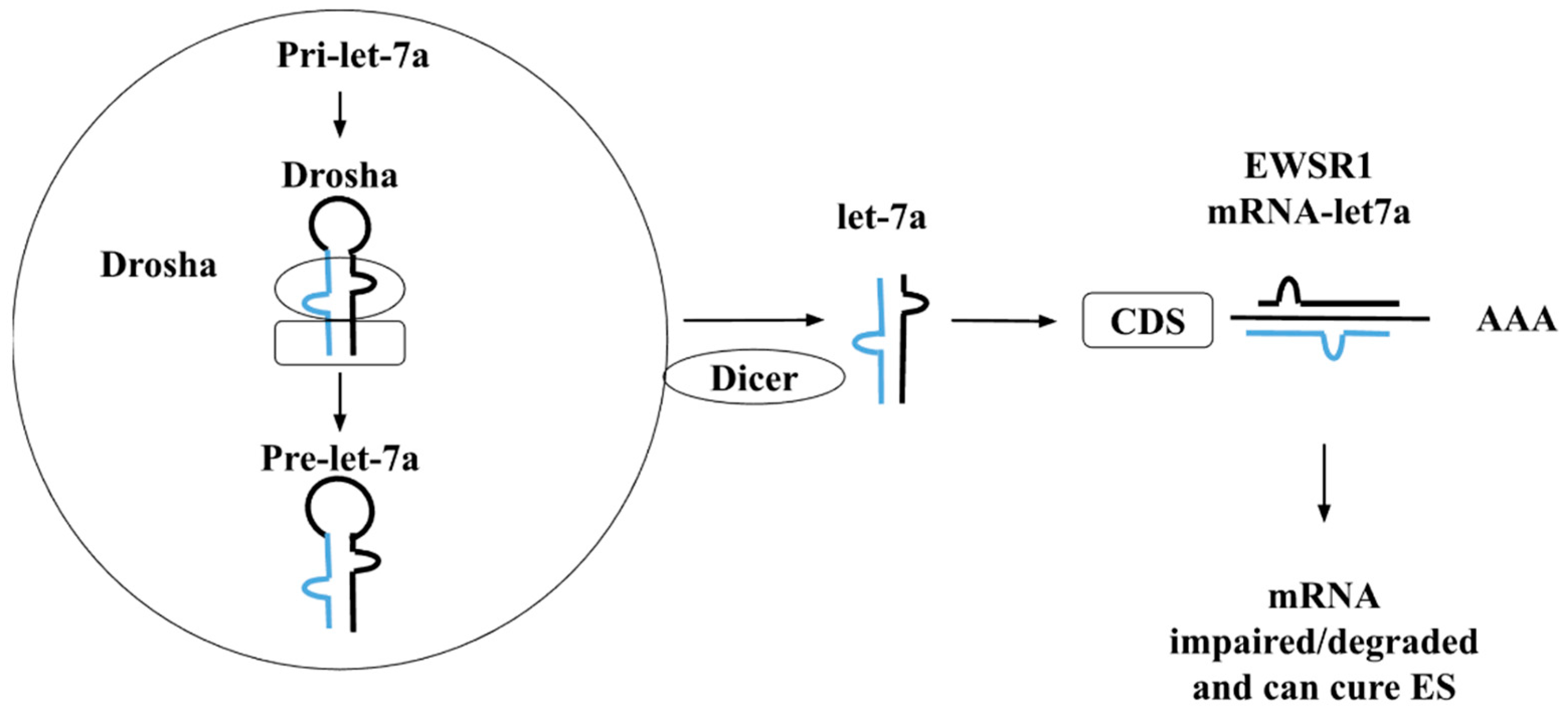
4. The Functional Role of Let-7 Family in ES
5. miRs as Therapeutic Agents and Applications
6. Conclusions and Future Prospectus
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Roundhill, E.A.; Pantziarka, P.; Liddle, D.E.; Shaw, L.A.; Albadrani, G.; Burchill, S.A. Exploiting the stemness and chemoresistance transcriptome of Ewing sarcoma to identify candidate therapeutic targets and drug-repurposing candidates. Cancers 2023, 15, 769. [Google Scholar] [CrossRef] [PubMed]
- Yasir, M.; Park, J.; Han, E.-T.; Park, W.S.; Han, J.-H.; Kwon, Y.-S.; Lee, H.-J.; Hassan, M.; Kloczkowski, A.; Chun, W. Exploration of Flavonoids as Lead Compounds against Ewing Sarcoma through Molecular Docking, Pharmacogenomics Analysis, and Molecular Dynamics Simulations. Molecules 2023, 28, 414. [Google Scholar] [CrossRef] [PubMed]
- Caltavituro, A.; Buonaiuto, R.; Salomone, F.; Morra, R.; Pietroluongo, E.; De Placido, P.; Tortora, M.; Peddio, A.; Picozzi, F.; Ottaviano, M. Extraskeletal Ewing’s sarcoma of the mediastinum: Case report. Front. Oncol. 2023, 13, 1074378. [Google Scholar] [CrossRef]
- Burchill, S. Ewing’s sarcoma: Diagnostic, prognostic, and therapeutic implications of molecular abnormalities. J. Clin. Pathol. 2003, 56, 96–102. [Google Scholar] [CrossRef]
- Chen, F.; Wang, S.; Wei, Y.; Wu, J.; Huang, G.; Chen, J.; Shi, J.; Xia, J. Norcantharidin modulates the miR-30a/Metadherin/AKT signaling axis to suppress proliferation and metastasis of stromal tumor cells in giant cell tumor of bone. Biomed. Pharmacother. 2018, 103, 1092–1100. [Google Scholar] [CrossRef]
- Herrero-Martin, D.; Fourtouna, A.; Niedan, S.; Riedmann, L.T.; Schwentner, R.; Aryee, D.N. Factors affecting EWS-FLI1 activity in Ewing’s sarcoma. Sarcoma 2011, 2011, 352580. [Google Scholar] [CrossRef]
- Garcia-Echeverria, C.; Hoffmann, F.; Trumpp, A.; Stamenkovic, I. Development of Ewing’s Sarcoma from Primary Bone Marrow–Derived Mesenchymal Progenitor Cells. Cancer Res. 2005, 65, 11459–11468. [Google Scholar]
- Boro, A.; Prêtre, K.; Rechfeld, F.; Thalhammer, V.; Oesch, S.; Wachtel, M.; Schäfer, B.W.; Niggli, F.K. Small-molecule screen identifies modulators of EWS/FLI1 target gene expression and cell survival in Ewing’s sarcoma. Int. J. Cancer 2012, 131, 2153–2164. [Google Scholar] [CrossRef]
- Palmini, G.; Brandi, M.L. microRNAs and bone tumours: Role of tiny molecules in the development and progression of chondrosarcoma, of giant cell tumour of bone and of Ewing’s sarcoma. Bone 2021, 149, 115968. [Google Scholar] [CrossRef]
- Nishimori, H.; Sasaki, Y.; Yoshida, K.; Irifune, H.; Zembutsu, H.; Tanaka, T.; Aoyama, T.; Hosaka, T.; Kawaguchi, S.; Wada, T. The Id2 gene is a novel target of transcriptional activation by EWS-ETS fusion proteins in Ewing family tumors. Oncogene 2002, 21, 8302–8309. [Google Scholar] [CrossRef]
- Cotterill, S.; Ahrens, S.; Paulussen, M.; Jurgens, H.; Voute, P.; Gadner, H.; Craft, A. Prognostic factors in Ewing’s tumor of bone: Analysis of 975 patients from the European Intergroup Cooperative Ewing’s Sarcoma Study Group. J. Clin. Oncol. 2000, 18, 3108–3114. [Google Scholar] [CrossRef] [PubMed]
- McKinsey, E.; Parrish, J.; Irwin, A.; Niemeyer, B.; Kern, H.; Birks, D.; Jedlicka, P. A novel oncogenic mechanism in Ewing sarcoma involving IGF pathway targeting by EWS/Fli1-regulated microRNAs. Oncogene 2011, 30, 4910–4920. [Google Scholar] [CrossRef] [PubMed]
