
Large-Scale Sequencing of Borreliaceae for the Construction of Pan-Genomic-Based Diagnostics

 ,
,  , and
, and 
Abstract
1. Introduction
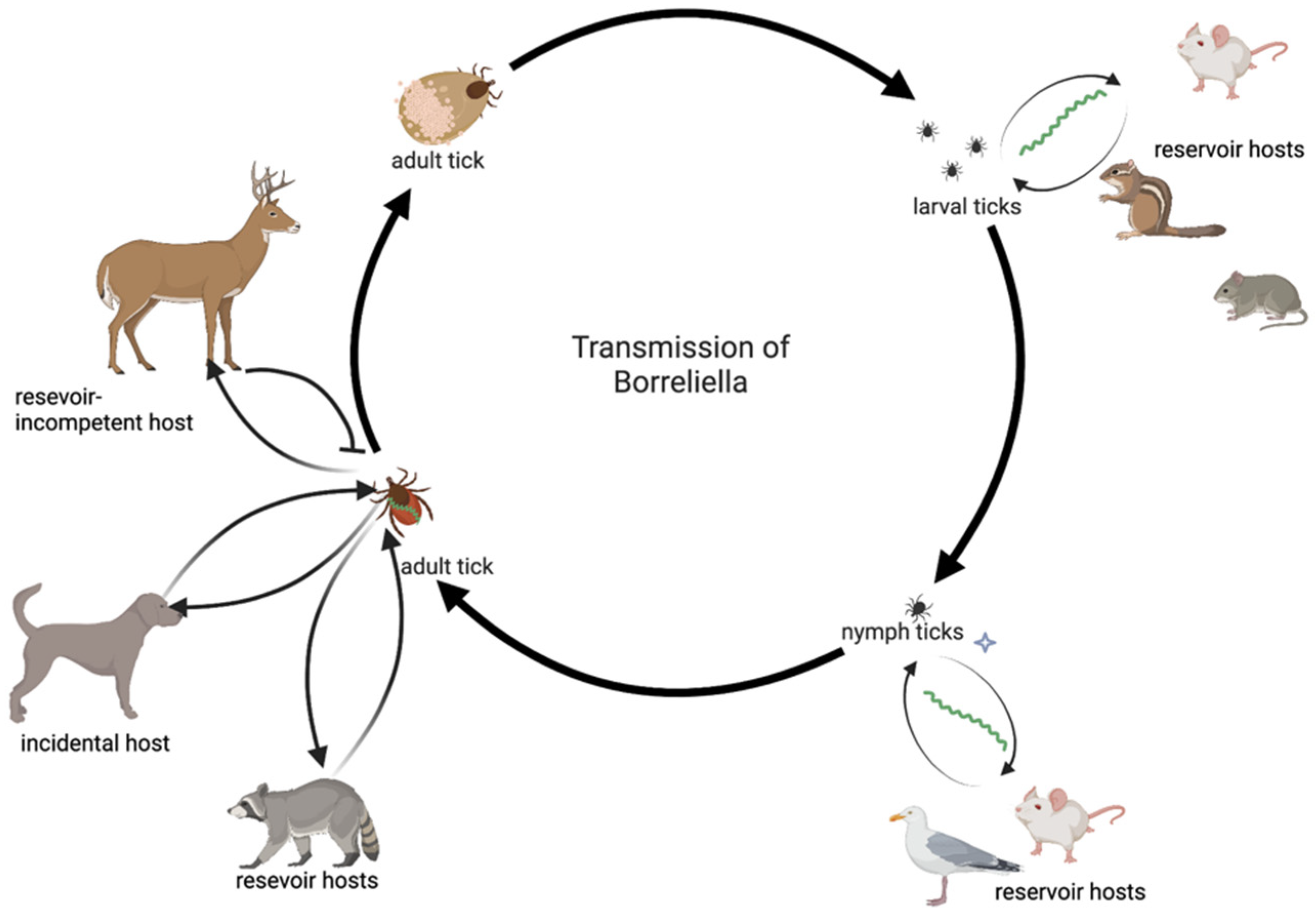
2. The Macro and Micro Ecology of Ticks: Their Role in the Environment and Their Microbiomes
3. Borreliosis, the Most Common Type of Tick-Borne Disease
4. Borreliaceae Diagnostics
5. The Borreliella and Borrelia Genomes
6. Pangenomic Applications
Author Contributions
Funding
Conflicts of Interest
References
- Gubler, D.J. Arthropods in Disease Transmission. In Hunter’s Tropical Medicine and Emerging Infectious Disease; Saunders: Philadelphia, PA, USA, 2013. [Google Scholar]
- Anderson, J.F.; Magnarelli, L.A. Biology of Ticks. Infect. Dis. Clin. North Am. 2008, 22, 195–215. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.F. The Natural History of Ticks. Med. Clin. North Am. 2002, 86, 205–218. [Google Scholar] [CrossRef]
- Obenchain, F.D.; Galun, R. Physiology of Ticks: Current Themes in Tropical Science; Elsevier: Amsterdam, The Netherlands, 2013; ISBN 9781483162348. [Google Scholar]
- Smith, T.; Kilborne, F.L. Investigations into the Nature, Causation, and Prevention of Texas Or Southern Cattle Fever; U.S. Government Printing Office: Washington, DC, USA, 1893.
- Beard, C.B.; Eisen, L.; Eisen, R.J. The Rise of Ticks and Tickborne Diseases in the United States-Introduction. J. Med. Entomol. 2021, 58, 1487–1489. [Google Scholar] [CrossRef]
- Aguero-Rosenfeld, M.E.; Wang, G.; Schwartz, I.; Wormser, G.P. Diagnosis of Lyme Borreliosis. Clin. Microbiol. Rev. 2005, 18, 484–509. [Google Scholar] [CrossRef] [PubMed]
- Mosel, M.R.; Aucott, J.; Schutzer, S.E.; Marques, A.; Arnaboldi, P.M.; Dattwyler, R.; Eshoo, M.W. Lyme Disease Diagnostics. In Lyme Disease and Relapsing Fever Spirochetes: Genomics, Molecular Biology, Host Interactions and Disease Pathogenesis 2021; Caister Academic Press: Wymondham, UK, 2021. [Google Scholar]
- Bowman, A.S.; Nuttall, P.A. Ticks: Biology, Disease and Control; Cambridge University Press: Cambridge, UK, 2008; ISBN 9781107321076. [Google Scholar]
- Entomological Society of America Bloodsucking Ticks (Ixodoidea)—Vectors of Diseases of Man and Animals—Translated From Russia; Entomological Society of America: Annapolis, MD, USA, 1908.
- Khatchikian, C.E.; Prusinski, M.A.; Stone, M.; Backenson, P.B.; Wang, I.-N.; Foley, E.; Seifert, S.N.; Levy, M.Z.; Brisson, D. Recent and Rapid Population Growth and Range Expansion of the Lyme Disease Tick Vector, Ixodes Scapularis, in North America. Evolution 2015, 69, 1678–1689. [Google Scholar] [CrossRef]
- Leighton, P.A.; Koffi, J.K.; Pelcat, Y.; Lindsay, L.R.; Ogden, N.H. Predicting the Speed of Tick Invasion: An Empirical Model of Range Expansion for the Lyme Disease vectorIxodes Scapularisin Canada. J. Appl. Ecol. 2012, 49, 457–464. [Google Scholar] [CrossRef]
- Ogden, N.H.; Maarouf, A.; Barker, I.K.; Bigras-Poulin, M.; Lindsay, L.R.; Morshed, M.G.; O’callaghan, C.J.; Ramay, F.; Waltner-Toews, D.; Charron, D.F. Climate Change and the Potential for Range Expansion of the Lyme Disease Vector Ixodes Scapularis in Canada. Int. J. Parasitol. 2006, 36, 63–70. [Google Scholar] [CrossRef]
- Ogden, N.H.; St-Onge, L.; Barker, I.K.; Brazeau, S.; Bigras-Poulin, M.; Charron, D.F.; Francis, C.M.; Heagy, A.; Lindsay, R.; Maarouf, A.; et al. Risk Maps for Range Expansion of the Lyme Disease Vector, Ixodes Scapularis, in Canada Now and with Climate Change. Int. J. Health Geogr. 2008, 7, 24. [Google Scholar] [CrossRef]
- Gardner, A.M.; Pawlikowski, N.C.; Hamer, S.A.; Hickling, G.J.; Miller, J.R.; Schotthoefer, A.M.; Tsao, J.I.; Allan, B.F. Landscape Features Predict the Current and Forecast the Future Geographic Spread of Lyme Disease. Proc. Biol. Sci. 2020, 287, 20202278. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L.; Beard, C.B. County-Scale Distribution of Ixodes Scapularis and Ixodes Pacificus (Acari: Ixodidae) in the Continental United States. J. Med. Entomol. 2016, 53, 349–386. [Google Scholar] [CrossRef]
- Clover, J.R.; Lane, R.S. Evidence Implicating Nymphal Ixodes Pacificus (Acari: Ixodidae) in the Epidemiology of Lyme Disease in California. Am. J. Trop. Med. Hyg. 1995, 53, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Sonenshine, D.E. Range Expansion of Tick Disease Vectors in North America: Implications for Spread of Tick-Borne Disease. Int. J. Environ. Res. Public Health 2018, 15, 478. [Google Scholar] [CrossRef] [PubMed]
- Hahn, M.B.; Jarnevich, C.S.; Monaghan, A.J.; Eisen, R.J. Modeling the Geographic Distribution of Ixodes Scapularis and Ixodes Pacificus (Acari: Ixodidae) in the Contiguous United States. J. Med. Entomol. 2016, 53, 1176–1191. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Trudel, L.; Artsob, H.; Barker, I.K.; Beauchamp, G.; Charron, D.F.; Drebot, M.A.; Galloway, T.D.; O’Handley, R.; Thompson, R.A.; et al. Ixodes Scapularis Ticks Collected by Passive Surveillance in Canada: Analysis of Geographic Distribution and Infection with Lyme Borreliosis Agent Borrelia Burgdorferi. J. Med. Entomol. 2006, 43, 600–609. [Google Scholar] [CrossRef]
- Descamps, S. Winter Temperature Affects the Prevalence of Ticks in an Arctic Seabird. PLoS ONE 2013, 8, e65374. [Google Scholar]
- Dietrich, M.; Gómez-Díaz, E.; McCoy, K.D. Worldwide Distribution and Diversity of Seabird Ticks: Implications for the Ecology and Epidemiology of Tick-Borne Pathogens. Vector Borne Zoonotic Dis. 2011, 11, 453–470. [Google Scholar] [CrossRef]
- Smith, R.P., Jr.; Muzaffar, S.B.; Lavers, J.; Lacombe, E.H.; Cahill, B.K.; Lubelczyk, C.B.; Kinsler, A.; Mathers, A.J.; Rand, P.W. Borrelia Garinii in Seabird Ticks (Ixodes Uriae), Atlantic Coast, North America. Emerg. Infect. Dis. 2006, 12, 1909–1912. [Google Scholar] [CrossRef]
- Munro, H.J.; Ogden, N.H.; Lindsay, L.R.; Robertson, G.J.; Whitney, H.; Lang, A.S. Evidence for Borrelia Bavariensis Infections of Ixodes Uriae within Seabird Colonies of the North Atlantic Ocean. Appl. Environ. Microbiol. 2017, 83, e01087-17. [Google Scholar] [CrossRef]
- Olsén, B.; Jaenson, T.G.; Noppa, L.; Bunikis, J.; Bergström, S. A Lyme Borreliosis Cycle in Seabirds and Ixodes Uriae Ticks. Nature 1993, 362, 340–342. [Google Scholar] [CrossRef]
- U.S. Centers for Disease Control and Prevention. Others What You Need To Know about Asian Longhorned Ticks-A New Tick in the United States; CDC: Atlanta, GA, USA, 2019. [Google Scholar]
- Nuttall, P. Climate, Ticks and Disease; CABI: Wallingford, UK, 2021; ISBN 9781789249637. [Google Scholar]
- Beard, C.B.; Ben Beard, C.; Occi, J.; Bonilla, D.L.; Egizi, A.M.; Fonseca, D.M.; Mertins, J.W.; Backenson, B.P.; Bajwa, W.I.; Barbarin, A.M.; et al. Multistate Infestation with the Exotic Disease–Vector Tick Haemaphysalis Longicornis—United States, August 2017–September 2018. MMWR. Morbid. Mortal. Wkly Rep. 2018, 67, 1310–1313. [Google Scholar] [CrossRef]
- CDC Regions Where Ticks Live. Available online: https://www.cdc.gov/ticks/geographic_distribution.html (accessed on 9 February 2022).
