
In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients


Abstract
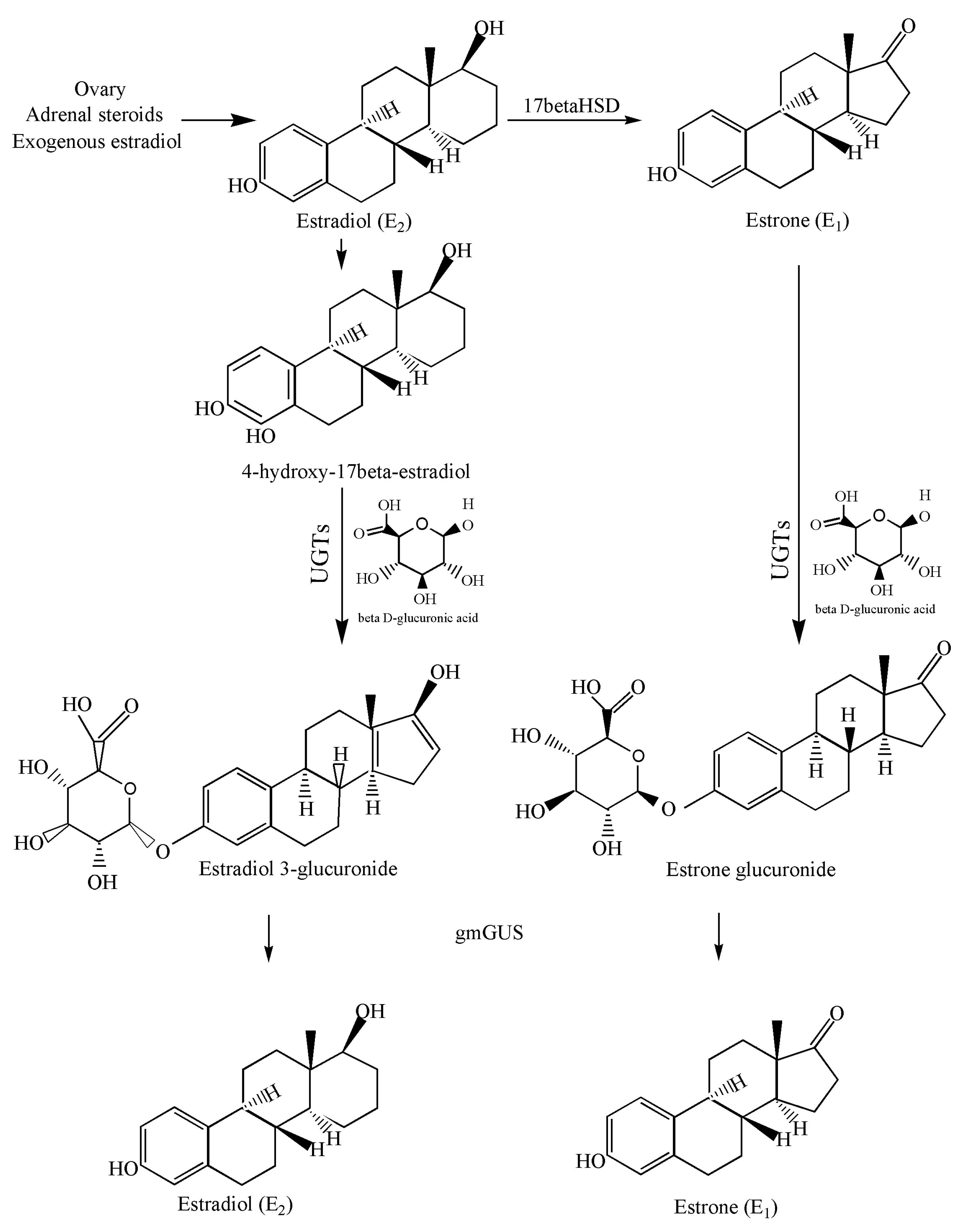
1. Introduction
2. Methodology
2.1. Protein Sequences
2.2. Phylogeny Analysis
2.3. Prediction of Physicochemical Properties
2.4. Sub-Cellular Localization and Ontology Analysis
2.5. 2D Structure
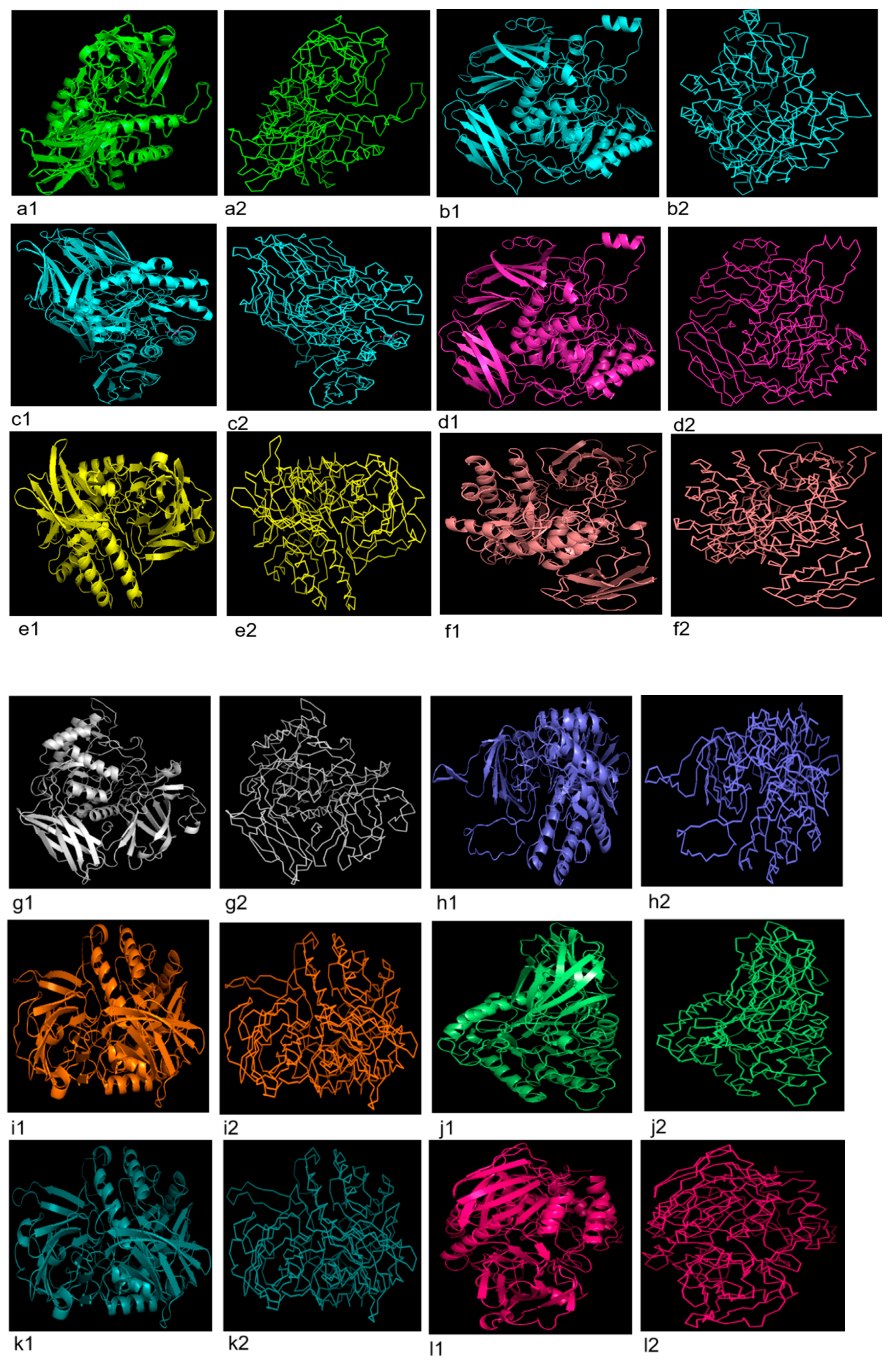
2.6. 3D Structure
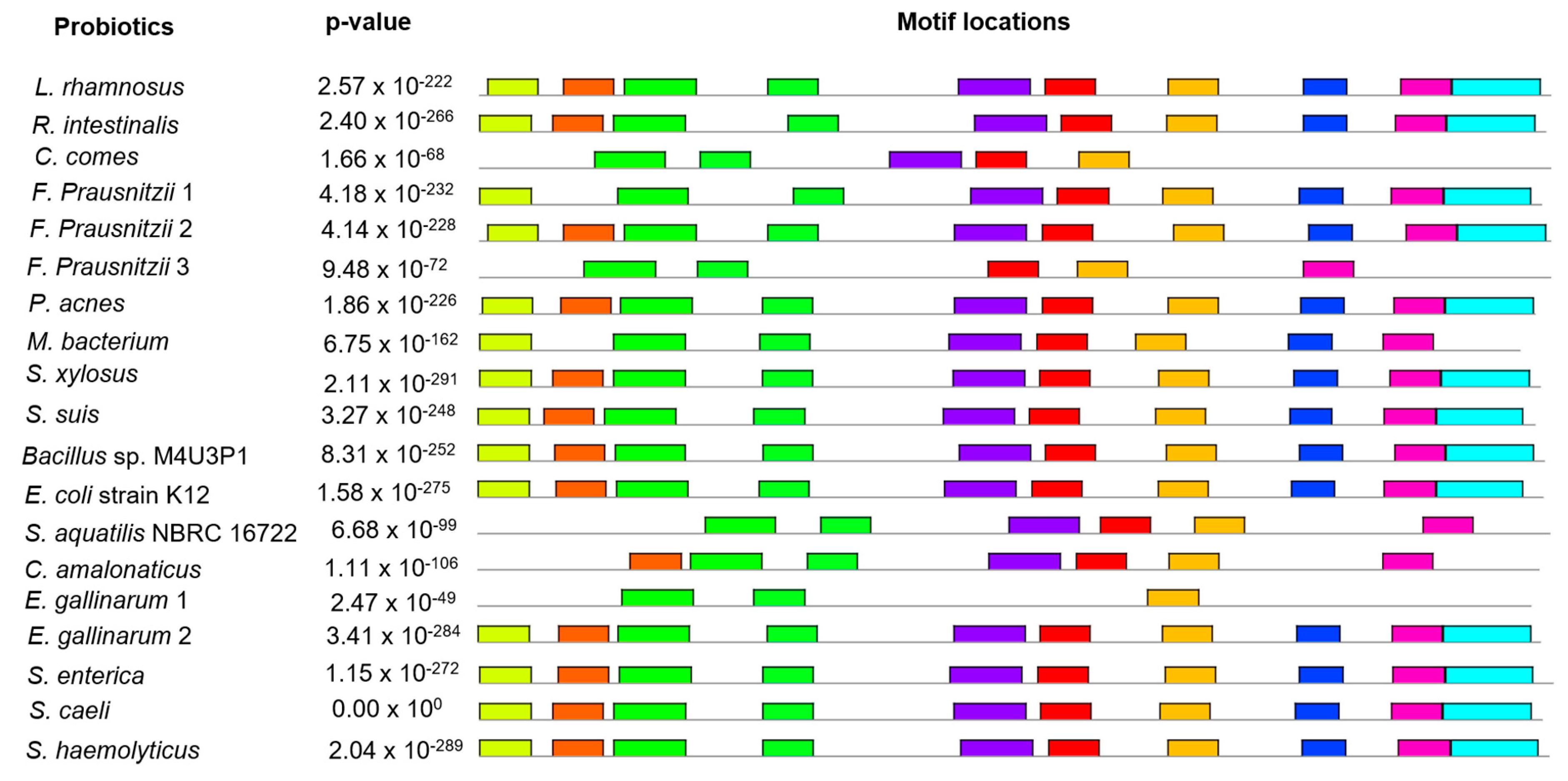
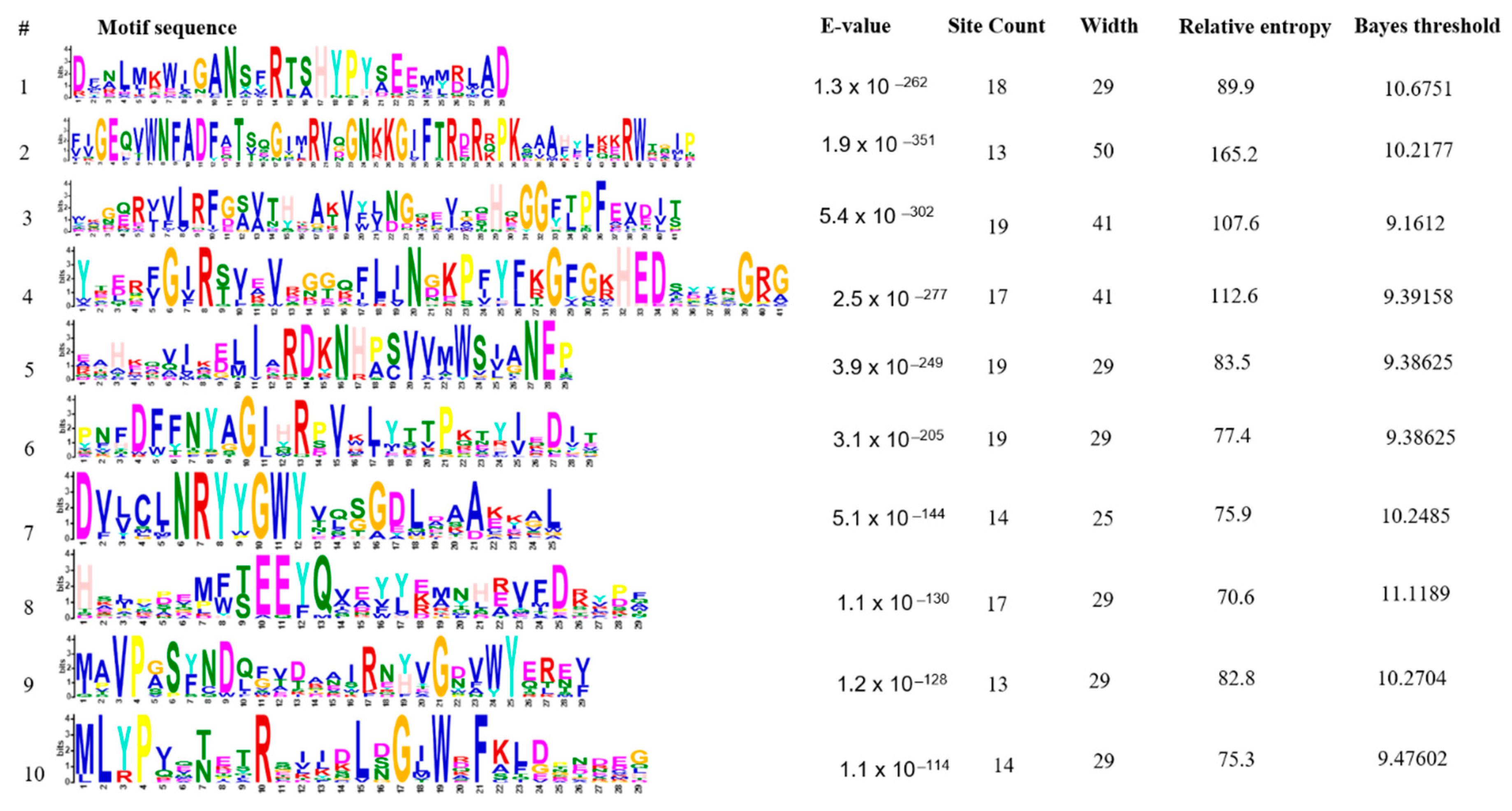
2.7. Conserved Protein Motifs Analysis
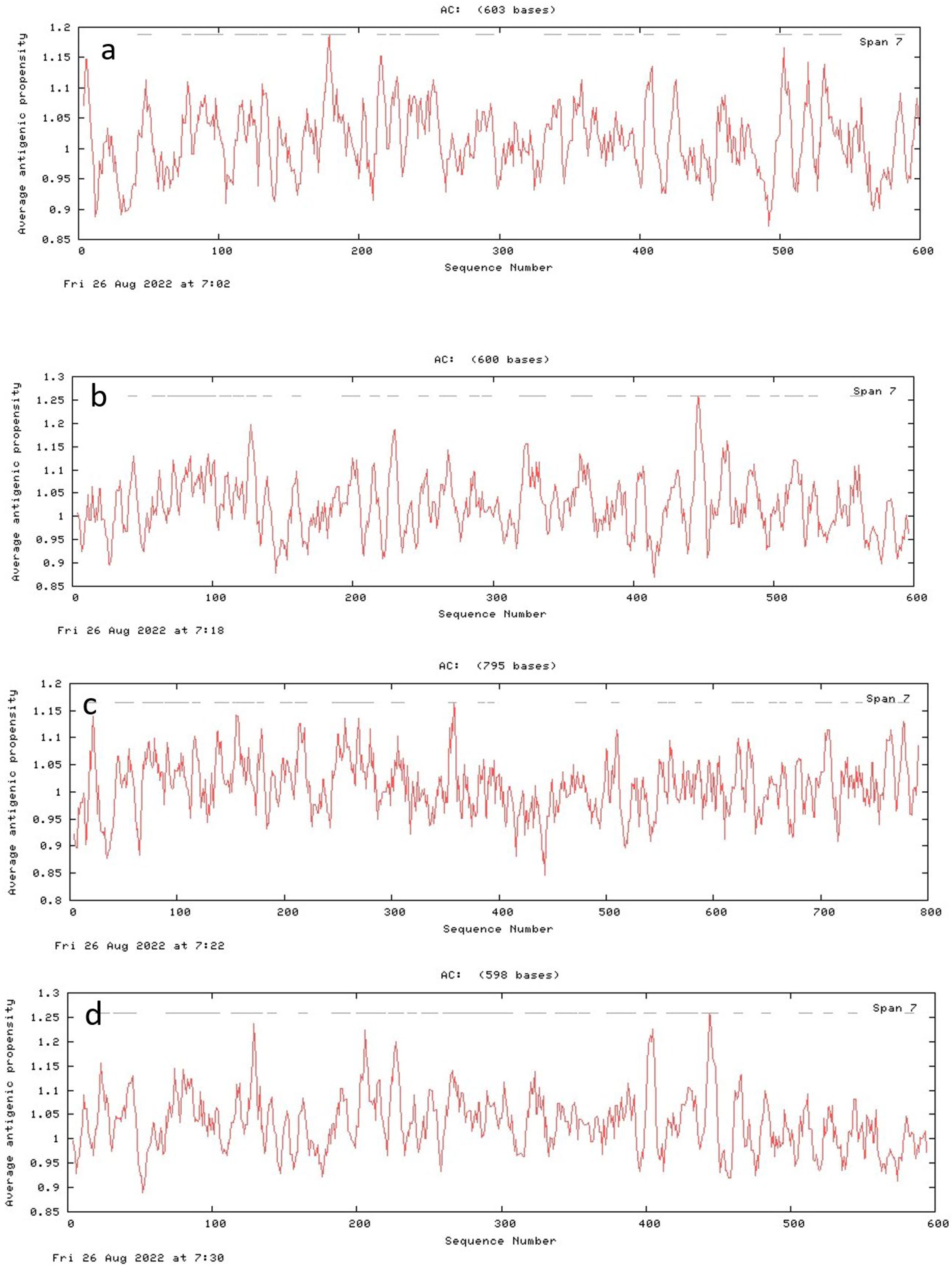
2.8. Predicted Antigenic Peptides Tool
3. Results
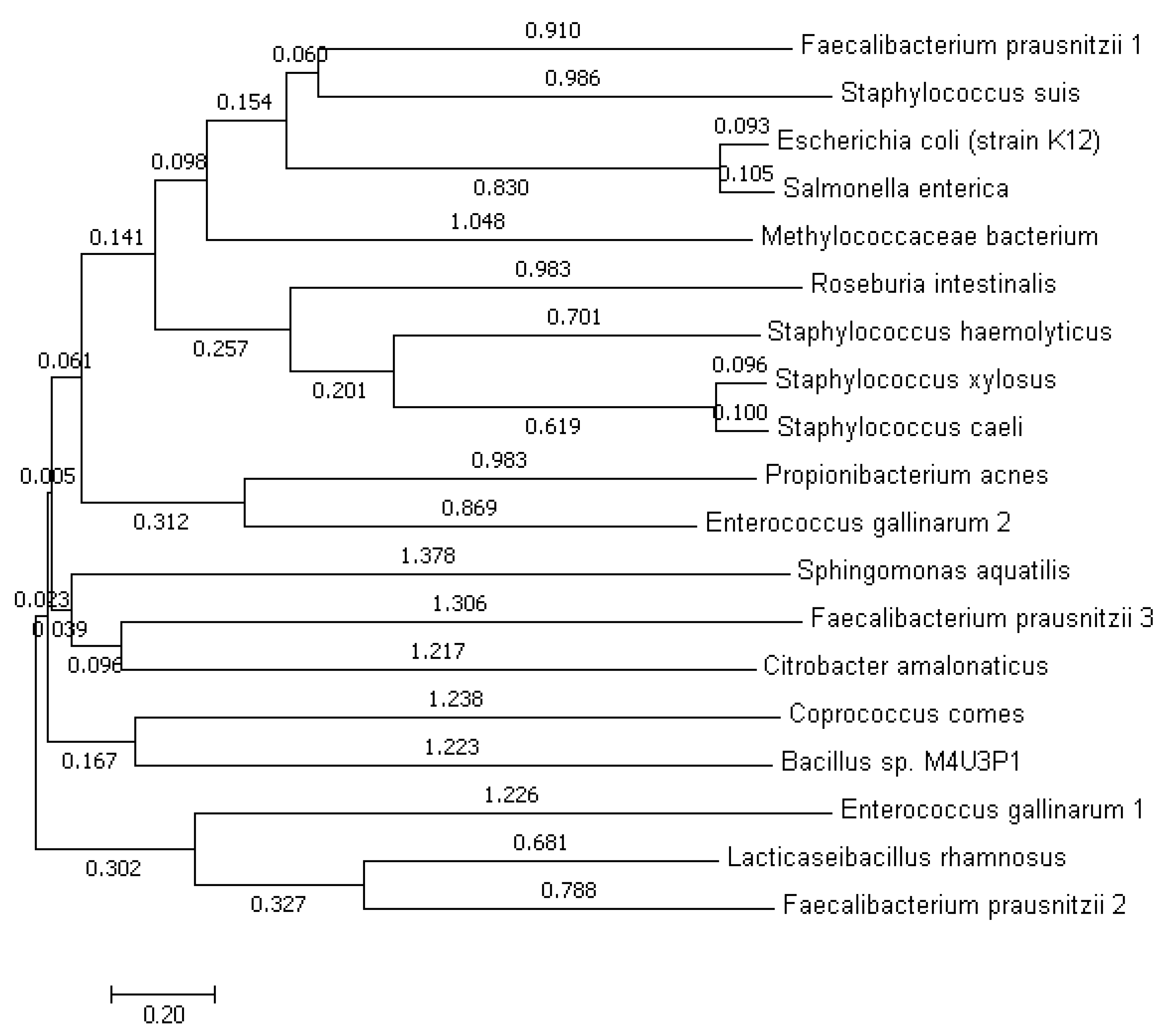
3.1. Phylogeny Prediction Based on uidA Gene Sequence
3.2. Physicochemical Attributes
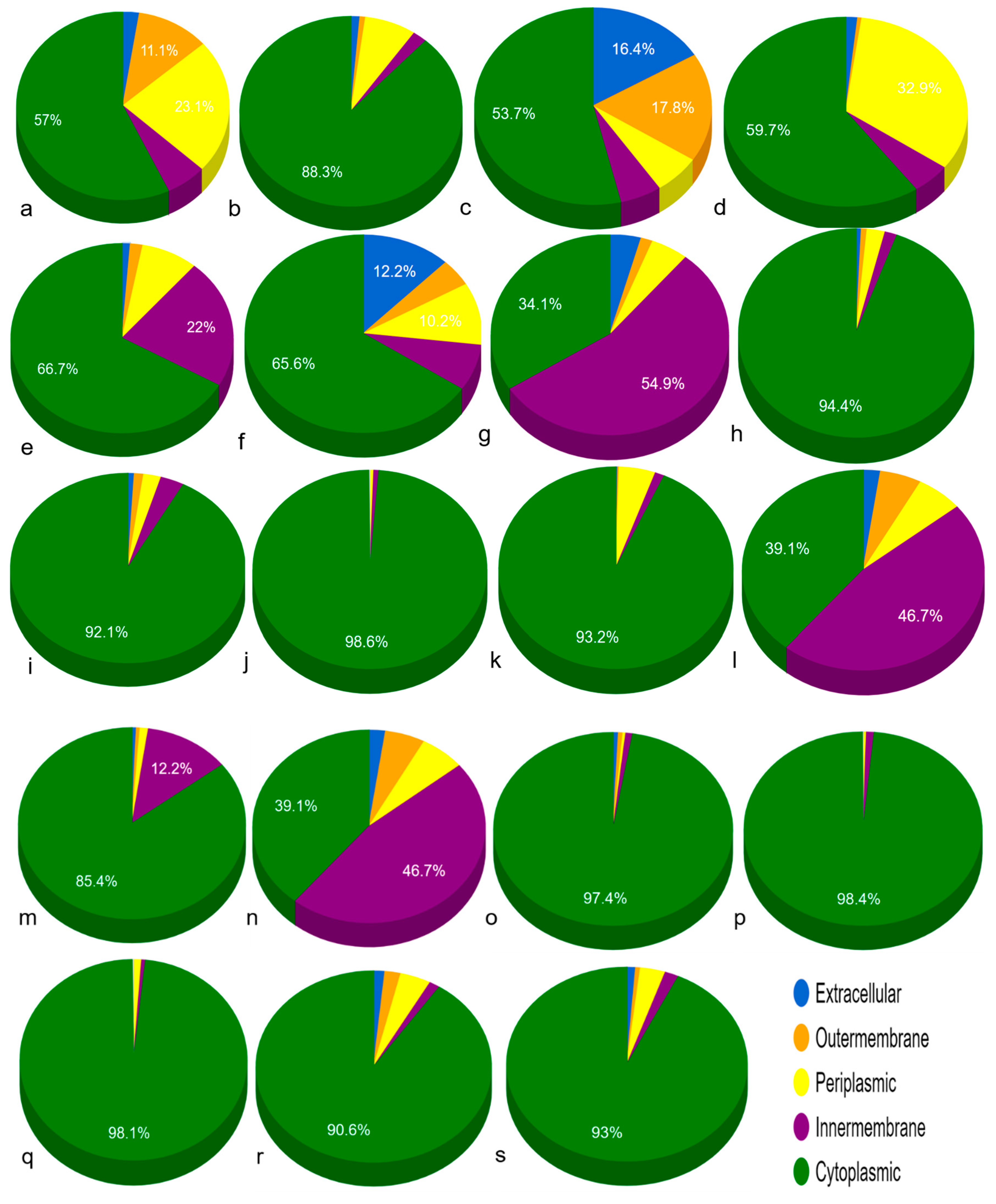
3.3. Sub-Cellular Localization
3.4. 2D Structure Prediction
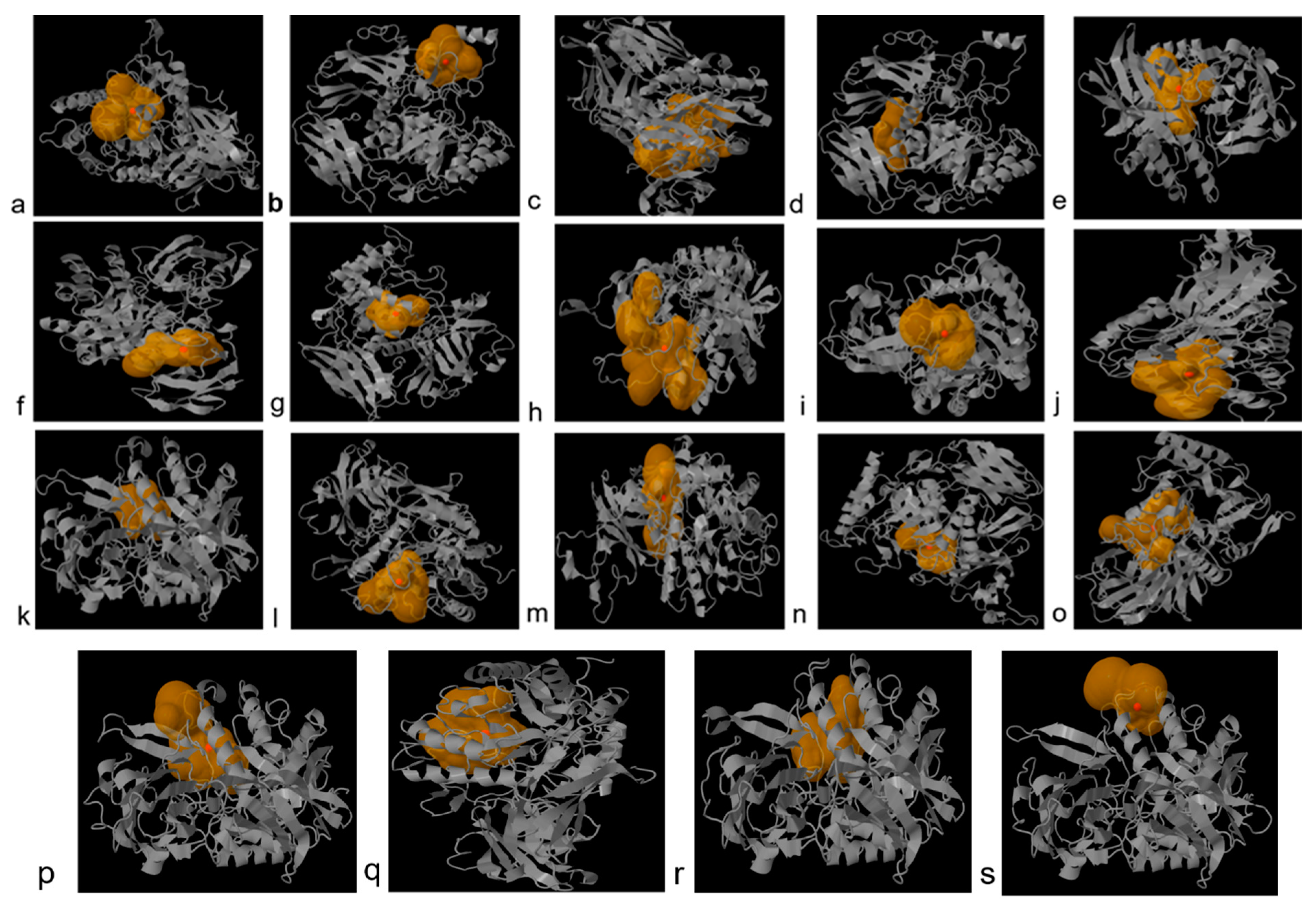
3.5. 3D Structure Prediction
3.6. Conserved Protein Motifs Prediction
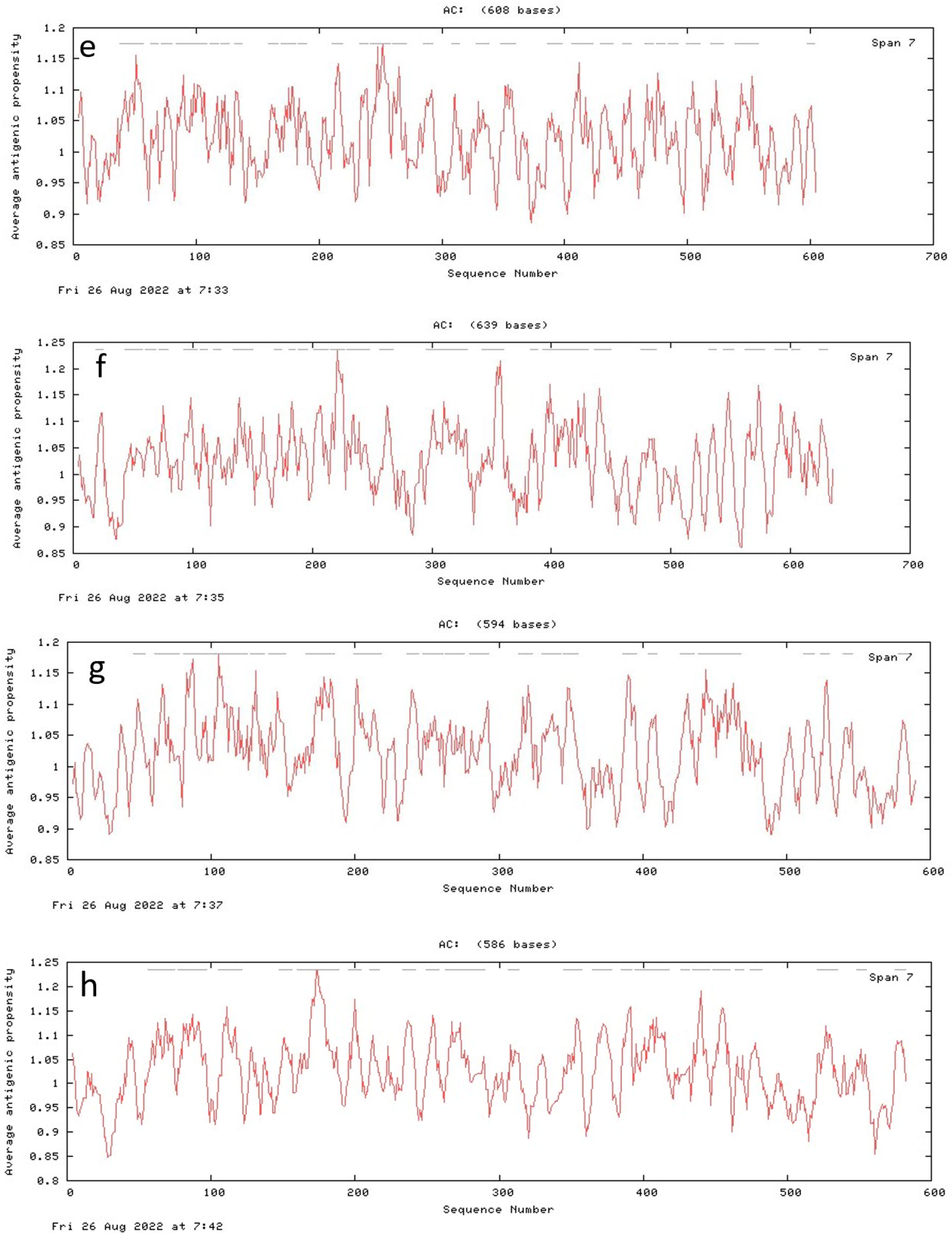
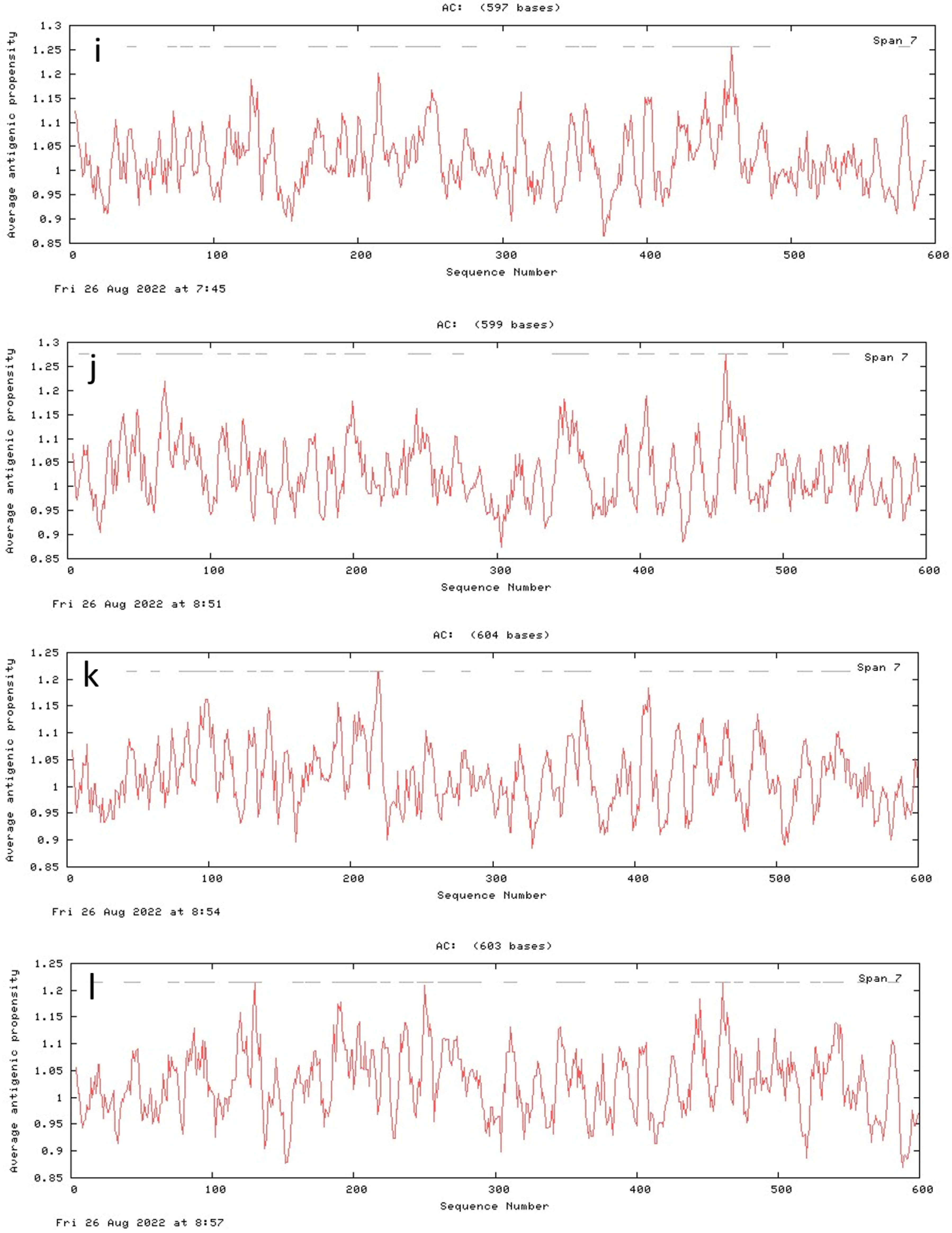
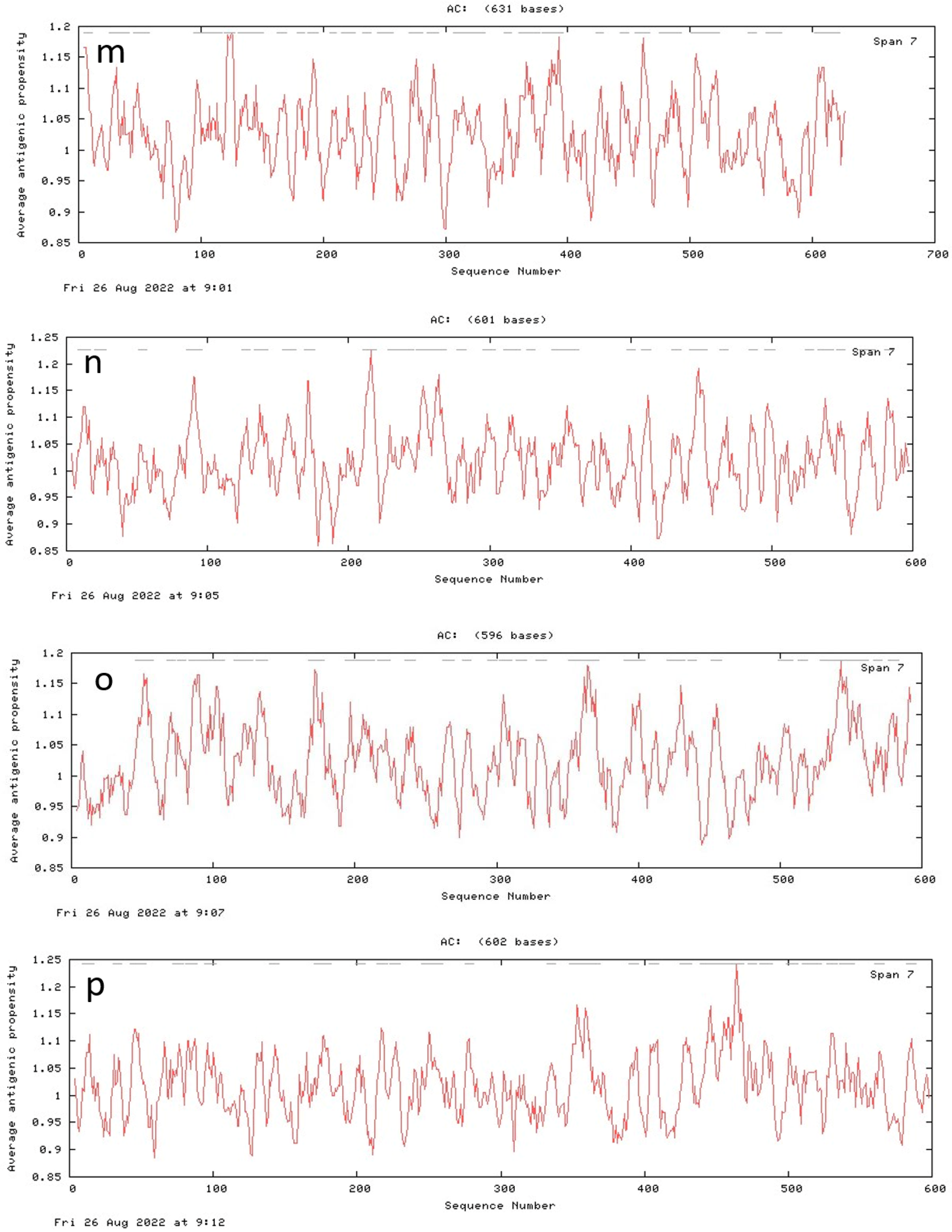
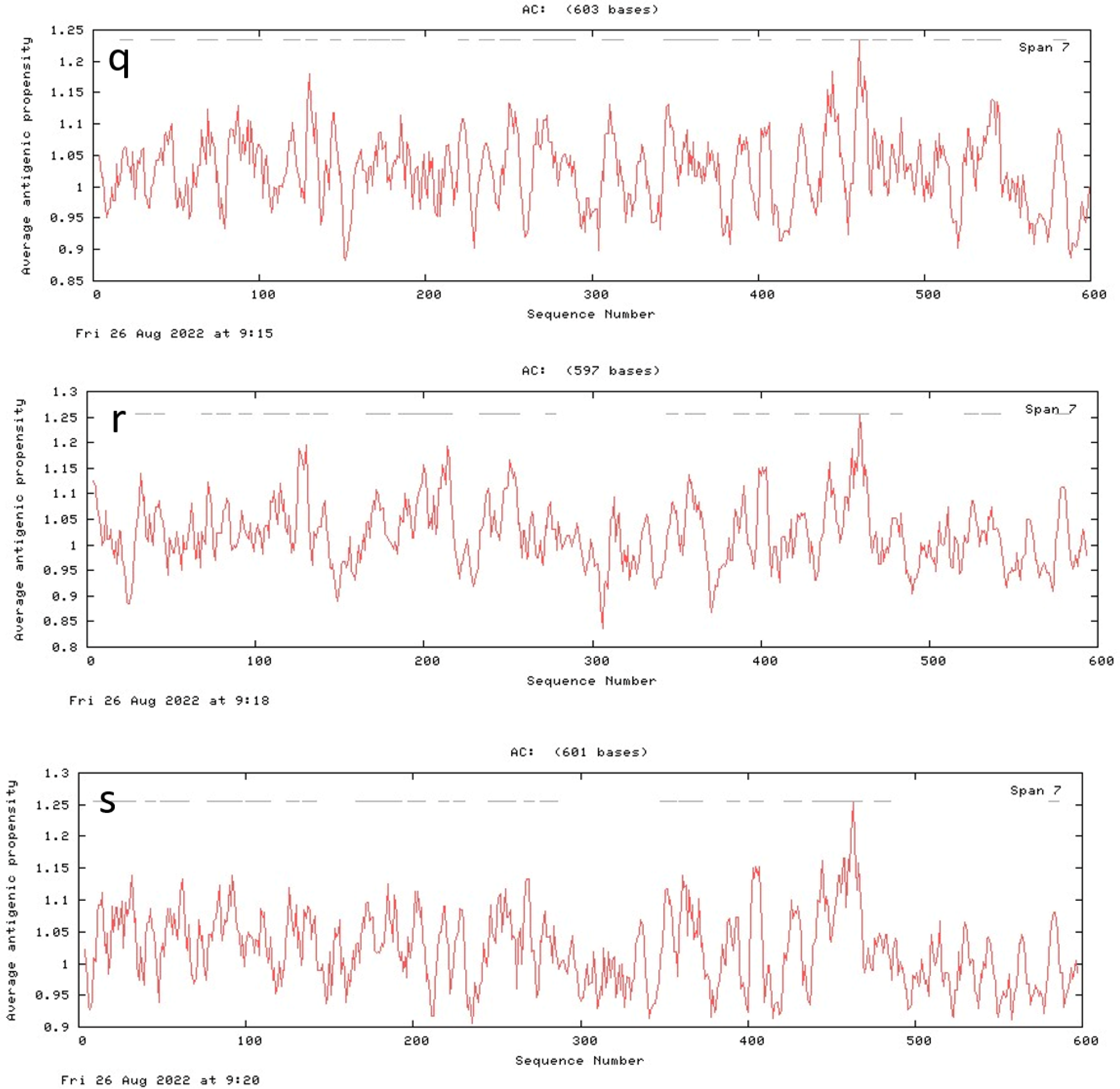
3.7. Antigenic Peptide Prediction
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ganguly, N.K.; Bhattacharya, S.K.; Sesikeran, B.; Nair, G.B.; Ramakrishna, B.S.; Sachdev, H.P.S.; Batish, V.K.; Kanagasabapathy, A.S.; Muthuswamy, V.; Kathuria, S.C. ICMR-DBT guidelines for evaluation of probiotics in food. Indian J. Med. Res. 2011, 134, 22. [Google Scholar]