- Dylla, L.; Moore, C.; Jedlicka, P. MicroRNAs in Ewing sarcoma. Front. Oncol. 2013, 3, 46128. [Google Scholar] [CrossRef] [PubMed]
- Parafioriti, A.; Bason, C.; Armiraglio, E.; Calciano, L.; Daolio, P.A.; Berardocco, M.; Di Bernardo, A.; Colosimo, A.; Luksch, R.; Berardi, A.C. Ewing’s sarcoma: An analysis of miRNA expression profiles and target genes in paraffin-embedded primary tumor tissue. Int. J. Mol. Sci. 2016, 17, 656. [Google Scholar] [CrossRef]
- Li, Z.; Yu, X.; Shen, J.; Wu, W.K.K.; Chan, M.T. MicroRNA expression and its clinical implications in Ewing’s sarcoma. Cell Prolif. 2015, 48, 1–6. [Google Scholar] [CrossRef]
- Sassen, S.; Miska, E.A.; Caldas, C. MicroRNA—Implications for cancer. Virchows Arch. 2008, 452, 1–10. [Google Scholar] [CrossRef]
- Li, M.A.; He, L. microRNAs as novel regulators of stem cell pluripotency and somatic cell reprogramming. Bioessays 2012, 34, 670–680. [Google Scholar] [CrossRef] [PubMed]
- Crist, C.G.; Montarras, D.; Pallafacchina, G.; Rocancourt, D.; Cumano, A.; Conway, S.J.; Buckingham, M. Muscle stem cell behavior is modified by microRNA-27 regulation of Pax3 expression. Proc. Natl. Acad. Sci. USA 2009, 106, 13383–13387. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Dey, B.K.; Dutta, A. MiR-322/424 and-503 are induced during muscle differentiation and promote cell cycle quiescence and differentiation by down-regulation of Cdc25A. Mol. Biol. Cell 2010, 21, 2138–2149. [Google Scholar] [CrossRef]
- Khach Lai, V.; Ashraf, M.; Jiang, S.; Haider, K. MicroRNA-143 is a critical regulator of cell cycle activity in stem cells with co-overexpression of Akt and angiopoietin-1 via transcriptional regulation of Erk5/cyclin D1 signaling. Cell Cycle 2012, 11, 767–777. [Google Scholar] [CrossRef]
- Wahid, F.; Shehzad, A.; Khan, T.; Kim, Y.Y. MicroRNAs: Synthesis, mechanism, function, and recent clinical trials. Biochim. Et Biophys. Acta (BBA)-Mol. Cell Res. 2010, 1803, 1231–1243. [Google Scholar] [CrossRef]
- Hassan, M.; Iqbal, M.S.; Naqvi, S.; Alashwal, H.; Moustafa, A.A.; Kloczkowski, A. Prediction of Site Directed miRNAs as Key Players of Transcriptional Regulators Against Influenza C Virus Infection Through Computational Approaches. Front. Mol. Biosci. 2022, 9, 866072. [Google Scholar] [CrossRef]
- Wojciechowska, A.; Braniewska, A.; Kozar-Kamińska, K. MicroRNA in cardiovascular biology and disease. Adv. Clin. Exp. Med. Off. Organ Wroc. Med. Univ. 2017, 26, 865–874. [Google Scholar] [CrossRef]
- Packer, A.N.; Xing, Y.; Harper, S.Q.; Jones, L.; Davidson, B.L. The bifunctional microRNA miR-9/miR-9* regulates REST and CoREST and is downregulated in Huntington’s disease. J. Neurosci. 2008, 28, 14341–14346. [Google Scholar] [CrossRef] [PubMed]
- Hébert, S.S.; Horré, K.; Nicolaï, L.; Papadopoulou, A.S.; Mandemakers, W.; Silahtaroglu, A.N.; Kauppinen, S.; Delacourte, A.; De Strooper, B. Loss of microRNA cluster miR-29a/b-1 in sporadic Alzheimer’s disease correlates with increased BACE1/β-secretase expression. Proc. Natl. Acad. Sci. USA 2008, 105, 6415–6420. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Esser, K.A. MicroRNA-1 and microRNA-133a expression are decreased during skeletal muscle hypertrophy. J. Appl. Physiol. 2007, 102, 306–313. [Google Scholar] [CrossRef]