- Hahn, M.B.; Disler, G.; Durden, L.A.; Coburn, S.; Witmer, F.; George, W.; Beckmen, K.; Gerlach, R. Establishing a Baseline for Tick Surveillance in Alaska: Tick Collection Records from 1909-2019. Ticks Tick Borne Dis. 2020, 11, 101495. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Malawista, S.E.; Snydman, D.R.; Shope, R.E.; Andiman, W.A.; Ross, M.R.; Steele, F.M. Lyme arthritis: An epidemic of oligoarticular arthritis in children and adults in three Connecticut communities. Arthritis Rheum. 1977, 20, 7–17. [Google Scholar] [CrossRef] [PubMed]
- Steere, A.C.; Hardin, J.A.; Malawista, S.E. Lyme Arthritis: A New Clinical Entity. Hosp. Pract. 1978, 13, 143–158. [Google Scholar] [CrossRef] [PubMed]
- Barbour, A.G.; Benach, J.L. Discovery of the Lyme Disease Agent. MBio 2019, 10, e02166-19. [Google Scholar] [CrossRef]
- Anderson, J.F.; Magnarelli, L.A.; Burgdorfer, W.; Barbour, A.G. Spirochetes in Ixodes Dammini and Mammals from Connecticut. Am. J. Trop. Med. Hyg. 1983, 32, 818–824. [Google Scholar] [CrossRef] [PubMed]
- Bosler, E.M.; Coleman, J.L.; Benach, J.L.; Massey, D.A.; Hanrahan, J.P.; Burgdorfer, W.; Barbour, A.G. Natural Distribution of the Ixodes Dammini Spirochete. Science 1983, 220, 321–322. [Google Scholar] [CrossRef]
- Benach, J.L.; Bosler, E.M.; Hanrahan, J.P.; Coleman, J.L.; Habicht, G.S.; Bast, T.F.; Cameron, D.J.; Ziegler, J.L.; Barbour, A.G.; Burgdorfer, W.; et al. Spirochetes Isolated from the Blood of Two Patients with Lyme Disease. N. Engl. J. Med. 1983, 308, 740–742. [Google Scholar] [CrossRef]
- Steere, A.C.; Grodzicki, R.L.; Kornblatt, A.N.; Craft, J.E.; Barbour, A.G.; Burgdorfer, W.; Schmid, G.P.; Johnson, E.; Malawista, S.E. The Spirochetal Etiology of Lyme Disease. N. Engl. J. Med. 1983, 308, 733–740. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Barbour, A.G.; Hayes, S.F.; Benach, J.L.; Grunwaldt, E.; Davis, J.P. Lyme Disease—A Tick-Borne Spirochetosis? Science 1982, 216, 1317–1319. [Google Scholar] [CrossRef]
- Oliver, J.H., Jr.; Owsley, M.R.; Hutcheson, H.J.; James, A.M.; Chen, C.; Irby, W.S.; Dotson, E.M.; McLain, D.K. Conspecificity of the Ticks Ixodes Scapularis and I. Dammini (Acari: Ixodidae). J. Med. Entomol. 1993, 30, 54–63. [Google Scholar] [CrossRef]
- Piesman, J.; Mather, T.N.; Sinsky, R.J.; Spielman, A. Duration of Tick Attachment and Borrelia Burgdorferi Transmission. J. Clin. Microbiol. 1987, 25, 557–558. [Google Scholar] [CrossRef] [PubMed]
- Peavey, C.A.; Lane, R.S. Transmission of Borrelia Burgdorferi by Ixodes Pacificus Nymphs and Reservoir Competence of Deer Mice (Peromyscus Maniculatus) Infected by Tick-Bite. J. Parasitol. 1995, 81, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Lane, R.S.; Brown, R.N.; Piesman, J.; Peavey, C.A. Vector Competence of Ixodes Pacificus and Dermacentor Occidentalis (Acari: Ixodidae) for Various Isolates of Lyme Disease Spirochetes. J. Med. Entomol. 1994, 31, 417–424. [Google Scholar] [CrossRef] [PubMed]
- Piesman, J.; Sinsky, R.J. Ability of Ixodes Scapularis, Dermacentor Variabilis, and Amblyomma Americanum (Acari: Ixodidae) to Acquire, Maintain, and Transmit Lyme Disease Spirochetes (Borrelia Burgdorferi). J. Med. Entomol. 1988, 25, 336–339. [Google Scholar] [CrossRef] [PubMed]
- Eisen, R.J.; Eisen, L. The Blacklegged Tick, Ixodes Scapularis: An Increasing Public Health Concern. Trends Parasitol. 2018, 34, 295–309. [Google Scholar] [CrossRef]
- Hamer, S.A.; Hickling, G.J.; Sidge, J.L.; Walker, E.D.; Tsao, J.I. Synchronous Phenology of Juvenile Ixodes Scapularis, Vertebrate Host Relationships, and Associated Patterns of Borrelia Burgdorferi Ribotypes in the Midwestern United States. Ticks Tick Borne Dis. 2012, 3, 65–74. [Google Scholar] [CrossRef]
- Hunter, D.J.; Torkelson, J.L.; Bodnar, J.; Mortazavi, B.; Laurent, T.; Deason, J.; Thephavongsa, K.; Zhong, J. The Rickettsia Endosymbiont of Ixodes Pacificus Contains All the Genes of De Novo Folate Biosynthesis. PLoS ONE 2015, 10, e0144552. [Google Scholar] [CrossRef]
- Sojka, D.; Franta, Z.; Horn, M.; Caffrey, C.R.; Mareš, M.; Kopáček, P. New Insights into the Machinery of Blood Digestion by Ticks. Trends Parasitol. 2013, 29, 276–285. [Google Scholar] [CrossRef]
- Brown, R.N.; Lane, R.S. Reservoir Competence of Four Chaparral-Dwelling Rodents for Borrelia Burgdorferi in California. Am. J. Trop. Med. Hyg. 1996, 54, 84–91. [Google Scholar] [CrossRef]
- Tracy, K.E.; Baumgarth, N. Borrelia Burgdorferi Manipulates Innate and Adaptive Immunity to Establish Persistence in Rodent Reservoir Hosts. Front. Immunol. 2017, 8, 116. [Google Scholar] [CrossRef]
- Barbour, A.G.; Bunikis, J.; Travinsky, B.; Hoen, A.G.; Diuk-Wasser, M.A.; Fish, D.; Tsao, J.I. Niche Partitioning of Borrelia Burgdorferi and Borrelia Miyamotoi in the Same Tick Vector and Mammalian Reservoir Species. Am. J. Trop. Med. Hyg. 2009, 81, 1120–1131. [Google Scholar] [CrossRef] [PubMed]
- Narasimhan, S.; Fikrig, E. Tick Microbiome: The Force within. Trends Parasitol. 2015, 31, 315–323. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine (US) Committee on Lyme Disease; Other Tick-Borne Diseases: The State of the Science. An Overview of Tick-Borne Diseases; National Academies Press (US): Washington, DC, USA, 2011. [Google Scholar]
- Salkeld, D.J.; Leonhard, S.; Girard, Y.A.; Hahn, N.; Mun, J.; Padgett, K.A.; Lane, R.S. Identifying the Reservoir Hosts of the Lyme Disease Spirochete Borrelia Burgdorferi in California: The Role of the Western Gray Squirrel (Sciurus Griseus). Am. J. Trop. Med. Hyg. 2008, 79, 535–540. [Google Scholar] [CrossRef] [PubMed]
- McKee, C.D.; Bai, Y.; Webb, C.T.; Kosoy, M.Y. Bats Are Key Hosts in the Radiation of Mammal-Associated Bartonella Bacteria. Infect. Genet. Evol. 2021, 89, 104719. [Google Scholar] [CrossRef] [PubMed]
- Hersh, M.H.; Tibbetts, M.; Strauss, M.; Ostfeld, R.S.; Keesing, F. Reservoir Competence of Wildlife Host Species for Babesia Microti. Emerg. Infect. Dis. 2012, 18, 1951–1957. [Google Scholar] [CrossRef]
- Zolnik, C.P.; Makkay, A.M.; Falco, R.C.; Daniels, T.J. American Black Bears as Hosts of Blacklegged Ticks (Acari: Ixodidae) in the Northeastern United States. J. Med. Entomol. 2015, 52, 1103–1110. [Google Scholar] [CrossRef]
- Halsey, S.J.; Allan, B.F.; Miller, J.R. The Role of Ixodes Scapularis, Borrelia Burgdorferi and Wildlife Hosts in Lyme Disease Prevalence: A Quantitative Review. Ticks Tick Borne Dis. 2018, 9, 1103–1114. [Google Scholar] [CrossRef]
- Sidge, J.L.; Foster, E.S.; Buttke, D.E.; Hojgaard, A.; Graham, C.B.; Tsao, J.I. Lake Michigan Insights from Island Studies: The Roles of Chipmunks and Coyotes in Maintaining Ixodes Scapularis and Borrelia Burgdorferi in the Absence of White-Tailed Deer. Ticks Tick Borne Dis. 2021, 12, 101761. [Google Scholar] [CrossRef]
- Lane, R.S.; Loye, J.E. Lyme Disease in California: Interrelationship of Ixodes Pacificus (Acari: Ixodidae), the Western Fence Lizard (Sceloporus Occidentalis), and Borrelia Burgdorferi. J. Med. Entomol. 1989, 26, 272–278. [Google Scholar] [CrossRef]
- Eisen, R.J.; Eisen, L.; Lane, R.S. Habitat-Related Variation in Infestation of Lizards and Rodents with Ixodes Ticks in Dense Woodlands in Mendocino County, California. Exp. Appl. Acarol. 2004, 33, 215–233. [Google Scholar] [CrossRef]
- Casher, L.; Lane, R.; Barrett, R.; Eisen, L. Relative Importance of Lizards and Mammals as Hosts for Ixodid Ticks in Northern California. Exp. Appl. Acarol. 2002, 26, 127. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Raoult, D. Ticks and Tickborne Bacterial Diseases in Humans: An Emerging Infectious Threat. Clin. Infect. Dis. 2001, 32, 897–928. [Google Scholar] [CrossRef]
- Yabsley, M.J.; Shock, B.C. Natural History of Zoonotic Babesia: Role of Wildlife Reservoirs. Int. J. Parasitol. Parasites Wildl. 2013, 2, 18–31. [Google Scholar] [CrossRef]
- Woldehiwet, Z. The Natural History of Anaplasma Phagocytophilum. Vet. Parasitol. 2010, 167, 108–122. [Google Scholar] [CrossRef] [PubMed]
- Fukunaga, M.; Takahashi, Y.; Tsuruta, Y.; Matsushita, O.; Ralph, D.; McClelland, M.; Nakao, M. Genetic and Phenotypic Analysis of Borrelia Miyamotoi Sp. Nov., Isolated from the Ixodid Tick Ixodes Persulcatus, the Vector for Lyme Disease in Japan. Int. J. Syst. Bacteriol. 1995, 45, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Kingry, L.C.; Batra, D.; Replogle, A.; Rowe, L.A.; Pritt, B.S.; Petersen, J.M. Whole Genome Sequence and Comparative Genomics of the Novel Lyme Borreliosis Causing Pathogen, Borrelia Mayonii. PLoS ONE 2016, 11, e0168994. [Google Scholar] [CrossRef] [PubMed]
- Paddock, C.D.; Childs, J.E. Ehrlichia Chaffeensis: A Prototypical Emerging Pathogen. Clin. Microbiol. Rev. 2003, 16, 37–64. [Google Scholar] [CrossRef]
- Pritt, B.S.; Sloan, L.M.; Johnson, D.K.H.; Munderloh, U.G.; Paskewitz, S.M.; McElroy, K.M.; McFadden, J.D.; Binnicker, M.J.; Neitzel, D.F.; Liu, G.; et al. Emergence of a New Pathogenic Ehrlichia Species, Wisconsin and Minnesota, 2009. N. Engl. J. Med. 2011, 365, 422–429. [Google Scholar] [CrossRef]
- McCoy, G.W. Chapin Bacterium Tularense, the Cause of a Plague-like Disease of Rodents. J. Infect. Dis. (Public Health Bull.) 1912, 10, 61–72. [Google Scholar]
- Paddock, C.D.; Sumner, J.W.; Comer, J.A.; Zaki, S.R.; Goldsmith, C.S.; Goddard, J.; McLellan, S.L.F.; Tamminga, C.L.; Ohl, C.A. Rickettsia Parkeri: A Newly Recognized Cause of Spotted Fever Rickettsiosis in the United States. Clin. Infect. Dis. 2004, 38, 805–811. [Google Scholar] [CrossRef]
- Kosoy, O.I.; Lambert, A.J.; Hawkinson, D.J.; Pastula, D.M.; Goldsmith, C.S.; Charles Hunt, D.; Erin Staples, J. Novel Thogotovirus Associated with Febrile Illness and Death, United States, 2014. Emerg. Infect. Dis. 2015, 21, 760–764. [Google Scholar] [CrossRef] [PubMed]