- Fuller, R. Probiotics in man and animals. J. Appl. Bacteriol. 1989, 66, 365–378. [Google Scholar]
- Nagpal, R.; Kumar, A.; Kumar, M.; Behare, P.V.; Jain, S.; Yadav, H. Probiotics, their health benefits and applications for developing healthier foods: A review. FEMS Microbiol. Lett. 2012, 334, 1–15. [Google Scholar]
- Şener, D.; Bulut, H.N.; Güneş Bayir, A. Probiotics and Relationship between Probiotics and Cancer Types. Bezmialem Sci. 2021, 9, 490–497. [Google Scholar] [CrossRef]
- Lee, J.H.; Nam, S.H.; Seo, W.T.; Yun, H.D.; Hong, S.Y.; Kim, M.K.; Cho, K.M. The production of surfactin during the fermentation of cheonggukjang by potential probiotic Bacillus subtilis CSY191 and the resultant growth suppression of MCF-7 human breast cancer cells. Food Chem. 2012, 131, 1347–1354. [Google Scholar] [CrossRef]
- Bland, K.I.; Copeland, E.M.; Klimberg, V.S.; Gradishar, W.J. The Breast E-Book: Comprehensive Management of Benign and Malignant Diseases; Elsevier Health Sciences: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2019. Cancer J. Clin. 2019, 69, 7–34. [Google Scholar] [CrossRef]
- Kumar, V.; Naik, B.; Kumar, A.; Khanduri, N.; Rustagi, S.; Kumar, S. Probiotics media: Significance, challenges, and future perspective-a mini review. Food Prod. Process Nutr. 2022, 4, 1–13. [Google Scholar]