- Maroney, P.A.; Yu, Y.; Fisher, J.; Nilsen, T.W. Evidence that microRNAs are associated with translating messenger RNAs in human cells. Nat. Struct. Mol. Biol. 2006, 13, 1102–1107. [Google Scholar] [CrossRef]
- Jackson, R.J.; Standart, N. How do microRNAs regulate gene expression? Sci. STKE 2007, 2007, re1. [Google Scholar] [CrossRef] [PubMed]
- Loganathan, T.; Doss C, G.P. Non-coding RNAs in human health and disease: Potential function as biomarkers and therapeutic targets. Funct. Integr. Genom. 2023, 23, 33. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, N.; Al Samarrai, O.R.; Al-Zaidi, H.M.H.; Mirzaei, A.R.; Heidari, M.J. Comprehensive analysis of microRNA (miRNA) in cancer cells. Cell. Mol. Biomed. Rep. 2023, 3, 89–97. [Google Scholar] [CrossRef]
- Baran, Y. MicroRNAs and Long Non-Coding RNAs as Novel Targets in Anti-Cancer Drug Development. Curr. Pharm. Biotechnol. 2023, 24, 913–925. [Google Scholar]
- Calin, G.A.; Dumitru, C.D.; Shimizu, M.; Bichi, R.; Zupo, S.; Noch, E.; Aldler, H.; Rattan, S.; Keating, M.; Rai, K. Frequent deletions and down-regulation of micro-RNA genes miR15 and miR16 at 13q14 in chronic lymphocytic leukemia. Proc. Natl. Acad. Sci. USA 2002, 99, 15524–15529. [Google Scholar] [CrossRef] [PubMed]
- Dalmay, T.; Edwards, D. MicroRNAs and the hallmarks of cancer. Oncogene 2006, 25, 6170–6175. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.; Hou, W. Regulation of angiogenesis by microRNAs in cancer. Mol. Med. Rep. 2021, 24, 583. [Google Scholar] [CrossRef]
- Huang, Q.; Gumireddy, K.; Schrier, M.; Le Sage, C.; Nagel, R.; Nair, S.; Egan, D.A.; Li, A.; Huang, G.; Klein-Szanto, A.J. The microRNAs miR-373 and miR-520c promote tumour invasion and metastasis. Nat. Cell Biol. 2008, 10, 202–210. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.F.; Chin, A.R. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 2014, 25, 501–515. [Google Scholar] [CrossRef] [PubMed]
- Soheilyfar, S.; Velashjerdi, Z.; Hajizadeh, Y.S.; Maroufi, N.F.; Amini, Z.; Khorrami, A.; Azimian, S.H.; Isazadeh, A.; Taefehshokr, S.; Taefehshokr, N. In vivo and in vitro impact of miR-31 and miR-143 on the suppression of metastasis and invasion in breast cancer. J. BUON 2018, 23, 1290–1296. [Google Scholar] [PubMed]
- Chou, J.; Lin, J.H.; Brenot, A.; Kim, J.-w.; Provot, S.; Werb, Z. GATA3 suppresses metastasis and modulates the tumour microenvironment by regulating microRNA-29b expression. Nat. Cell Biol. 2013, 15, 201–213. [Google Scholar] [CrossRef]
- Tay, Y.; Zhang, J.; Thomson, A.M.; Lim, B.; Rigoutsos, I. MicroRNAs to Nanog, Oct4 and Sox2 coding regions modulate embryonic stem cell differentiation. Nature 2008, 455, 1124–1128. [Google Scholar] [CrossRef]
- Xu, N.; Papagiannakopoulos, T.; Pan, G.; Thomson, J.A.; Kosik, K.S. MicroRNA-145 regulates OCT4, SOX2, and KLF4 and represses pluripotency in human embryonic stem cells. Cell 2009, 137, 647–658. [Google Scholar] [CrossRef]
- Zoni, E.; Van Der Horst, G.; Van de Merbel, A.; Chen, L.; Rane, J.; Pelger, R.; Collins, A.; Visakorpi, T.; Snaar-Jagalska, B.; Maitland, N. miR-25 modulates invasiveness and dissemination of human prostate cancer cells via regulation of αv-and α6-integrin expression. Cancer Res. 2015, 75, 2326–2336. [Google Scholar] [CrossRef]