- Florio, F.; Miller, M.S.; Mugrage, E.R. Colorado Tick Fever: Isolation of the Virus from Dermacentor Andersoni in Nature and a Laboratory Study of the Transmission of the Virus in the Tick. J. Immunol. 1950, 64, 257–263. [Google Scholar] [PubMed]
- McMullan, L.K.; Folk, S.M.; Kelly, A.J.; MacNeil, A.; Goldsmith, C.S.; Metcalfe, M.G.; Batten, B.C.; Albariño, C.G.; Zaki, S.R.; Rollin, P.E.; et al. A New Phlebovirus Associated with Severe Febrile Illness in Missouri. N. Engl. J. Med. 2012, 367, 834–841. [Google Scholar] [CrossRef] [PubMed]
- McLEAN, D.M.; Donohue, W.L. Powassan Virus: Isolation of Virus from a Fatal Case of Encephalitis. Can. Med. Assoc. J. 1959, 80, 708–711. [Google Scholar] [PubMed]
- Cutler, S.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A New Borrelia on the Block: Borrelia Miyamotoi—A Human Health Risk? Euro. Surveill. 2019, 24, 180017. [Google Scholar] [CrossRef]
- Johnson, T.L.; Graham, C.B.; Hojgaard, A.; Breuner, N.E.; Maes, S.E.; Boegler, K.A.; Replogle, A.J.; Kingry, L.C.; Petersen, J.M.; Eisen, L.; et al. Isolation of the Lyme Disease Spirochete Borrelia Mayonii From Naturally Infected Rodents in Minnesota. J. Med. Entomol. 2017, 54, 1088–1092. [Google Scholar] [CrossRef]
- Rochlin, I.; Egizi, A.; Lindström, A. The Original Scientific Description of the Lone Star Tick (Amblyomma Americanum, Acari: Ixodidae) and Implications for the Species’ Past and Future Geographic Distributions. J. Med. Entomol. 2022, 59, 412–420. [Google Scholar] [CrossRef]
- Stromdahl, E.Y.; Nadolny, R.M.; Hickling, G.J.; Hamer, S.A.; Ogden, N.H.; Casal, C.; Heck, G.A.; Gibbons, J.A.; Cremeans, T.F.; Pilgard, M.A. Amblyomma Americanum (Acari: Ixodidae) Ticks Are Not Vectors of the Lyme Disease Agent, Borrelia Burgdorferi (Spirocheatales: Spirochaetaceae): A Review of the Evidence. J. Med. Entomol. 2018, 55, 501–514. [Google Scholar] [CrossRef]
- Burgdorfer, W.; Lane, R.S.; Barbour, A.G.; Gresbrink, R.A.; Anderson, J.R. The Western Black-Legged Tick, Ixodes Pacificus: A Vector of Borrelia Burgdorferi. Am. J. Trop. Med. Hyg. 1985, 34, 925–930. [Google Scholar] [CrossRef]
- Padgett, K.A.; Lane, R.S. Life Cycle of Ixodes Pacificus (Acari: Ixodidae): Timing of Developmental Processes under Field and Laboratory Conditions. J. Med. Entomol. 2001, 38, 684–693. [Google Scholar] [CrossRef]
- Chang, C.C.; Chomel, B.B.; Kasten, R.W.; Romano, V.; Tietze, N. Molecular Evidence of Bartonella Spp. in Questing Adult Ixodes Pacificus Ticks in California. J. Clin. Microbiol. 2001, 39, 1221–1226. [Google Scholar] [CrossRef]
- Dingler, R.J.; Wright, S.A.; Donohue, A.M.; Macedo, P.A.; Foley, J.E. Surveillance for Ixodes Pacificus and the Tick-Borne Pathogens Anaplasma Phagocytophilum and Borrelia Burgdorferi in Birds from California’s Inner Coast Range. Ticks Tick Borne Dis. 2014, 5, 436–445. [Google Scholar] [CrossRef] [PubMed]
- Kwan, J.Y.; Griggs, R.; Chicana, B.; Miller, C.; Swei, A. Vertical vs. Horizontal Transmission of the Microbiome in a Key Disease Vector, Ixodes Pacificus. Mol. Ecol. 2017, 26, 6578–6589. [Google Scholar] [CrossRef]
- Loye, J.E.; Lane, R.S. Questing Behavior of Ixodes Pacificus (Acari: Ixodidae) in Relation to Meteorological and Seasonal Factors. J. Med. Entomol. 1988, 25, 391–398. [Google Scholar] [CrossRef] [PubMed]
- Lederberg, J.; McCray, A.T. Ome SweetOmics—A Genealogical Treasury of Words. Scientist 2001, 15, 8. [Google Scholar]
- Jansson, J.K.; Hofmockel, K.S. The Soil Microbiome—From Metagenomics to Metaphenomics. Curr. Opin. Microbiol. 2018, 43, 162–168. [Google Scholar] [CrossRef]
- Sunagawa, S.; Coelho, L.P.; Chaffron, S.; Kultima, J.R.; Labadie, K.; Salazar, G.; Djahanschiri, B.; Zeller, G.; Mende, D.R.; Alberti, A.; et al. Ocean Plankton. Structure and Function of the Global Ocean Microbiome. Science 2015, 348, 1261359. [Google Scholar] [CrossRef]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The Human Microbiome Project. Nature 2007, 449, 804–810. [Google Scholar] [CrossRef]
- Ehrlich, G.D.; Hiller, N.L.; Hu, F.Z. What Makes Pathogens Pathogenic. Genome Biol. 2008, 9, 225. [Google Scholar] [CrossRef]
- Bonnet, S.I.; Binetruy, F.; Hernández-Jarguín, A.M.; Duron, O. The Tick Microbiome: Why Non-Pathogenic Microorganisms Matter in Tick Biology and Pathogen Transmission. Front. Cell. Infect. Microbiol. 2017, 7, 236. [Google Scholar] [CrossRef]
- Estrada-Peña, A.; Cabezas-Cruz, A.; Obregón, D. Behind Taxonomic Variability: The Functional Redundancy in the Tick Microbiome. Microorganisms 2020, 8, 1829. [Google Scholar] [CrossRef] [PubMed]
- Varela-Stokes, A.S.; Park, S.H.; Kim, S.A.; Ricke, S.C. Microbial Communities in North American Ixodid Ticks of Veterinary and Medical Importance. Front. Vet. Sci. 2017, 4, 179. [Google Scholar] [CrossRef]
- Zhong, J.; Jasinskas, A.; Barbour, A.G. Antibiotic Treatment of the Tick Vector Amblyomma Americanum Reduced Reproductive Fitness. PLoS ONE 2007, 2, e405. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Perveen, N.; Hussain, A.; Song, B.; Aziz, M.U.; Zeb, J.; Li, J.; George, D.; Cabezas-Cruz, A.; Sparagano, O. The Symbiotic Continuum Within Ticks: Opportunities for Disease Control. Front. Microbiol. 2022, 13, 854803. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Zhong, T.; Peng, Y.; Zhou, X.; Wang, Z.; Tang, H.; Wang, J. Symbiont-Regulated Serotonin Biosynthesis Modulates Tick Feeding Activity. Cell Host Microbe 2021, 29, 1545–1557.e4. [Google Scholar] [CrossRef]
- Ross, B.D.; Hayes, B.; Radey, M.C.; Lee, X.; Josek, T.; Bjork, J.; Neitzel, D.; Paskewitz, S.; Chou, S.; Mougous, J. Ixodes Scapularis Does Not Harbor a Stable Midgut Microbiome. ISME J. 2018, 12, 2596–2607. [Google Scholar] [CrossRef]
- Zolnik, C.P.; Falco, R.C.; Daniels, T.J.; Kolokotronis, S.-O. Transient Influence of Blood Meal and Natural Environment on Blacklegged Tick Bacterial Communities. Ticks Tick Borne Dis. 2018, 9, 563–572. [Google Scholar] [CrossRef]
- Thapa, S.; Zhang, Y.; Allen, M.S. Bacterial Microbiomes of Ixodes Scapularis Ticks Collected from Massachusetts and Texas, USA. BMC Microbiol. 2019, 19, 138. [Google Scholar] [CrossRef]
- Obregón, D.; Bard, E.; Abrial, D.; Estrada-Peña, A.; Cabezas-Cruz, A. Sex-Specific Linkages Between Taxonomic and Functional Profiles of Tick Gut Microbiomes. Front. Cell. Infect. Microbiol. 2019, 9, 298. [Google Scholar] [CrossRef]
- Travanty, N.V.; Ponnusamy, L.; Kakumanu, M.L.; Nicholson, W.L.; Apperson, C.S. Diversity and Structure of the Bacterial Microbiome of the American Dog Tick, Dermacentor Variabilis, Is Dominated by the Endosymbiont Francisella. Symbiosis 2019, 79, 239–250. [Google Scholar] [CrossRef]
- Brinkerhoff, R.J.; Clark, C.; Ocasio, K.; Gauthier, D.T.; Hynes, W.L. Factors Affecting the Microbiome of Ixodes Scapularis and Amblyomma Americanum. PLoS ONE 2020, 15, e0232398. [Google Scholar] [CrossRef] [PubMed]
- Chicana, B.; Couper, L.I.; Kwan, J.Y.; Tahiraj, E.; Swei, A. Comparative Microbiome Profiles of Sympatric Tick Species from the Far-Western United States. Insects 2019, 10, 353. [Google Scholar] [CrossRef] [PubMed]
- Wu-Chuang, A.; Hodžić, A.; Mateos-Hernández, L.; Estrada-Peña, A.; Obregon, D.; Cabezas-Cruz, A. Current Debates and Advances in Tick Microbiome Research. Curr Res Parasitol Vector Borne Dis 2021, 1, 100036. [Google Scholar] [CrossRef] [PubMed]
- Williams-Newkirk, A.J.; Rowe, L.A.; Mixson-Hayden, T.R.; Dasch, G.A. Characterization of the Bacterial Communities of Life Stages of Free Living Lone Star Ticks (Amblyomma Americanum). PLoS ONE 2014, 9, e102130. [Google Scholar]
- Zhang, R.; Yu, G.; Huang, Z.; Zhang, Z. Microbiota Assessment across Different Developmental Stages of Dermacentor Silvarum (Acari: Ixodidae) Revealed Stage-Specific Signatures. Ticks Tick Borne Dis. 2020, 11, 101321. [Google Scholar] [CrossRef]
- Eshoo, M.W.; Carolan, H.E.; Massire, C.; Chou, D.M.; Crowder, C.D.; Rounds, M.A.; Phillipson, C.A.; Schutzer, S.E.; Ecker, D.J. Survey of Ixodes Pacificus Ticks in California Reveals a Diversity of Microorganisms and a Novel and Widespread Anaplasmataceae Species. PLoS ONE 2015, 10, e0135828. [Google Scholar] [CrossRef]
- Afzelius, A. Verhandlungen Der Dermatologischen Gesellschaft Zu Stockholm. Arch. Dermatol. Syph. 1910, 101, 404. [Google Scholar]
- Hellerstrom, S. Erythema Chronicum Migrans Afzelius with Meningitis. Acta Derm. Venereol. 1951, 31, 227–234. [Google Scholar] [CrossRef]
- Garin, C.H.; Bujadoux, C.H. Paralysie Par Les Tiques. J. Med. Lyon 1922, 71, 7. [Google Scholar]
- Benach, J.; Garcia Monco, J. The Woldwide Saga of Lyme Borreliosis. Borrelia. Molecular Biology, Host Interaction and Pathogenesis; Caister Academic Press: Norwich, UK, 2010; pp. 7–26. [Google Scholar]
- Steere, A.C.; Malawista, S.E.; Hardin, J.A.; Ruddy, S.; Askenase, W.; Andiman, W.A. Erythema Chronicum Migrans and Lyme Arthritis. The Enlarging Clinical Spectrum. Ann. Intern. Med. 1977, 86, 685–698. [Google Scholar] [CrossRef]
- Reik, L.; Steere, A.C.; Bartenhagen, N.H.; Shope, R.E.; Malawista, S.E. Neurologic Abnormalities of Lyme Disease. Medicine 1979, 58, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Walter, K.S.; Carpi, G.; Caccone, A.; Diuk-Wasser, M.A. Genomic Insights into the Ancient Spread of Lyme Disease across North America. Nat Ecol Evol 2017, 1, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Earnhart, C.G.; Marconi, R.T. Chapter 52—Lyme Disease. In Vaccines for Biodefense and Emerging and Neglected Diseases; Barrett, A.D.T., Stanberry, L.R., Eds.; Academic Press: London, UK, 2009; pp. 1031–1060. ISBN 9780123694089. [Google Scholar]
- Data and Surveillance Lyme Disease|CDC. Available online: https://www.cdc.gov/lyme/datasurveillance/index.html (accessed on 25 April 2019).