- Doron, S.I.; Hibberd, P.L.; Gorbach, S.L. Probiotics for prevention of antibiotic-associated diarrhea. J. Clin. Gastroenterol. 2008, 42, S58–S63. [Google Scholar] [CrossRef]
- Choi, C.H.; Jo, S.Y.; Park, H.J.; Chang, S.K.; Byeon, J.S.; Myung, S.J. A randomized, double-blind, placebo-controlled multicenter trial of Saccharomyces boulardii in irritable bowel syndrome: Effect on quality of life. J. Clin. Gastroenterol. 2011, 45, 679–683. [Google Scholar] [CrossRef]
- Xuan, C.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PLoS ONE 2014, 9, e83744. [Google Scholar]
- Dieleman, S.; Aarnoutse, R.; Ziemons, J.; Kooreman, L.; Boleij, A.; Smidt, M. Exploring the potential of breast microbiota as biomarker for breast cancer and therapeutic response. AME J. Pathol. 2021, 191, 968–982. [Google Scholar] [CrossRef]
- Wang, H.; Altemus, J.; Niazi, F.; Green, H.; Calhoun, B.C.; Sturgis, C.; Grobmyer, S.R.; Eng, C. Breast tissue, oral and urinary microbiomes in breast cancer. Oncotarget 2017, 8, 88122. [Google Scholar]
- Hieken, T.J.; Chen, J.; Hoskin, T.L.; Walther-Antonio, M.; Johnson, S.; Ramaker, S.; Xiao, J.; Radisky, D.C.; Knutson, K.L.; Kalari, K.R. The microbiome of aseptically collected human breast tissue in benign and malignant disease. Sci. Rep. 2016, 6, 30751. [Google Scholar] [CrossRef]