- Hwang, W.-L.; Jiang, J.-K.; Yang, S.-H.; Huang, T.-S.; Lan, H.-Y.; Teng, H.-W.; Yang, C.-Y.; Tsai, Y.-P.; Lin, C.-H.; Wang, H.-W. MicroRNA-146a directs the symmetric division of Snail-dominant colorectal cancer stem cells. Nat. Cell Biol. 2014, 16, 268–280. [Google Scholar] [CrossRef]
- Li, H.-Y.; Liang, J.-L.; Kuo, Y.-L.; Lee, H.-H.; Calkins, M.J.; Chang, H.-T.; Lin, F.-C.; Chen, Y.-C.; Hsu, T.-I.; Hsiao, M. miR-105/93-3p promotes chemoresistance and circulating miR-105/93-3p acts as a diagnostic biomarker for triple negative breast cancer. Breast Cancer Res. 2017, 19, 133. [Google Scholar] [CrossRef]
- Franzetti, G.; Laud-Duval, K.; Bellanger, D.; Stern, M.; Sastre-Garau, X.; Delattre, O. MiR-30a-5p connects EWS-FLI1 and CD99, two major therapeutic targets in Ewing tumor. Oncogene 2013, 32, 3915–3921. [Google Scholar] [CrossRef]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. MicroRNA-20b promotes cell proliferation via targeting of TGF-β receptor II and upregulates MYC expression in Ewing’s sarcoma cells. Int. J. Oncol. 2017, 51, 1842–1850. [Google Scholar] [CrossRef] [PubMed]
- Satterfield, L.; Shuck, R.; Kurenbekova, L.; Allen-Rhoades, W.; Edwards, D.; Huang, S.; Rajapakshe, K.; Coarfa, C.; Donehower, L.A.; Yustein, J.T. miR-130b directly targets Arhgap1 to drive activation of a metastatic CDC42-PAK1-AP1 positive feedback loop in Ewing sarcoma. Int. J. Cancer 2017, 141, 2062–2075. [Google Scholar] [CrossRef]
- Lu, Q.; Lu, M.; Li, D.; Zhang, S. MicroRNA-34b promotes proliferation, migration and invasion of Ewing’s sarcoma cells by downregulating Notch1. Mol. Med. Rep. 2018, 18, 3577–3588. [Google Scholar] [CrossRef]
- Lobry, C.; Oh, P.; Aifantis, I. Oncogenic and tumor suppressor functions of Notch in cancer: It’s NOTCH what you think. J. Exp. Med. 2011, 208, 1931–1935. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yang, F.; Xu, H.; Yue, Z.; Fang, X.; Liu, J. MicroRNA-708 is downregulated in hepatocellular carcinoma and suppresses tumor invasion and migration. Biomed. Pharmacother. 2015, 73, 154–159. [Google Scholar] [CrossRef] [PubMed]
- Lei, S.-L.; Zhao, H.; Yao, H.-L.; Chen, Y.; Lei, Z.-D.; Liu, K.-J.; Yang, Q. Regulatory roles of microRNA-708 and microRNA-31 in proliferation, apoptosis and invasion of colorectal cancer cells. Oncol. Lett. 2014, 8, 1768–1774. [Google Scholar] [CrossRef] [PubMed]
- Roberto, G.M.; Vieira, G.M.; Delsin, L.E.A.; de Oliveira Silva, M.; Hakime, R.G.; Engel, E.E.; Scrideli, C.A.; Tone, L.G.; Brassesco, M.S. MiR-708-5p is inversely associated with EWS/FLI1 Ewing sarcoma but does not represent a prognostic predictor. Cancer Genet. 2019, 230, 21–27. [Google Scholar] [CrossRef]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. MicroRNA-181c prevents apoptosis by targeting of FAS receptor in Ewing’s sarcoma cells. Cancer Cell Int. 2018, 18, 37. [Google Scholar] [CrossRef]
- Gao, X.-N.; Lin, J.; Gao, L.; Li, Y.-H.; Wang, L.-L.; Yu, L. MicroRNA-193b regulates c-Kit proto-oncogene and represses cell proliferation in acute myeloid leukemia. Leuk. Res. 2011, 35, 1226–1232. [Google Scholar] [CrossRef]
- Li, Y.; Shao, G.; Zhang, M.; Zhu, F.; Zhao, B.; He, C.; Zhang, Z. miR-124 represses the mesenchymal features and suppresses metastasis in Ewing sarcoma. Oncotarget 2017, 8, 10274. [Google Scholar] [CrossRef] [PubMed]
- Moore, C.; Parrish, J.K.; Jedlicka, P. MiR-193b, downregulated in Ewing Sarcoma, targets the ErbB4 oncogene to inhibit anchorage-independent growth. PLoS ONE 2017, 12, e0178028. [Google Scholar] [CrossRef] [PubMed]