- Dworkin, M.S.; Schwan, T.G.; Anderson, D.E., Jr.; Borchardt, S.M. Tick-Borne Relapsing Fever. Infect. Dis. Clin. North Am. 2008, 22, 449–468. [Google Scholar] [CrossRef] [PubMed]
- Pritt, B.S.; Mead, P.S.; Johnson, D.K.H.; Neitzel, D.F.; Respicio-Kingry, L.B.; Davis, J.P.; Schiffman, E.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; et al. Identification of a Novel Pathogenic Borrelia Species Causing Lyme Borreliosis with Unusually High Spirochaetaemia: A Descriptive Study. Lancet Infect. Dis. 2016, 16, 556–564. [Google Scholar] [CrossRef]
- Crowder, C.D.; Matthews, H.E.; Schutzer, S.; Rounds, M.A.; Luft, B.J.; Nolte, O.; Campbell, S.R.; Phillipson, C.A.; Li, F.; Sampath, R.; et al. Genotypic Variation and Mixtures of Lyme Borrelia in Ixodes Ticks from North America and Europe. PLoS ONE 2010, 5, e10650. [Google Scholar] [CrossRef]
- Margos, G.; Vollmer, S.A.; Ogden, N.H.; Fish, D. Population Genetics, Taxonomy, Phylogeny and Evolution of Borrelia Burgdorferi Sensu Lato. Infect. Genet. Evol. 2011, 11, 1545–1563. [Google Scholar] [CrossRef]
- Ras, N.M.; Lascola, B.; Postic, D.; Cutler, S.J.; Rodhain, F.; Baranton, G.; Raoult, D. Phylogenesis of Relapsing Fever Borrelia spp. Int. J. Syst. Bacteriol. 1996, 46, 859–865. [Google Scholar]
- Adeolu, M.; Gupta, R.S. A Phylogenomic and Molecular Marker Based Proposal for the Division of the Genus Borrelia into Two Genera: The Emended Genus Borrelia Containing Only the Members of the Relapsing Fever Borrelia, and the Genus Borreliella Gen. Nov. Containing the Members of the Lyme Disease Borrelia (Borrelia Burgdorferi Sensu Lato Complex). Antonie Van Leeuwenhoek 2014, 105, 1049–1072. [Google Scholar]
- Müllegger, R.R.; Glatz, M. Skin Manifestations of Lyme Borreliosis. Am. J. Clin. Dermatol. 2008, 9, 355–368. [Google Scholar] [CrossRef]
- Verma, V.; Roman, M.; Shah, D.; Zaretskaya, M.; Yassin, M.H. A Case of Chronic Progressive Lyme Encephalitis as a Manifestation of Late Lyme Neuroborreliosis. Infect. Dis. Rep. 2014, 6, 5496. [Google Scholar] [CrossRef]
- Brodziński, S.; Nasierowski, T. Psychosis in Borrelia Burgdorferi Infection—Part I: Epidemiology, Pathogenesis, Diagnosis and Treatment of Neuroborreliosis. Psychiatr. Pol. 2019, 53, 629–640. [Google Scholar] [CrossRef] [PubMed]
- Arav-Boger, R.; Crawford, T.; Steere, A.C.; Halsey, N.A. Cerebellar Ataxia as the Presenting Manifestation of Lyme Disease. Pediatr. Infect. Dis. J. 2002, 21, 353–356. [Google Scholar] [CrossRef] [PubMed]
- Rauer, S.; Kastenbauer, S.; Fingerle, V.; Hunfeld, K.-P.; Huppertz, H.-I.; Dersch, R. Lyme Neuroborreliosis. Dtsch. Arztebl. Int. 2018, 115, 751–756. [Google Scholar] [CrossRef] [PubMed]
- Cardenas-de la Garza, J.A.; De la Cruz-Valadez, E.; Ocampo-Candiani, J.; Welsh, O. Clinical Spectrum of Lyme Disease. Eur. J. Clin. Microbiol. Infect. Dis. 2019, 38, 201–208. [Google Scholar] [CrossRef]
- Cassarino, D.S.; Quezado, M.M.; Ghatak, N.R.; Duray, P.H. Lyme-Associated Parkinsonism: A Neuropathologic Case Study and Review of the Literature. Arch. Pathol. Lab. Med. 2003, 127, 1204–1206. [Google Scholar] [CrossRef]
- Stanek, G.; Wormser, G.P.; Gray, J.; Strle, F. Lyme Borreliosis. Lancet 2012, 379, 461–473. [Google Scholar] [CrossRef]
- Blanc, F.; Philippi, N.; Cretin, B.; Kleitz, C.; Berly, L.; Jung, B.; Kremer, S.; Namer, I.J.; Sellal, F.; Jaulhac, B.; et al. Lyme Neuroborreliosis and Dementia. J. Alzheimers. Dis. 2014, 41, 1087–1093. [Google Scholar] [CrossRef]
- Steere, A.C.; Malawista, S.E.; Bartenhagen, N.H.; Spieler, P.N.; Newman, J.H.; Rahn, D.W.; Hutchinson, G.J.; Green, J.; Snydman, D.R.; Taylor, E. The Clinical Spectrum and Treatment of Lyme Disease. Yale J. Biol. Med. 1984, 57, 453–461. [Google Scholar]
- Fix, A.D.; Peña, C.A.; Strickland, G.T. Racial Differences in Reported Lyme Disease Incidence. Am. J. Epidemiol. 2000, 152, 756–759. [Google Scholar] [CrossRef]
- Coburn, J.; Garcia, B.; Hu, L.T.; Jewett, M.W.; Kraiczy, P.; Norris, S.J.; Skare, J. Lyme Disease Pathogenesis. Curr. Issues Mol. Biol. 2020, 42, 473–518. [Google Scholar]
- Torbahn, G.; Hofmann, H.; Rücker, G.; Bischoff, K.; Freitag, M.H.; Dersch, R.; Fingerle, V.; Motschall, E.; Meerpohl, J.J.; Schmucker, C. Efficacy and Safety of Antibiotic Therapy in Early Cutaneous Lyme Borreliosis: A Network Meta-Analysis. JAMA Dermatol. 2018, 154, 1292–1303. [Google Scholar] [CrossRef] [PubMed]
- Tilly, K.; Rosa, P.A.; Stewart, P.E. Biology of Infection with Borrelia Burgdorferi. Infect. Dis. Clin. North Am. 2008, 22, 217–234. [Google Scholar] [CrossRef] [PubMed]
- DeHart, T.G.; Kushelman, M.R.; Hildreth, S.B.; Helm, R.F.; Jutras, B.L. The Unusual Cell Wall of the Lyme Disease Spirochaete Borrelia Burgdorferi Is Shaped by a Tick Sugar. Nat. Microbiol. 2021, 6, 1583–1592. [Google Scholar] [CrossRef] [PubMed]
- Motaleb, M.A.; Corum, L.; Bono, J.L.; Elias, A.F.; Rosa, P.; Samuels, D.S.; Charon, N.W. Borrelia Burgdorferi Periplasmic Flagella Have Both Skeletal and Motility Functions. Proc. Natl. Acad. Sci. USA 2000, 97, 10899–10904. [Google Scholar] [CrossRef] [PubMed]
- Sapi, E.; Bastian, S.L.; Mpoy, C.M.; Scott, S.; Rattelle, A.; Pabbati, N.; Poruri, A.; Burugu, D.; Theophilus, P.A.S.; Pham, T.V.; et al. Characterization of Biofilm Formation by Borrelia Burgdorferi in Vitro. PLoS ONE 2012, 7, e48277. [Google Scholar] [CrossRef]
- Sapi, E.; Balasubramanian, K.; Poruri, A.; Maghsoudlou, J.S.; Socarras, K.M.; Timmaraju, A.V.; Filush, K.R.; Gupta, K.; Shaikh, S.; Theophilus, P.A.S.; et al. Evidence of In Vivo Existence of Borrelia Biofilm in Borrelial Lymphocytomas. Eur. J. Microbiol. Immunol. 2016, 6, 9–24. [Google Scholar] [CrossRef]
- Timmaraju, V.A.; Theophilus, P.A.S.; Balasubramanian, K.; Shakih, S.; Luecke, D.F.; Sapi, E. Biofilm Formation by Borrelia Burgdorferi Sensu Lato. FEMS Microbiol. Lett. 2015, 362, fnv120. [Google Scholar] [CrossRef][Green Version]
- Shaikh, S.; Timmaraju, V.A.; Torres, J.P.; Socarras, K.M.; Theophilus, P.A.S.; Sapi, E. Influence of Tick and Mammalian Physiological Temperatures on Borrelia Burgdorferi Biofilms. Microbiology 2016, 162, 1984–1995. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Kybicova, K.; Vancova, M. Metamorphoses of Lyme Disease Spirochetes: Phenomenon of Borrelia Persisters. Parasit. Vectors 2019, 12, 237. [Google Scholar] [CrossRef]
- Pal, U.; Li, X.; Wang, T.; Montgomery, R.R.; Ramamoorthi, N.; Desilva, A.M.; Bao, F.; Yang, X.; Pypaert, M.; Pradhan, D.; et al. TROSPA, an Ixodes Scapularis Receptor for Borrelia Burgdorferi. Cell 2004, 119, 457–468. [Google Scholar] [CrossRef]
- Kurokawa, C.; Lynn, G.E.; Pedra, J.H.F.; Pal, U.; Narasimhan, S.; Fikrig, E. Interactions between Borrelia Burgdorferi and Ticks. Nat. Rev. Microbiol. 2020, 18, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Hovius, J.W.R.; van Dam, A.P.; Fikrig, E. Tick–host–pathogen Interactions in Lyme Borreliosis. Trends Parasitol. 2007, 23, 434–438. [Google Scholar] [CrossRef] [PubMed]
- CDC Transmission. Available online: https://www.cdc.gov/lyme/transmission/index.html (accessed on 17 March 2022).