- Komorowski, A.S.; Pezo, R.C. Untapped “-omics”: The microbial metagenome, estrobolome, and their influence on the development of breast cancer and response to treatment. Breast Cancer Res. Treat. 2020, 179, 287–300. [Google Scholar]
- Alizadehmohajer, N.; Shojaeifar, S.; Nedaeinia, R.; Esparvarinha, M.; Mohammadi, F.; Ferns, G.A.; Ghayour-Mobarhan, M.; Manian, M.; Balouchi, A. Association between the microbiota and women’s cancers–Cause or consequences? Biomed. Pharm. 2020, 127, 110203. [Google Scholar]
- Faith, J.J.; McNulty, N.P.; Rey, F.E.; Gordon, J.I. Predicting a human gut microbiota’s response to diet in gnotobiotic mice. Science 2011, 333, 101–104. [Google Scholar] [CrossRef]
- Aminov, R.I.; Walker, A.W.; Duncan, S.H.; Harmsen, H.J.M.; Welling, G.W.; Flint, H.J. Molecular diversity, cultivation, and improved detection by fluorescent in situ hybridization of a dominant group of human gut bacteria related to Roseburia spp. or Eubacterium rectale. Appl. Environ. Microbiol. 2006, 72, 6371–6376. [Google Scholar] [CrossRef]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The microbiota of breast tissue and its association with breast cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar]