- Mendoza-Naranjo, A.; El-Naggar, A.; Wai, D.H.; Mistry, P.; Lazic, N.; Ayala, F.R.R.; da Cunha, I.W.; Rodriguez-Viciana, P.; Cheng, H.; Tavares Guerreiro Fregnani, J.H. ERBB 4 confers metastatic capacity in ewing sarcoma. EMBO Mol. Med. 2013, 5, 1087–1102. [Google Scholar] [CrossRef] [PubMed]
- Esposito, F.; Tornincasa, M.; Pallante, P.; Federico, A.; Borbone, E.; Pierantoni, G.M.; Fusco, A. Down-regulation of the miR-25 and miR-30d contributes to the development of anaplastic thyroid carcinoma targeting the polycomb protein EZH2. J. Clin. Endocrinol. 2012, 97, E710–E718. [Google Scholar] [CrossRef]
- Chen, D.; Guo, W.; Qiu, Z.; Wang, Q.; Li, Y.; Liang, L.; Liu, L.; Huang, S.; Zhao, Y.; He, X. MicroRNA-30d-5p inhibits tumour cell proliferation and motility by directly targeting CCNE2 in non-small cell lung cancer. Cancer Lett. 2015, 362, 208–217. [Google Scholar] [CrossRef]
- Zenali, M.J.; Zhang, P.L.; Bendel, A.E.; Brown, R.E. Morphoproteomic confirmation of constitutively activated mTOR, ERK, and NF-kappaB pathways in Ewing family of tumors. Ann. Clin. Lab. Sci. 2009, 39, 160–166. [Google Scholar]
- Roskoski, R., Jr. ERK1/2 MAP kinases: Structure, function, and regulation. Pharmacol. Res. 2012, 66, 105–143. [Google Scholar] [CrossRef]
- Chen, J.; Zhou, X.; Xiao, Q.; Wang, T.; Shao, G.; Li, Y.; Zhang, Z. MiR-107 suppresses cell proliferation and tube formation of Ewing sarcoma cells partly by targeting HIF-1β. Hum. Cell 2018, 31, 42–49. [Google Scholar] [CrossRef]
- Liu, C.; Li, G.; Ren, S.; Su, Z.; Wang, Y.; Tian, Y.; Liu, Y.; Qiu, Y. miR-185-3p regulates the invasion and metastasis of nasopharyngeal carcinoma by targeting WNT2B in vitro. Oncol. Lett. 2017, 13, 2631–2636. [Google Scholar] [CrossRef]
- Li, S.; Ma, Y.; Hou, X.; Liu, Y.; Li, K.; Xu, S.; Wang, J. MiR-185 acts as a tumor suppressor by targeting AKT1 in non-small cell lung cancer cells. Int. J. Clin. Exp. Pathol. 2015, 8, 11854. [Google Scholar]
- Ordóñez, J.L.; Osuna, D.; Herrero, D.; de Alava, E.; Madoz-Gúrpide, J. Advances in Ewing’s sarcoma research: Where are we now and what lies ahead? Cancer Res. 2009, 69, 7140–7150. [Google Scholar] [CrossRef]
- Zhou, X.; Chen, J.; Xiao, Q.; Wang, T.; Yu, Y.; Li, B.; Shao, G.; Li, Y.; Zhang, Z. MicroRNA-638 inhibits cell growth and tubule formation by suppressing VEGFA expression in human Ewing sarcoma cells. Biosci. Rep. 2018, 38, BSR20171017. [Google Scholar] [CrossRef]
- Liu, Y.; Chen, G.; Liu, H.; Li, Z.; Yang, Q.; Gu, X.; Du, Z.; Zhang, G.; Wang, J. Integrated bioinformatics analysis of miRNA expression in Ewing sarcoma and potential regulatory effects of miR-21 via targeting ALCAM/CD166. Artif. Cells Nanomed. Biotechnol. 2019, 47, 2114–2122. [Google Scholar] [CrossRef]
- Rodrigues, D.S.; Monteiro, V.S.; Navegantes-Lima, K.C.; Oliveira, A.; Gaspar, S.; Quadros, L.G.; Monteiro, M.C. MicroRNAs in cell cycle progression and proliferation: Molecular mechanisms and pathways. Non-Coding RNA Investig. 2018, 2, 28. [Google Scholar] [CrossRef]
- Sun, D.; Wu, Y.; Zhang, S.; Han, Y.; Shen, J.; Zheng, W.; Wei, L.; Liu, Y.; Ren, L.; Gu, Z. Distinct roles of miR-34 family members on suppression of lung squamous cell carcinoma. Biomed. Pharmacother. 2021, 142, 111967. [Google Scholar] [CrossRef]