- Eisen, L. Pathogen Transmission in Relation to Duration of Attachment by Ixodes Scapularis Ticks. Ticks Tick Borne Dis. 2018, 9, 535–542. [Google Scholar] [CrossRef] [PubMed]
- Cook, M.J. Lyme Borreliosis: A Review of Data on Transmission Time after Tick Attachment. Int. J. Gen. Med. 2015, 8, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Hyde, J.A. Borrelia Burgdorferi Keeps Moving and Carries on: A Review of Borrelial Dissemination and Invasion. Front. Immunol. 2017, 8, 114. [Google Scholar] [CrossRef]
- Oosting, M.; Buffen, K.; van der Meer, J.W.M.; Netea, M.G.; Joosten, L.A.B. Innate Immunity Networks during Infection with Borrelia Burgdorferi. Crit. Rev. Microbiol. 2016, 42, 233–244. [Google Scholar]
- Bockenstedt, L.K.; Wooten, R.M.; Baumgarth, N. Immune response to Borrelia: Lessons from Lyme disease spirochetes. Curr. Issues Mol. Biol. 2021, 42, 145–190. [Google Scholar]
- Elsner, R.A.; Barthold, S.W.; Baumgarth, N. Delays and Diversions Mark the Development of B Cell Responses to Borrelia Burgdorferi Infection. J. Immunol. 2012, 188, 5612–5622. [Google Scholar]
- Elsner, R.A.; Hastey, C.J.; Baumgarth, N. CD4+ T Cells Promote Antibody Production but Not Sustained Affinity Maturation during Borrelia Burgdorferi Infection. Infect. Immun. 2015, 83, 48–56. [Google Scholar] [CrossRef]
- Röttgerding, F.; Kraiczy, P. Immune Evasion Strategies of Relapsing Fever Spirochetes. Front. Immunol. 2020, 11, 1560. [Google Scholar] [CrossRef]
- Hallström, T.; Siegel, C.; Mörgelin, M.; Kraiczy, P.; Skerka, C.; Zipfel, P.F. CspA from Borrelia Burgdorferi Inhibits the Terminal Complement Pathway. MBio 2013, 4, e00481-13. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.; Nguyen, N.T.T.; Nowak, N.A.; Zhang, F.; Linhardt, R.J.; Diuk-Wasser, M.; Ram, S.; Kraiczy, P.; Lin, Y.-P. Polymorphic Factor H-Binding Activity of CspA Protects Lyme Borreliae from the Host Complement in Feeding Ticks to Facilitate Tick-to-Host Transmission. PLoS Pathog. 2018, 14, e1007106. [Google Scholar] [CrossRef]
- Lin, Y.-P.; Frye, A.M.; Nowak, T.A.; Kraiczy, P. New Insights Into CRASP-Mediated Complement Evasion in the Lyme Disease Enzootic Cycle. Front. Cell. Infect. Microbiol. 2020, 10, 1. [Google Scholar] [CrossRef] [PubMed]
- Garcia, B.L.; Zhi, H.; Wager, B.; Höök, M.; Skare, J.T. Borrelia Burgdorferi BBK32 Inhibits the Classical Pathway by Blocking Activation of the C1 Complement Complex. PLoS Pathog. 2016, 12, e1005404. [Google Scholar] [CrossRef] [PubMed]
- Kraiczy, P. Hide and Seek: How Lyme Disease Spirochetes Overcome Complement Attack. Front. Immunol. 2016, 7, 385. [Google Scholar] [CrossRef] [PubMed]
- Locke, J.W. Complement Evasion in Borrelia Spirochetes: Mechanisms and Opportunities for Intervention. Antibiotics 2019, 8, 80. [Google Scholar] [CrossRef]
- Koenigs, A.; Hammerschmidt, C.; Jutras, B.L.; Pogoryelov, D.; Barthel, D.; Skerka, C.; Kugelstadt, D.; Wallich, R.; Stevenson, B.; Zipfel, P.F.; et al. BBA70 of Borrelia Burgdorferi Is a Novel Plasminogen-Binding Protein. J. Biol. Chem. 2013, 288, 25229–25243. [Google Scholar] [CrossRef]
- Talagrand-Reboul, E.; Boyer, P.H.; Bergström, S.; Vial, L.; Boulanger, N. Relapsing Fevers: Neglected Tick-Borne Diseases. Front. Cell. Infect. Microbiol. 2018, 8, 98. [Google Scholar] [CrossRef]
- Cutler, S.J. Relapsing Fever—A Forgotten Disease Revealed. J. Appl. Microbiol. 2010, 108, 1115–1122. [Google Scholar] [CrossRef]
- Forrester, J.D.; Kjemtrup, A.M.; Fritz, C.L.; Marsden-Haug, N.; Nichols, J.B.; Tengelsen, L.A.; Sowadsky, R.; DeBess, E.; Cieslak, P.R.; Weiss, J.; et al. Tickborne Relapsing Fever—United States, 1990–2011. MMWR Morb. Mortal. Wkly. Rep. 2015, 64, 58–60. [Google Scholar]
- CDC Symptoms. Available online: https://www.cdc.gov/relapsing-fever/symptoms/index.html (accessed on 10 February 2022).
- Lopez, J.; Hovius, J.W.; Bergström, S. Pathogenesis of Relapsing Fever. In Lyme Disease and Relapsing Fever Spirochetes: Genomics, Molecular Biology, Host Interactions and Disease Pathogenesis 2021; Caister Academic Press: Wymondham, UK, 2021. [Google Scholar]
- Adrion, E.R.; Aucott, J.; Lemke, K.W.; Weiner, J.P. Health Care Costs, Utilization and Patterns of Care Following Lyme Disease. PLoS ONE 2015, 10, e0116767. [Google Scholar]
- Pfoh, E.; Wessels, M.R.; Goldmann, D.; Lee, G.M. Burden and Economic Cost of Group A Streptococcal Pharyngitis. Pediatrics 2008, 121, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Xie, Y.; Xiao, Y. Laboratory Diagnostic Tools for Syphilis: Current Status and Future Prospects. Front. Cell. Infect. Microbiol. 2020, 10, 574806. [Google Scholar] [CrossRef] [PubMed]
- Park, I.U.; Fakile, Y.F.; Chow, J.M.; Gustafson, K.J.; Jost, H.; Schapiro, J.M.; Novak-Weekley, S.; Tran, A.; Nomura, J.H.; Chen, V.; et al. Performance of Treponemal Tests for the Diagnosis of Syphilis. Clin. Infect. Dis. 2019, 68, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Worrall, G.; Hutchinson, J.; Sherman, G.; Griffiths, J. Diagnosing Streptococcal Sore Throat in Adults: Randomized Controlled Trial of in-Office Aids. Can. Fam. Phys. 2007, 53, 666–671. [Google Scholar]
- Tansarli, G.S.; LeBlanc, L.; Auld, D.B.; Chapin, K.C. Diagnostic Accuracy of Presurgical Staphylococcus Aureus PCR Assay Compared with Culture and Post-PCR Implementation Surgical Site Infection Rates. J. Mol. Diagn. 2020, 22, 1063–1069. [Google Scholar] [CrossRef]
- Schmidt, B.L. PCR in Laboratory Diagnosis of Human Borrelia Burgdorferi Infections. Clin. Microbiol. Rev. 1997, 10, 185–201. [Google Scholar] [CrossRef]
- Aucott, J.; Morrison, C.; Munoz, B.; Rowe, P.C.; Schwarzwalder, A.; West, S.K. Diagnostic Challenges of Early Lyme Disease: Lessons from a Community Case Series. BMC Infect. Dis. 2009, 9, 79. [Google Scholar] [CrossRef]
- Barrett, J. It’s about Lyme: Why Congress Must Enact Medical Insurance Coverage Laws for Lyme Disease Patients Now. Seattle U. L. Rev. SUpra 2022, 45, 50. [Google Scholar]
- St Pierre, S.E.; Gould, O.N.; Lloyd, V. Knowledge and Knowledge Needs about Lyme Disease among Occupational and Recreational Users of the Outdoors. Int. J. Environ. Res. Public Health 2020, 17, 355. [Google Scholar] [CrossRef]
- Moore, A.; Nelson, C.; Molins, C.; Mead, P.; Schriefer, M. Current Guidelines, Common Clinical Pitfalls, and Future Directions for Laboratory Diagnosis of Lyme Disease, United States. Emerg. Infect. Dis. 2016, 22, 1167–1177. [Google Scholar] [CrossRef] [PubMed]
- Waddell, L.A.; Greig, J.; Mascarenhas, M.; Harding, S.; Lindsay, R.; Ogden, N. The Accuracy of Diagnostic Tests for Lyme Disease in Humans, A Systematic Review and Meta-Analysis of North American Research. PLoS ONE 2016, 11, e0168613. [Google Scholar] [CrossRef] [PubMed]
- Radolf, J.D.; Scott Samuels, D. Borrelia: Molecular Biology, Host Interaction and Pathogenesis; Horizon Scientific Press: Norwich, UK, 2010; ISBN 9781904455585. [Google Scholar]
- Marques, A.R. Laboratory Diagnosis of Lyme Disease: Advances and Challenges. Infect. Dis. Clin. North Am. 2015, 29, 295–307. [Google Scholar] [CrossRef] [PubMed]
- Zagorac, G.B.; Kezele, T.G. Ceftriaxone and Doxycycline Induced Seroconversion in Previously Seronegative Patient with Clinically Suspected Disseminated Lyme Disease: Case Report. Infect Chemother. 2021, 53, 582–588. [Google Scholar] [CrossRef]
- Coyle, P.K. Advances and Pitfalls in the Diagnosis of Lyme Disease. FEMS Immunol. Med. Microbiol. 1997, 19, 103–109. [Google Scholar] [CrossRef]
- Anderson, C.; Brissette, C.A. The Brilliance of Borrelia: Mechanisms of Host Immune Evasion by Lyme Disease-Causing Spirochetes. Pathogens 2021, 10, 281. [Google Scholar] [CrossRef]
- Wang, G.; Aguero-Rosenfeld, M.E.; Wormser, G.P. Detection of Borrelia Burgdorferi; Caister Academic Press: Norfolk, UK, 2010. [Google Scholar]
- Dumler, J. Molecular Diagnosis of Lyme Disease: Review and Meta-Analysis. Mol. Diagn. 2001, 6, 1–11. [Google Scholar] [CrossRef]
- Singh, S.K.; Girschick, H.J. Lyme Borreliosis: From Infection to Autoimmunity. Clin. Microbiol. Infect. 2004, 10, 598–614. [Google Scholar] [CrossRef]
- Norris, S.J.; Coburn, J.; Leong, J.M.; Hu, L.T.; Hook, M. Pathobiology of Lyme Disease Borrelia. In Borrelia: Molecular Biology, Host Interaction And Pathogenesis; Horizon Scientific Press: Norwich, UK, 2010; pp. 293–325. [Google Scholar]
- Barbour, A.G.; Guo, B.P. Pathogenesis of Relapsing Fever. Borrelia Mol. Biol. Host Interact. Pathogenesis 2010, 333–357. [Google Scholar]
- Dattwyler, R.J.; Wormser, G.P.; Rush, T.J.; Finkel, M.F.; Schoen, R.T.; Grunwaldt, E.; Franklin, M.; Hilton, E.; Bryant, G.L.; Agger, W.A.; et al. A Comparison of Two Treatment Regimens of Ceftriaxone in Late Lyme Disease. Wien. Klin. Wochenschr. 2005, 117, 393–397. [Google Scholar] [CrossRef]
- Donta, S.T. What We Know and Don’t Know About Lyme Disease. Front Public Health 2021, 9, 819541. [Google Scholar] [CrossRef] [PubMed]
- Tokarz, R.; Mishra, N.; Tagliafierro, T.; Sameroff, S.; Caciula, A.; Chauhan, L.; Patel, J.; Sullivan, E.; Gucwa, A.; Fallon, B.; et al. A Multiplex Serologic Platform for Diagnosis of Tick-Borne Diseases. Sci. Rep. 2018, 8, 3158. [Google Scholar] [CrossRef]