- Esposito, M.V.; Fosso, B.; Nunziato, M.; Casaburi, G.; D’Argenio, V.; Calabrese, A.; D’Aiuto, M.; Botti, G.; Pesole, G.; Salvatore, F. Microbiome composition indicate dysbiosis and lower richness in tumor breast tissues compared to healthy adjacent paired tissue, within the same women. BMC Cancer 2022, 22, 30. [Google Scholar] [CrossRef]
- Ma, J.; Sun, L.; Liu, Y.; Ren, H.; Shen, Y.; Bi, F.; Zhang, T.; Wang, X. Alter between gut bacteria and blood metabolites and the anti-tumor effects of Faecalibacterium prausnitzii in breast cancer. BMC Microbiol. 2020, 20, 1–19. [Google Scholar] [CrossRef]
- He, C.; Liu, Y.; Ye, S.; Yin, S.; Gu, J. Changes of intestinal microflora of breast cancer in premenopausal women. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 503–513. [Google Scholar] [CrossRef]
- Costantini, L.; Magno, S.; Albanese, D.; Donati, C.; Molinari, R.; Filippone, A.; Masetti, R.; Merendino, N. Characterization of human breast tissue microbiota from core needle biopsies through the analysis of multi hypervariable 16S-rRNA gene regions. Sci. Rep. 2018, 8, 16893. [Google Scholar] [CrossRef]
- Awolade, P.; Cele, N.; Kerru, N.; Gummidi, L.; Oluwakemi, E.; Singh, P. Therapeutic significance of β-glucuronidase activity and its inhibitors: A review. Eur. J. Med. Chem. 2020, 187, 111921. [Google Scholar] [CrossRef]
- Pollet, R.M.; D’Agostino, E.H.; Walton, W.G.; Xu, Y.; Little, M.S.; Biernat, K.A.; Pellock, S.J.; Patterson, L.M.; Creekmore, B.C.; Isenberg, H.N. An atlas of β-glucuronidases in the human intestinal microbiome. Structure 2017, 25, 967–977.e965. [Google Scholar] [CrossRef]