- Barrett, C.; Budhiraja, A.; Parashar, V.; Batish, M. The landscape of regulatory noncoding rnas in ewing’s sarcoma. Biomedicines 2021, 9, 933. [Google Scholar] [CrossRef]
- Ye, C.; Yu, X.; Liu, X.; Dai, M.; Zhang, B. miR-30d inhibits cell biological progression of Ewing’s sarcoma by suppressing the MEK/ERK and PI3K/Akt pathways in vitro Corrigendum in/10.3892/ol. 2020.12394. Oncol. Lett. 2018, 15, 4390–4396. [Google Scholar]
- Zhang, S.; Li, D.; Jiao, G.-J.; Wang, H.-L.; Yan, T.-B. miR-185 suppresses progression of Ewing’s sarcoma via inhibiting the PI3K/AKT and Wnt/β-catenin pathways. OncoTargets Ther. 2018, 11, 7967. [Google Scholar] [CrossRef]
- Huang, W.-K.; Yeh, C.-N. The emerging role of MicroRNAs in regulating the drug response of cholangiocarcinoma. Biomolecules 2020, 10, 1396. [Google Scholar] [CrossRef]
- De Feo, A.; Pazzaglia, L.; Ciuffarin, L.; Mangiagli, F.; Pasello, M.; Simonetti, E.; Pellegrini, E.; Ferrari, C.; Bianchi, G.; Spazzoli, B. miR-214-3p Is Commonly Downregulated by EWS-FLI1 and by CD99 and Its Restoration Limits Ewing Sarcoma Aggressiveness. Cancers 2022, 14, 1762. [Google Scholar] [CrossRef]
- Nugent, M. microRNA and bone cancer. Microrna Cancer 2015, 889, 201–230. [Google Scholar]
- Nugent, M. MicroRNA function and dysregulation in bone tumors: The evidence to date. Cancer Manag. Res. 2014, 6, 15. [Google Scholar] [CrossRef]
- Boyerinas, B.; Park, S.-M.; Hau, A.; Murmann, A.E.; Peter, M.E. The role of let-7 in cell differentiation and cancer. Endocr. -Relat. Cancer 2010, 17, F19–F36. [Google Scholar] [CrossRef]
- De Vito, C.; Riggi, N.; Suva, M.-L.; Janiszewska, M.; Horlbeck, J.; Baumer, K.; Provero, P.; Stamenkovic, I. Let-7a is a direct EWS-FLI-1 target implicated in Ewing’s sarcoma development. PLoS ONE 2011, 6, e23592. [Google Scholar] [CrossRef]
- Dong, Q.; Meng, P.; Wang, T.; Qin, W.; Qin, W.; Wang, F.; Yuan, J.; Chen, Z.; Yang, A.; Wang, H. MicroRNA let-7a inhibits proliferation of human prostate cancer cells in vitro and in vivo by targeting E2F2 and CCND2. PLoS ONE 2010, 5, e10147. [Google Scholar] [CrossRef]
- Zhang, Z.; Li, Y.; Huang, L.; Xiao, Q.; Chen, X.; Zhong, J.; Chen, Y.; Yang, D.; Han, Z.; Shu, Y. Let-7a suppresses macrophage infiltrations and malignant phenotype of Ewing sarcoma via STAT3/NF-κB positive regulatory circuit. Cancer Lett. 2016, 374, 192–201. [Google Scholar] [CrossRef]
- Sand, L.G.; Berghuis, D.; Szuhai, K.; Hogendoorn, P.C. Expression of CCL21 in Ewing sarcoma shows an inverse correlation with metastases and is a candidate target for immunotherapy. Cancer Immunol. Immunother. 2016, 65, 995–1002. [Google Scholar] [CrossRef]
- Bill, C.A.; Allen, C.M.; Vines, C.M. CC chemokine receptor 7 in cancer. Cells 2022, 11, 656. [Google Scholar] [CrossRef]
- Soini, Y. Epigenetic and genetic changes in soft tissue sarcomas: A review. Apmis 2016, 124, 925–934. [Google Scholar] [CrossRef]
- Fu, T.-Y.; Chang, C.-C.; Lin, C.-T.; Lai, C.-H.; Peng, S.-Y.; Ko, Y.-J.; Tang, P.-C. Let-7b-mediated suppression of basigin expression and metastasis in mouse melanoma cells. Exp. Cell Res. 2011, 317, 445–451. [Google Scholar] [CrossRef]
- Hameiri-Grossman, M.; Porat-Klein, A.; Yaniv, I.; Ash, S.; Cohen, I.J.; Kodman, Y.; Haklai, R.; Elad-Sfadia, G.; Kloog, Y.; Chepurko, E. The association between let-7, RAS and HIF-1α in Ewing Sarcoma tumor growth. Oncotarget 2015, 6, 33834. [Google Scholar] [CrossRef]