- Perronne, C. Lyme and Associated Tick-Borne Diseases: Global Challenges in the Context of a Public Health Threat. Front. Cell. Infect. Microbiol. 2014, 4, 74. [Google Scholar] [CrossRef]
- Maraspin, V.; Ogrinc, K.; Ružić-Sabljić, E.; Lotrič-Furlan, S.; Strle, F. Isolation of Borrelia Burgdorferi Sensu Lato from Blood of Adult Patients with Borrelial Lymphocytoma, Lyme Neuroborreliosis, Lyme Arthritis and Acrodermatitis Chronica Atrophicans. Infection 2011, 39, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Schmidli, J.; Hunziker, T.; Moesli, P.; Schaad, U.B. Cultivation of Borrelia Burgdorferi from Joint Fluid Three Months after Treatment of Facial Palsy due to Lyme Borreliosis. J. Infect. Dis. 1988, 158, 905–906. [Google Scholar] [CrossRef] [PubMed]
- Stanek, G.; Klein, J.; Bittner, R.; Glogar, D. Isolation of Borrelia Burgdorferi from the Myocardium of a Patient with Long-Standing Cardiomyopathy. N. Engl. J. Med. 1990, 322, 249–252. [Google Scholar] [CrossRef] [PubMed]
- Preac-Mursic, V.; Pfister, H.W.; Spiegel, H.; Burk, R.; Wilske, B.; Reinhardt, S.; Böhmer, R. First Isolation of Borrelia Burgdorferi from an Iris Biopsy. J. Clin. Neuroophthalmol. 1993, 13, 155–161, discussion 162. [Google Scholar] [PubMed]
- Chao, L.-L.; Chen, Y.-J.; Shih, C.-M. First Isolation and Molecular Identification of Borrelia Burgdorferi Sensu Stricto and Borrelia Afzelii from Skin Biopsies of Patients in Taiwan. Int. J. Infect. Dis. 2011, 15, e182–e187. [Google Scholar] [CrossRef][Green Version]
- Liveris, D.; Schwartz, I.; McKenna, D.; Nowakowski, J.; Nadelman, R.; DeMarco, J.; Iyer, R.; Bittker, S.; Cooper, D.; Holmgren, D.; et al. Comparison of Five Diagnostic Modalities for Direct Detection of Borrelia Burgdorferi in Patients with Early Lyme Disease. Diagn. Microbiol. Infect. Dis. 2012, 73, 243–245. [Google Scholar] [CrossRef]
- Liveris, D.; Wang, G.; Girao, G.; Byrne, D.W.; Nowakowski, J.; McKenna, D.; Nadelman, R.; Wormser, G.P.; Schwartz, I. Quantitative Detection of Borrelia Burgdorferi in 2-Millimeter Skin Samples of Erythema Migrans Lesions: Correlation of Results with Clinical and Laboratory Findings. J. Clin. Microbiol. 2002, 40, 1249–1253. [Google Scholar] [CrossRef]
- Eshoo, M.W.; Crowder, C.C.; Rebman, A.W.; Rounds, M.A.; Matthews, H.E.; Picuri, J.M.; Soloski, M.J.; Ecker, D.J.; Schutzer, S.E.; Aucott, J.N. Direct Molecular Detection and Genotyping of Borrelia Burgdorferi from Whole Blood of Patients with Early Lyme Disease. PLoS ONE 2012, 7, e36825. [Google Scholar] [CrossRef] [PubMed]
- Branda, J.A.; Steere, A.C. Laboratory Diagnosis of Lyme Borreliosis. Clin. Microbiol. Rev. 2021, 34, e00018-19. [Google Scholar] [CrossRef] [PubMed]
- Lambert, J.S.; Cook, M.J.; Healy, J.E.; Murtagh, R.; Avramovic, G.; Lee, S.H. Metagenomic 16S rRNA Gene Sequencing Survey of Borrelia Species in Irish Samples of Ixodes Ricinus Ticks. PLoS ONE 2019, 14, e0209881. [Google Scholar] [CrossRef] [PubMed]
- Schettig, R.; Tan-Lim, R.; Warren, D.; Poteet, Z.; Sears, R.; Hummel, M.; Coffin, C.; Quart, K.; Aussems, C.; Matthias, R., Jr.; et al. PathoDNA, an Advanced Diagnostic for Lyme Disease & Co-Infections Utilizing next Generation DNA Sequencing with Greater Sensitivity and Selectivity than ELISA/western Blot. Adv. Infect. Dis. 2021, 11, 405–429. [Google Scholar]
- Rauter, C.; Mueller, M.; Diterich, I.; Zeller, S.; Hassler, D.; Meergans, T.; Hartung, T. Critical Evaluation of Urine-Based PCR Assay for Diagnosis of Lyme Borreliosis. Clin. Diagn. Lab. Immunol. 2005, 12, 910–917. [Google Scholar] [CrossRef]
- Mosel, M.R.; Carolan, H.E.; Rebman, A.W.; Castro, S.; Massire, C.; Ecker, D.J.; Soloski, M.J.; Aucott, J.N.; Eshoo, M.W. Molecular Testing of Serial Blood Specimens from Patients with Early Lyme Disease during Treatment Reveals Changing Coinfection with Mixtures of Borrelia Burgdorferi Genotypes. Antimicrob. Agents Chemother. 2019, 63, e00237-19. [Google Scholar] [CrossRef]
- Eshoo, M.W.; Schutzer, S.E.; Crowder, C.D.; Carolan, H.E.; Ecker, D.J. Achieving Molecular Diagnostics for Lyme Disease. Expert Rev. Mol. Diagn. 2013, 13, 875–883. [Google Scholar] [CrossRef]
- Branda, J.A.; Lemieux, J.E.; Blair, L.; Ahmed, A.A.; Hong, D.K.; Bercovici, S.; Blauwkamp, T.A.; Hollemon, D.; Ho, C.; Strle, K.; et al. Detection of Borrelia Burgdorferi Cell-Free DNA in Human Plasma Samples for Improved Diagnosis of Early Lyme Borreliosis. Clin. Infect. Dis. 2021, 73, e2355–e2361. [Google Scholar] [CrossRef]
- Lee, S.H.; Healy, J.E.; Lambert, J.S. Single Core Genome Sequencing for Detection of Both Borrelia Burgdorferi Sensu Lato and Relapsing Fever Borrelia Species. Int. J. Environ. Res. Public Health 2019, 16, 1779. [Google Scholar] [CrossRef]
- Stefas, I.; Dubois, G.; Tigrett, S.; Lucarz, E.; Veas, F. Apolipoprotein H, an acute phase protein, a performing tool for ultra-sensitive detection and isolation of microorganisms from different origins. In Acute Phase Proteins as Early Non-Specific Biomarkers of Human and Veterinary Diseases; Veas, F., Ed.; InTechOpen: London, UK, 2011; pp. 21–42. [Google Scholar]
- Lacout, A.; Mone, Y.; Franck, M.; Marcy, P.-Y.; Mas, M.; Veas, F.; Perronne, C. Blood Cell Disruption to Significantly Improve the Borrelia PCR Detection Sensitivity in Borreliosis in Humans. Med. Hypotheses 2018, 116, 1–3. [Google Scholar] [CrossRef]
- Adlhoch, C.; Kaiser, M.; Hoehne, M.; Mas Marques, A.; Stefas, I.; Veas, F.; Ellerbrok, H. Highly Sensitive Detection of the Group A Rotavirus Using Apolipoprotein H-Coated ELISA Plates Compared to Quantitative Real-Time PCR. Virol. J. 2011, 8, 63. [Google Scholar] [CrossRef] [PubMed]
- Stefas, I.; Rucheton, M.; D’Angeac, A.D.; Morel-Baccard, C.; Seigneurin, J.M.; Zarski, J.P.; Martin, M.; Cerutti, M.; Bossy, J.P.; Missé, D.; et al. Hepatitis B Virus Dane Particles Bind to Human Plasma Apolipoprotein H. Hepatology 2001, 33, 207–217. [Google Scholar] [CrossRef]
- Stefas, E.; Rucheton, M.; Graafland, H.; Moynier, M.; Sompeyrac, C.; Bahraoui, E.M.; Veas, F. Human Plasmatic Apolipoprotein H Binds Human Immunodeficiency Virus Type 1 and Type 2 Proteins. AIDS Res. Hum. Retroviruses 1997, 13, 97–104. [Google Scholar] [CrossRef]
- Greissl, J.; Pesesky, M.; Dalai, S.C.; Rebman, A.W.; Soloski, M.J.; Horn, E.J.; Dines, J.N.; Gittelman, R.M.; Snyder, T.M.; Emerson, R.O.; et al. Immunosequencing of the T-Cell Receptor Repertoire Reveals Signatures Specific for Diagnosis and Characterization of Early Lyme Disease. bioRxiv 2021. [Google Scholar] [CrossRef]
- Hefter, Y.; D’Arco, C.; Shute, T.; Dattwyler, R.; Arnaboldi, P.; Nolan, S. 1760. Interferon Gamma Release Assay for Diagnosis of Lyme Disease. Open Forum Infect Dis 2018, 5, S61. [Google Scholar] [CrossRef]
- Shan, J.; Jia, Y.; Teulières, L.; Patel, F.; Clokie, M.R.J. Targeting Multicopy Prophage Genes for the Increased Detection of Borrelia Burgdorferi Sensu Lato (s.l.), the Causative Agents of Lyme Disease, in Blood. Front. Microbiol. 2021, 12, 651217. [Google Scholar] [CrossRef]
- Zhou, Y.; Qin, S.; Sun, M.; Tang, L.; Yan, X.; Kim, T.-K.; Caballero, J.; Glusman, G.; Brunkow, M.E.; Soloski, M.J.; et al. Measurement of Organ-Specific and Acute-Phase Blood Protein Levels in Early Lyme Disease. J. Proteome Res. 2020, 19, 346–359. [Google Scholar] [CrossRef]
- Marques, A.; Telford, S.R., 3rd; Turk, S.-P.; Chung, E.; Williams, C.; Dardick, K.; Krause, P.J.; Brandeburg, C.; Crowder, C.D.; Carolan, H.E.; et al. Xenodiagnosis to Detect Borrelia Burgdorferi Infection: A First-in-Human Study. Clin. Infect. Dis. 2014, 58, 937–945. [Google Scholar] [CrossRef]
- Murfin, K.E.; Kleinbard, R.; Aydin, M.; Salazar, S.A.; Fikrig, E. Borrelia Burgdorferi Chemotaxis toward Tick Protein Salp12 Contributes to Acquisition. Ticks Tick-Borne Dis. 2019, 10, 1124–1134. [Google Scholar] [CrossRef]
- Shih, C.-M.; Chao, L.-L.; Yu, C.-P. Chemotactic Migration of the Lyme Disease Spirochete (Borrelia Burgdorferi) to Salivary Gland Extracts of Vector Ticks. Am. J. Trop. Med. Hyg. 2002, 66, 616–621. [Google Scholar] [CrossRef]
- Embers, M.E.; Barthold, S.W.; Borda, J.T.; Bowers, L.; Doyle, L.; Hodzic, E.; Jacobs, M.B.; Hasenkampf, N.R.; Martin, D.S.; Narasimhan, S.; et al. Persistence of Borrelia Burgdorferi in Rhesus Macaques Following Antibiotic Treatment of Disseminated Infection. PLoS ONE 2012, 7, e29914. [Google Scholar] [CrossRef]
- Embers, M.E.; Hasenkampf, N.R.; Jacobs, M.B.; Tardo, A.C.; Doyle-Meyers, L.A.; Philipp, M.T.; Hodzic, E. Variable Manifestations, Diverse Seroreactivity and Post-Treatment Persistence in Non-Human Primates Exposed to Borrelia Burgdorferi by Tick Feeding. PLoS ONE 2017, 12, e0189071. [Google Scholar] [CrossRef] [PubMed]
- Bockenstedt, L.K.; Mao, J.; Hodzic, E.; Barthold, S.W.; Fish, D. Detection of Attenuated, Noninfectious Spirochetes inBorrelia burgdorferi–Infected Mice after Antibiotic Treatment. J. Infect. Dis. 2002, 186, 1430–1437. [Google Scholar] [CrossRef] [PubMed]
- Molins, C.R.; Ashton, L.V.; Wormser, G.P.; Hess, A.M.; Delorey, M.J.; Mahapatra, S.; Schriefer, M.E.; Belisle, J.T. Development of a Metabolic Biosignature for Detection of Early Lyme Disease. Clin. Infect. Dis. 2015, 60, 1767–1775. [Google Scholar] [CrossRef]