- Wallace, B.D.; Wang, H.; Lane, K.T.; Scott, J.E.; Orans, J.; Koo, J.S.; Venkatesh, M.; Jobin, C.; Yeh, L.A.; Mani, S. Alleviating cancer drug toxicity by inhibiting a bacterial enzyme. Science 2010, 330, 831–835. [Google Scholar] [CrossRef]
- Pellock, S.J.; Walton, W.G.; Biernat, K.A.; Torres-Rivera, D.; Creekmore, B.C.; Xu, Y.; Liu, J.; Tripathy, A.; Stewart, L.J.; Redinbo, M.R. Three structurally and functionally distinct β-glucuronidases from the human gut microbe Bacteroides uniformis. J. Biol. Chem. 2018, 293, 18559–18573. [Google Scholar] [CrossRef]
- Ervin, S.M.; Li, H.; Lim, L.; Roberts, L.R.; Liang, X.; Mani, S.; Redinbo, M.R. Gut microbial β-glucuronidases reactivate estrogens as components of the estrobolome that reactivate estrogens. J. Biol. Chem. 2019, 294, 18586–18599. [Google Scholar] [CrossRef]
- Travis, R.C.; Key, T.J. Oestrogen exposure and breast cancer risk. Breast Cancer Res. 2003, 5, 239. [Google Scholar] [CrossRef]
- Schiffer, L.; Barnard, L.; Baranowski, E.S.; Gilligan, L.C.; Taylor, A.E.; Arlt, W.; Shackleton, C.H.L.; Storbeck, K.H. Human steroid biosynthesis, metabolism and excretion are differentially reflected by serum and urine steroid metabolomes: A comprehensive review. J. Steroid Biochem. Mol. Biol. 2019, 194, 105439. [Google Scholar]