- Sohn, E.J.; Park, J.; Wu, Y.-P. Accumulation of pre-let-7g and downregulation of mature let-7g with the depletion of EWS. Biochem. Biophys. Res. Commun. 2012, 426, 89–93. [Google Scholar] [CrossRef] [PubMed]
- D’souza, N.; Burns, J.S.; Grisendi, G.; Candini, O.; Veronesi, E.; Piccinno, S.; Horwitz, E.M.; Paolucci, P.; Conte, P.; Dominici, M. MSC and tumors: Homing, differentiation, and secretion influence therapeutic potential. Mesenchymal Stem Cells-Basics Clin. Appl. II 2013, 130, 209–266. [Google Scholar]
- Lin, R.-J.; Lin, Y.-C.; Chen, J.; Kuo, H.-H.; Chen, Y.-Y.; Diccianni, M.B.; London, W.B.; Chang, C.-H.; Yu, A.L. microRNA Signature and Expression of Dicer and Drosha Can Predict Prognosis and Delineate Risk Groups in NeuroblastomamicroRNA, Dicer, and Drosha Expression in Neuroblastoma. Cancer Res. 2010, 70, 7841–7850. [Google Scholar] [CrossRef] [PubMed]
- Merritt, W.M.; Lin, Y.G.; Han, L.Y.; Kamat, A.A.; Spannuth, W.A.; Schmandt, R.; Urbauer, D.; Pennacchio, L.A.; Cheng, J.-F.; Nick, A.M. Dicer, Drosha, and outcomes in patients with ovarian cancer. N. Engl. J. Med. 2008, 359, 2641–2650. [Google Scholar] [CrossRef] [PubMed]
- Ban, J.; Jug, G.; Mestdagh, P.; Schwentner, R.; Kauer, M.; Aryee, D.N.; Schaefer, K.-L.; Nakatani, F.; Scotlandi, K.; Reiter, M. Hsa-mir-145 is the top EWS-FLI1-repressed microRNA involved in a positive feedback loop in Ewing’s sarcoma. Oncogene 2011, 30, 2173–2180. [Google Scholar] [CrossRef]
- Nakatani, F.; Ferracin, M.; Manara, M.C.; Ventura, S.; Del Monaco, V.; Ferrari, S.; Alberghini, M.; Grilli, A.; Knuutila, S.; Schaefer, K.L. miR-34a predicts survival of Ewing’s sarcoma patients and directly influences cell chemo-sensitivity and malignancy. J. Pathol. 2012, 226, 796–805. [Google Scholar] [CrossRef]
- Mosakhani, N.; Guled, M.; Leen, G.; Calabuig-Fariñas, S.; Niini, T.; Machado, I.; Savola, S.; Scotlandi, K.; López-Guerrero, J.A.; Llombart-Bosch, A. An integrated analysis of miRNA and gene copy numbers in xenografts of Ewing’s sarcoma. J. Exp. Clin. Cancer Res. 2012, 31, 24. [Google Scholar] [CrossRef]
- Karnuth, B.; Dedy, N.; Spieker, T.; Lawlor, E.R.; Gattenlöhner, S.; Ranft, A.; Dirksen, U.; Jürgens, H.; Bräuninger, A. Differentially expressed miRNAs in Ewing sarcoma compared to mesenchymal stem cells: Low miR-31 expression with effects on proliferation and invasion. PLoS ONE 2014, 9, e93067. [Google Scholar] [CrossRef]
- Robin, T.P.; Smith, A.; McKinsey, E.; Reaves, L.; Jedlicka, P.; Ford, H.L. EWS/FLI1 Regulates EYA3 in Ewing Sarcoma via Modulation of miRNA-708, Resulting in Increased Cell Survival and ChemoresistanceNovel Mechanisms of Ewing Sarcoma Chemoresistance. Mol. Cancer Res. 2012, 10, 1098–1108. [Google Scholar] [CrossRef] [PubMed]
- Rocchi, A.; Manara, M.C.; Sciandra, M.; Zambelli, D.; Nardi, F.; Nicoletti, G.; Garofalo, C.; Meschini, S.; Astolfi, A.; Colombo, M.P. CD99 inhibits neural differentiation of human Ewing sarcoma cells and thereby contributes to oncogenesis. J. Clin. Investig. 2010, 120, 668–680. [Google Scholar] [CrossRef] [PubMed]
- Lamhamedi-Cherradi, S.-E.; Menegaz, B.A.; Ramamoorthy, V.; Vishwamitra, D.; Wang, Y.; Maywald, R.L.; Buford, A.S.; Fokt, I.; Skora, S.; Wang, J. IGF-1R and mTOR blockade: Novel resistance mechanisms and synergistic drug combinations for Ewing sarcoma. JNCI J. Natl. Cancer Inst. 2016, 108, djw182. [Google Scholar] [CrossRef]