- Fitzgerald, B.L.; Molins, C.R.; Islam, M.N.; Graham, B.; Hove, P.R.; Wormser, G.P.; Hu, L.; Ashton, L.V.; Belisle, J.T. Host Metabolic Response in Early Lyme Disease. J. Proteome Res. 2020, 19, 610–623. [Google Scholar] [CrossRef]
- Haslund-Gourley, B.S.; Grauzam, S.; Mehta, A.S.; Wigdahl, B.; Comunale, M.A. Acute lyme disease IgG N-linked glycans contrast the canonical inflammatory signature. Front. Immunol. 2022, 13, 949118. [Google Scholar] [CrossRef]
- Fraser, C.M.; Casjens, S.; Huang, W.M.; Sutton, G.G.; Clayton, R.; Lathigra, R.; White, O.; Ketchum, K.A.; Dodson, R.; Hickey, E.K.; et al. Genomic Sequence of a Lyme Disease Spirochaete, Borrelia Burgdorferi. Nature 1997, 390, 580–586. [Google Scholar] [CrossRef]
- Picardeau, M.; Lobry, J.R.; Hinnebusch, B.J. Physical Mapping of an Origin of Bidirectional Replication at the Centre of the Borrelia Burgdorferi Linear Chromosome. Mol. Microbiol. 1999, 32, 437–445. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, K.T.; Wu, J.-C.; Boylan, J.A.; Gherardini, F.C.; Pei, D. Zinc Is the Metal Cofactor of Borrelia Burgdorferi Peptide Deformylase. Arch. Biochem. Biophys. 2007, 468, 217–225. [Google Scholar] [CrossRef]
- Ouyang, Z.; He, M.; Oman, T.; Yang, X.F.; Norgard, M.V. A Manganese Transporter, BB0219 (BmtA), Is Required for Virulence by the Lyme Disease Spirochete, Borrelia Burgdorferi. Proc. Natl. Acad. Sci. USA 2009, 106, 3449–3454. [Google Scholar] [CrossRef]
- Groshong, A.M.; Blevins, J.S. Insights into the Biology of Borrelia Burgdorferi Gained through the Application of Molecular Genetics. Adv. Appl. Microbiol. 2014, 86, 141–143. [Google Scholar]
- Posey, J.E.; Gherardini, F.C. Lack of a Role for Iron in the Lyme Disease Pathogen. Science 2000, 288, 1651–1653. [Google Scholar] [CrossRef]
- Troxell, B.; Xu, H.; Frank Yang, X. Borrelia Burgdorferi, a Pathogen That Lacks Iron, Encodes Manganese-Dependent Superoxide Dismutase Essential for Resistance to Streptonigrin. J. Biol. Chem. 2012, 287, 19284–19293. [Google Scholar] [CrossRef] [PubMed]
- Aguirre, J.D.; Clark, H.M.; McIlvin, M.; Vazquez, C.; Palmere, S.L.; Grab, D.J.; Seshu, J.; Hart, P.J.; Saito, M.; Culotta, V.C. A Manganese-Rich Environment Supports Superoxide Dismutase Activity in a Lyme Disease Pathogen, Borrelia Burgdorferi. J. Biol. Chem. 2013, 288, 8468–8478. [Google Scholar] [CrossRef] [PubMed]
- Troxell, B.; Yang, X.F. Metal-Dependent Gene Regulation in the Causative Agent of Lyme Disease. Front. Cell. Infect. Microbiol. 2013, 3, 79. [Google Scholar] [CrossRef] [PubMed]
- Casjens, S.; Eggers, C.H.; Schwartz, I. Borrelia Genomics: Chromosome, Plasmids, Bacteriophages and Genetic Variation. Borrelia: Molecular Biology, Host Interaction and Pathogenesis; Caister Academic Press: Hethersett, UK, 2010; pp. 27–53. [Google Scholar]
- Margos, G.; Hojgaard, A.; Lane, R.S.; Cornet, M.; Fingerle, V.; Rudenko, N.; Ogden, N.; Aanensen, D.M.; Fish, D.; Piesman, J. Multilocus Sequence Analysis of Borrelia Bissettii Strains from North America Reveals a New Borrelia Species, Borrelia Kurtenbachii. Ticks Tick Borne Dis. 2010, 1, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Borrelia carolinensis Sp. Nov., a New (14th) Member of the Borrelia Burgdorferi Sensu Lato Complex from the Southeastern Region of the United States. J. Clin. Microbiol. 2009, 47, 134–141. [Google Scholar] [CrossRef]
- Rudenko, N.; Golovchenko, M.; Lin, T.; Gao, L.; Grubhoffer, L.; Oliver, J.H., Jr. Delineation of a New Species of the Borrelia Burgdorferi Sensu Lato Complex, Borrelia Americana Sp. Nov. J. Clin. Microbiol. 2009, 47, 3875–3880. [Google Scholar] [CrossRef]
- Stanek, G.; Reiter, M. The Expanding Lyme Borrelia Complex—Clinical Significance of Genomic Species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef]
- Lescot, M.; Audic, S.; Robert, C.; Nguyen, T.T.; Blanc, G.; Cutler, S.J.; Wincker, P.; Couloux, A.; Claverie, J.-M.; Raoult, D.; et al. The Genome of Borrelia Recurrentis, the Agent of Deadly Louse-Borne Relapsing Fever, Is a Degraded Subset of Tick-Borne Borrelia Duttonii. PLoS Genet. 2008, 4, e1000185. [Google Scholar] [CrossRef]
- Kuleshov, K.V.; Margos, G.; Fingerle, V.; Koetsveld, J.; Goptar, I.A.; Markelov, M.L.; Kolyasnikova, N.M.; Sarksyan, D.S.; Kirdyashkina, N.P.; Shipulin, G.A.; et al. Whole Genome Sequencing of Borrelia Miyamotoi Isolate Izh-4: Reference for a Complex Bacterial Genome. BMC Genom. 2020, 21, 16. [Google Scholar] [CrossRef] [PubMed]
- Carlyon, J.A.; Marconi, R.T. Cloning and Molecular Characterization of a Multicopy, Linear Plasmid-Carried, Repeat Motif-Containing Gene from Borrelia Turicatae, a Causative Agent of Relapsing Fever. J. Bacteriol. 1998, 180, 4974–4981. [Google Scholar] [CrossRef] [PubMed]
- Genomics of Relapsing Fever Spirochetes. In Lyme Disease and Relapsing Fever Spirochetes: Genomics, Molecular Biology, Host Interactions and Disease Pathogenesis; Caister Academic Press: Hethersett, UK, 2021; ISBN 9781913652616.
- Barbour, A.G. Phylogeny of a Relapsing Fever Borrelia Species Transmitted by the Hard Tick Ixodes Scapularis. Infect. Genet. Evol. 2014, 27, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Purser, J.E.; Norris, S.J. Correlation between Plasmid Content and Infectivity in Borrelia Burgdorferi. Proc. Natl. Acad. Sci. USA 2000, 97, 13865–13870. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, I.; Margos, G.; Casjens, S.R.; Qiu, W.-G.; Eggers, C.H. Multipartite Genome of Lyme Disease Borrelia: Structure, Variation and Prophages. Curr. Issues Mol. Biol. 2021, 42, 409–454. [Google Scholar]
- Casjens, S.R. The Unusual Linear Plasmids of the Lyme Disease Spirochete Borrelia Burgdorferi. Biochem. Soc. Transact. 2000, 28, A102. [Google Scholar] [CrossRef]
- Casjens, S.R.; Gilcrease, E.B.; Vujadinovic, M.; Mongodin, E.F.; Luft, B.J.; Schutzer, S.E.; Fraser, C.M.; Qiu, W.-G. Plasmid Diversity and Phylogenetic Consistency in the Lyme Disease Agent Borrelia Burgdorferi. BMC Genom. 2017, 18, 165. [Google Scholar] [CrossRef]
- Balakirev, E.S.; Ayala, F.J. Pseudogenes: Are They “Junk” or Functional DNA? Annu. Rev. Genet. 2003, 37, 123–151. [Google Scholar] [CrossRef]
- Casjens, S.R.; Di, L.; Akther, S.; Mongodin, E.F.; Luft, B.J.; Schutzer, S.E.; Fraser, C.M.; Qiu, W.-G. Primordial Origin and Diversification of Plasmids in Lyme Disease Agent Bacteria. BMC Genom. 2018, 19, 218. [Google Scholar] [CrossRef]
- Dulebohn, D.P.; Bestor, A.; Rosa, P.A. Borrelia Burgdorferi Linear Plasmid 28-3 Confers a Selective Advantage in an Experimental Mouse-Tick Infection Model. Infect. Immun. 2013, 81, 2986–2996. [Google Scholar] [CrossRef]
- Kobryn, K. Replication of the Borrelia Burgdorferi Genome. In Lyme Disease and Relapsing Fever Spirochetes: Genomics, Molecular Biology, Host Interactions and Disease Pathogenesis 2021; Caister Academic Press: Wymondham, UK, 2021. [Google Scholar]
- Kobryn, K.; Chaconas, G. ResT, a Telomere Resolvase Encoded by the Lyme Disease Spirochete. Mol. Cell 2002, 9, 195–201. [Google Scholar] [CrossRef]
- Iyer, R.; Kalu, O.; Purser, J.; Norris, S.; Stevenson, B.; Schwartz, I. Linear and Circular Plasmid Content in Borrelia Burgdorferi Clinical Isolates. Infect. Immun. 2003, 71, 3699–3706. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, B.; Porcella, S.F.; Oie, K.L.; Fitzpatrick, C.A.; Raffel, S.J.; Lubke, L.; Schrumpf, M.E.; Schwan, T.G. The Relapsing Fever Spirochete Borrelia Hermsii Contains Multiple, Antigen-Encoding Circular Plasmids That Are Homologous to the cp32 Plasmids of Lyme Disease Spirochetes. Infect. Immun. 2000, 68, 3900–3908. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Innamorati, K.A.; Earl, J.P.; Aggarwal, S.D.; Ehrlich, G.D.; Hiller, N.L. The Bacterial Guide to Designing a Diversified Gene Portfolio. In The Pangenome: Diversity, Dynamics and Evolution of Genomes; Tettelin, H., Medini, D., Eds.; Springer International Publishing: Cham, The Netherlands, 2020; pp. 51–87. ISBN 9783030382810. [Google Scholar]
- van Vliet, A.H.M. Use of Pan-Genome Analysis for the Identification of Lineage-Specific Genes of Helicobacter Pylori. FEMS Microbiol. Lett. 2017, 364, fnw296. [Google Scholar] [CrossRef]
- Hogg, J.S.; Hu, F.Z.; Janto, B.; Boissy, R.; Hayes, J.; Keefe, R.; Post, J.C.; Ehrlich, G.D. Characterization and Modeling of the Haemophilus Influenzae Core and Supragenomes Based on the Complete Genomic Sequences of Rd and 12 Clinical Nontypeable Strains. Genome Biol. 2007, 8, R103. [Google Scholar] [CrossRef]
- Forgetta, V.; Oughton, M.T.; Marquis, P.; Brukner, I.; Blanchette, R.; Haub, K.; Magrini, V.; Mardis, E.R.; Gerding, D.N.; Loo, V.G.; et al. Fourteen-Genome Comparison Identifies DNA Markers for Severe-Disease-Associated Strains of Clostridium Difficile. J. Clin. Microbiol. 2011, 49, 2230–2238. [Google Scholar] [CrossRef]
- Udaondo, Z.; Duque, E.; Ramos, J.-L. The Pangenome of the Genus Clostridium. Environ. Microbiol. 2017, 19, 2588–2603. [Google Scholar] [CrossRef]