- Adlercreutz, H.; Martin, F. Biliary excretion and intestinal metabolism of progesterone and estrogens in man. J. Steroid Biochem. 1980, 13, 231–244. [Google Scholar] [CrossRef]
- Flores, R.; Shi, J.; Fuhrman, B.; Xu, X.; Veenstra, T.D.; Gail, M.H.; Gajer, P.; Ravel, J.; Goedert, J.J. Fecal microbial determinants of fecal and systemic estrogens and estrogen metabolites: A cross-sectional study. J. Transl. Med. 2012, 10, 1–11. [Google Scholar] [CrossRef]
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk factors and preventions of breast cancer. Int. J. Biol. Sci. 2017, 13, 1387. [Google Scholar] [CrossRef]
- Godone, R.L.N.; Leitão, G.M.; Araújo, N.B.; Castelletti, C.H.M.; Lima-Filho, J.L.; Martins, D.B.G. Clinical and molecular aspects of breast cancer: Targets and therapies. Biomed. Pharm. 2018, 106, 14–34. [Google Scholar] [CrossRef]
- Kumar, S.; Srivastav, R.K.; Wilkes, D.W.; Ross, T.; Kim, S.; Kowalski, J.; Chatla, S.; Zhang, Q.; Nayak, A.; Guha, M. Estrogen-dependent DLL1-mediated Notch signaling promotes luminal breast cancer. Oncogene 2019, 38, 2092–2107. [Google Scholar] [CrossRef]
- Gupta, P.B.; Proia, D.; Cingoz, O.; Weremowicz, J.; Naber, S.P.; Weinberg, R.A.; Kuperwasser, C. Systemic stromal effects of estrogen promote the growth of estrogen receptor–negative cancers. Cancer Res. 2007, 67, 2062–2071. [Google Scholar] [CrossRef]