- Aryee, D.N.; Fock, V.; Kapoor, U.; Radic-Sarikas, B.; Kovar, H. Zooming in on long non-coding RNAs in ewing sarcoma pathogenesis. Cells 2022, 11, 1267. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, C.; Sharma, A.R.; Sharma, G.; Lee, S.-S. Therapeutic advances of miRNAs: A preclinical and clinical update. J. Adv. Res. 2021, 28, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Kalariya, N.; Brassil, K.; Calin, G. MicroRNAs: Clinical trials and potential applications. Clin. J. Oncol. Nurs. 2017, 21, 554–560. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Sun, X.; Zhang, Y.; Liu, L.; Yuan, Q. MiR-598: A tumor suppressor with biomarker significance in osteosarcoma. Life Sci. 2017, 188, 141–148. [Google Scholar] [CrossRef]
- Zhao, Y.; Tu, M.-J.; Yu, Y.-F.; Wang, W.-P.; Chen, Q.-X.; Qiu, J.-X.; Yu, A.-X.; Yu, A.-M. Combination therapy with bioengineered miR-34a prodrug and doxorubicin synergistically suppresses osteosarcoma growth. Biochem. Pharmacol. 2015, 98, 602–613. [Google Scholar] [CrossRef] [PubMed]
- Edwards, W.; Dore, S.; van Schalkwyk, J.; Armson, B.A. Prioritizing maternal sepsis: National adoption of an obstetric early warning system to prevent morbidity and mortality. J. Obstet. Gynaecol. Can. 2020, 42, 640–643. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, Q.; Liu, H.; Hu, B.; Zhou, W.; Cheng, Y. MicroRNA expression and its implication for the diagnosis and therapeutic strategies of gastric cancer. Cancer Lett. 2010, 297, 137–143. [Google Scholar] [CrossRef] [PubMed]
- Farazi, T.A.; Spitzer, J.I.; Morozov, P.; Tuschl, T. miRNAs in human cancer. J. Pathol. 2011, 223, 102–115. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| miRs | Target Genes | References |
|---|---|---|
| miR-20b | TGFRB2 | [45] |
| miR-130b | CDC42, ARHGAP1 | [46] |
| miR-34b | Notch1 | [68] |
| miR-708-5p | ROCK1-2, MMP-12 | [9] |
| miR-181c | TNSFRSF6 | [9] |
| miR-193b | ErbB4-C | [9,55] |
| miR-124 | SLUG, CCND2 | [69] |
| miR-30d | MMP-2, MMP-9, Bax, Bcl-2 | [70] |
| miR-107 | HIF-1β | [61] |
| miR-185 | E2F6 | [71] |
| miR-683 | VEGFA | [9] |
| miR-21 | ALCAM | [66] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hassan, M.; Shahzadi, S.; Malik, A.; Din, S.u.; Yasir, M.; Chun, W.; Kloczkowski, A. Oncomeric Profiles of microRNAs as New Therapeutic Targets for Treatment of Ewing’s Sarcoma: A Composite Review. Genes 2023, 14, 1849. https://doi.org/10.3390/genes14101849
Hassan M, Shahzadi S, Malik A, Din Su, Yasir M, Chun W, Kloczkowski A. Oncomeric Profiles of microRNAs as New Therapeutic Targets for Treatment of Ewing’s Sarcoma: A Composite Review. Genes. 2023; 14(10):1849. https://doi.org/10.3390/genes14101849
Chicago/Turabian StyleHassan, Mubashir, Saba Shahzadi, Amal Malik, Salah ud Din, Muhammad Yasir, Wanjoo Chun, and Andrzej Kloczkowski. 2023. "Oncomeric Profiles of microRNAs as New Therapeutic Targets for Treatment of Ewing’s Sarcoma: A Composite Review" Genes 14, no. 10: 1849. https://doi.org/10.3390/genes14101849
APA StyleHassan, M., Shahzadi, S., Malik, A., Din, S. u., Yasir, M., Chun, W., & Kloczkowski, A. (2023). Oncomeric Profiles of microRNAs as New Therapeutic Targets for Treatment of Ewing’s Sarcoma: A Composite Review. Genes, 14(10), 1849. https://doi.org/10.3390/genes14101849