- Marshall, D.J.; Heisler, L.M.; Lyamichev, V.; Murvine, C.; Olive, D.M.; Ehrlich, G.D.; Neri, B.P.; de Arruda, M. Determination of Hepatitis C Virus Genotypes in the United States by Cleavase Fragment Length Polymorphism Analysis. J. Clin. Microbiol. 1997, 35, 3156–3162. [Google Scholar] [CrossRef]
- Shen, K.; Wang, X.; Post, J.C. Molecular and Translational Research Approaches for the Study of Bacterial Pathogenesis in Otitis Media. In Evidence-Based Otitis Media, 2nd ed.; Rosenfeld, R., Bluestone, C.D., Eds.; B.C. Decker Inc.: Hamilton, ON, Canada, 2003; pp. 91–119. [Google Scholar]
- Ehrlich, G.D.; Hu, F.Z.; Post, J.C. Role for Biofilms in Infectious Disease. In Microbial Biofilms; American Society of Microbiology: Washington, DC, USA, 2004; pp. 332–358. ISBN 9781555818944. [Google Scholar]
- Gladitz, J.; Shen, K.; Antalis, P.; Hu, F.Z.; Post, J.C.; Ehrlich, G.D. Codon Usage Comparison of Novel Genes in Clinical Isolates of Haemophilus Influenzae. Nucleic Acids Res. 2005, 33, 3644–3658. [Google Scholar] [CrossRef][Green Version]
- Shen, K.; Antalis, P.; Gladitz, J.; Sayeed, S.; Ahmed, A.; Yu, S.; Hayes, J.; Johnson, S.; Dice, B.; Dopico, R.; et al. Identification, Distribution, and Expression of Novel Genes in 10 Clinical Isolates of Nontypeable Haemophilus Influenzae. Infect. Immun. 2005, 73, 3479–3491. [Google Scholar] [CrossRef]
- Shen, K.; Gladitz, J.; Antalis, P.; Dice, B.; Janto, B.; Keefe, R.; Hayes, J.; Ahmed, A.; Dopico, R.; Ehrlich, N.; et al. Characterization, Distribution, and Expression of Novel Genes among Eight Clinical Isolates of Streptococcus Pneumoniae. Infect. Immun. 2006, 74, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, P.G.; Morphew, R.M.; Whitworth, D.E. Genome Sequencing and Pan-Genome Analysis of 23 Corallococcus spp. Strains Reveal Unexpected Diversity, With Particular Plasticity of Predatory Gene Sets. Front. Microbiol. 2018, 9, 3187. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Zaiden, N.; Cao, B. The Core- and Pan-Genomic Analyses of the Genus Comamonas: From Environmental Adaptation to Potential Virulence. Front. Microbiol. 2018, 9, 3096. [Google Scholar] [CrossRef] [PubMed]
- Earl, J.P.; de Vries, S.P.W.; Ahmed, A.; Powell, E.; Schultz, M.P.; Hermans, P.W.M.; Hill, D.J.; Zhou, Z.; Constantinidou, C.I.; Hu, F.Z.; et al. Comparative Genomic Analyses of the Moraxella Catarrhalis Serosensitive and Seroresistant Lineages Demonstrate Their Independent Evolution. Genome Biol. Evol. 2016, 8, 955–974. [Google Scholar] [CrossRef]
- Nistico, L.; Earl, J.; Hiller, L. Using the Core and Supra Genomes to Determine Diversity and Natural Proclivities among Bacterial Strains. In Application of Molecular Microbiological Methods; Skovhus, T.L., Caffrey, S., Hubert, C., Eds.; Caister Academic Press: Norfolk, UK, 2014. [Google Scholar]
- Kress-Bennett, J.M.; Hiller, N.L.; Eutsey, R.A.; Powell, E.; Longwell, M.J.; Hillman, T.; Blackwell, T.; Byers, B.; Mell, J.C.; Post, J.C.; et al. Identification and Characterization of Msf, a Novel Virulence Factor in Haemophilus Influenzae. PLoS ONE 2016, 11, e0149891. [Google Scholar]
- Margos, G.; Hepner, S.; Mang, C.; Marosevic, D.; Reynolds, S.E.; Krebs, S.; Sing, A.; Derdakova, M.; Reiter, M.A.; Fingerle, V. Lost in Plasmids: Next Generation Sequencing and the Complex Genome of the Tick-Borne Pathogen Borrelia Burgdorferi. BMC Genom. 2017, 18, 422. [Google Scholar] [CrossRef]
- Mongodin, E.F.; Casjens, S.R.; Bruno, J.F.; Xu, Y.; Drabek, E.F.; Riley, D.R.; Cantarel, B.L.; Pagan, P.E.; Hernandez, Y.A.; Vargas, L.C.; et al. Inter- and Intra-Specific Pan-Genomes of Borrelia Burgdorferi Sensu Lato: Genome Stability and Adaptive Radiation. BMC Genom. 2013, 14, 693. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Amblyomma americanum | Amblyomma maculatum | Dermacentor andersonii | Dermacentor variabilis | Ixodes cookei | Ixodes pacificus | Ixodes scapularis | Rhicephalus sanguineus | Ornithodoros hermsii | |
|---|---|---|---|---|---|---|---|---|---|
| Common name 1 | Lone Star Tick | Gulf Coast Tick | Rocky Mountain Wood Tick | American Dog Tick | Groundhog Tick | Western Blacklegged tick | Blacklegged Tick/Deer Tick | Brown Dog Tick | |
| Type 3 | Hard | Hard | Hard | Hard | Hard | Hard | Hard | Hard | Soft |
| Prey 1,3 | rodents, domestic animals | rodents, domestic animals | rodents, domestic animals | domestic animals | groundhogs, skunks, rodents, racoons, foxes, weasels, domestic animals | mice, voles, weasels, deer, birds, lizards | mice, deer, birds | domestic animals | rodents |
| Feeding style 3 | 3-host tick | 3-host tick | 3-host tick | 3-host tick | 3-host tick | 3-host | 3-host | 3-host tick | |
| Dimorphic 3 | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes | Yes |
| Zone 1,2 | East and Southern US | South-mid US, Southern Arizona | East of Rocky Mountains, US, pacific coast | Eastern U.S | Western US | Eastern, Upper Midwest, and Southern US | World-wide, * Southern border of U.S | Northwest coast, Texas, Florida | |
| Peak activity 1,2 | Early Spring- Late fall | Early Spring- Late fall | Spring and Summer | Early spring-Mid Fall | Early spring-Mid Fall | Early spring-Mid Fall | Year-round | ||
| States 1,2 | AL, AR, CT, DE, FL, GA, IL, IN, IA, KS, KY, LA, ME, MD, MA, MS, MO, NE, NH, NJ, NY, NC, OH, OK, PA, RI, SC, TN, TX, VT, VA, WV, DC | AL, AR, FL, GA, KS, LA, MS, MO, NC, OK, SC, TN, TX, VA | AZ, CA, CO, ID, KS, MT NE, NV, NM, ND, OK, OR, SD, UT, WA, WY | AL, AR, CA, CT, DE, FL, GA, IL, IN, IA, KS, KY, LA, ME, MD, MA, MI, MN, MS, MO, MT, NE NH, NJ, NY, NC, ND, OH, OK, PA, RI, SC, SD, TN, TX, VT, VA, WV, WI, WY, DC | ME | AZ, CA, NV, OR, UT, WA | AL, AR, CT, DE, FL, GA, IL, IN, IA, KS, KY, LA, ME, MD, MA, MI, MN, MS, MO, NH, NJ, NY, NC, ND, OH, OK, PA, RI, SC, SD, TN, TX, VT, VA, WV, WI, DC | AL, AK, AZ, AR, CA, CO, CT, DE, FL, GA, HI, ID, IL, IN, IA, KS, KY, LA, ME, MD MA, MI, MN, MS, MO, MT, NE, NV, NH, NJ, NM, NY, NC, ND, OH, OK, OR, PA. RI, SC, SD, TN, TX, UT, VT, VA, WA, WV, WI, WY, DC | |
| Habitat 1,2 | Wooded areas | Coastal areas | scrublands, lightly wooded areas, and open grasslands | Human settlements | Human settlements |
| Babesia microti | Anaplasma phagocytophilum | Borrelia burgdorferi | Borrelia miyamotoi | Borrelia mayonii | Ehrlichia chafeensis | Ehrlichia muris | Ehrlichia ewingii | Francisella tularensis | Rickettsia parkeri | Rickettsia rickettsii | Bourbon virus | Coltivirus | Heartland virus | Powassan virus | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Type | Parasite | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Bacteria | Virus | Virus | Virus | Virus |
| Disease Name 1 | Babesiosis | Anaplasmosis | Lyme Borreliosis | Relapsing Fever | Lyme borreliosis | Ehrlichiosis | Ehrlichiosis | Ehrlichiosis | Tularemia | R. parkeri spotted fever | Rocky Mountain Spotted Fever | Bourbon virus disease | Colorado Tick Fever | Heartland virus disease | Powassan virus disease |
| Discovery 1 | 1990 [63] | 1932 [64] | 1982 | 1995 [65] | 2016 [66] | 1986 [67] | 2009 [68] | 1996 | 1912 [69] | 1937 [70] | 1906 | 2014 [71] | 1950 [72] | 2012 [73] | 1990 [74] |
| Reservoir 1 | Small mammals: [55] Peromyscus leucopus, Procyon lotor, Blarina brevicauda, Tamias striatus | Peromyscus leucopus, Odocoileus virginianus | Peromyscus leucopus, Odocoileus virginianus, Tamias striatus, Blarina brevicauda, Sorex cinereus, Sciurus carolinensis, etc. [53] | Peromyscus leucopus, Apodemus spp., Microtus spp., Tamias spp., Sciuridae spp., etc. [75] | Peromyscus leucopus, Tamiasciurus hudsonicus [76] | Odocoileus virginianus | Peromyscus leucopus | Odocoileus virginianus | Rodents | Unknown | Deer | ||||
| TickVector 1 | I. scapularis | I. scapularis | I. scapularis | I. scapularis | I. pacificus | A. americanum | I. scapularis | A. americanum | A.Americanum, D. variabilis, D. andersonii | D. andersonii | D. variabilis, R. sanguineus, D. andersonii | A. americanum | D. andersonii | A. americanum | I. scapularis, I. cookei |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Socarras, K.M.; Haslund-Gourley, B.S.; Cramer, N.A.; Comunale, M.A.; Marconi, R.T.; Ehrlich, G.D. Large-Scale Sequencing of Borreliaceae for the Construction of Pan-Genomic-Based Diagnostics. Genes 2022, 13, 1604. https://doi.org/10.3390/genes13091604
Socarras KM, Haslund-Gourley BS, Cramer NA, Comunale MA, Marconi RT, Ehrlich GD. Large-Scale Sequencing of Borreliaceae for the Construction of Pan-Genomic-Based Diagnostics. Genes. 2022; 13(9):1604. https://doi.org/10.3390/genes13091604
Chicago/Turabian StyleSocarras, Kayla M., Benjamin S. Haslund-Gourley, Nicholas A. Cramer, Mary Ann Comunale, Richard T. Marconi, and Garth D. Ehrlich. 2022. "Large-Scale Sequencing of Borreliaceae for the Construction of Pan-Genomic-Based Diagnostics" Genes 13, no. 9: 1604. https://doi.org/10.3390/genes13091604
APA StyleSocarras, K. M., Haslund-Gourley, B. S., Cramer, N. A., Comunale, M. A., Marconi, R. T., & Ehrlich, G. D. (2022). Large-Scale Sequencing of Borreliaceae for the Construction of Pan-Genomic-Based Diagnostics. Genes, 13(9), 1604. https://doi.org/10.3390/genes13091604