- Jiang, C.F.; Shi, Z.M.; Li, D.M.; Qian, Y.C.; Ren, Y.; Bai, X.M.; Xie, Y.X.; Wang, L.; Ge, X.; Liu, W.T. Estrogen-induced miR-196a elevation promotes tumor growth and metastasis via targeting SPRED1 in breast cancer. Mol. Cancer 2018, 17, 1–18. [Google Scholar] [CrossRef]
- Zeng, L.; Zielinska, H.A.; Arshad, A.; Shield, J.P.; Bahl, A.; Holly, J.M.P.; Perks, C.M. Hyperglycaemia-induced chemoresistance in breast cancer cells: Role of the estrogen receptor. Endocr. Relat. Cancer 2016, 23, 125–134. [Google Scholar] [CrossRef]
- Consortium, U. UniProt: A hub for protein information. Nucleic Acids Res. 2015, 43, D204–D212. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. The clustal omega multiple alignment package. In Multiple Sequence Alignment; Springer: Berlin/Heidelberg, Germany, 2021; pp. 3–16. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Zuckerkandl, E.; Pauling, L. Evolutionary divergence and convergence in proteins. In Evolving Genes and Proteins; Elsevier: Amsterdam, The Netherlands, 1965; pp. 97–166. [Google Scholar]
- Geourjon, C.; Deleage, G. SOPMA: Significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Bioinformatics 1995, 11, 681–684. [Google Scholar] [CrossRef] [PubMed]
- Stourac, J.; Vavra, O.; Kokkonen, P.; Filipovic, J.; Pinto, G.; Brezovsky, J.; Damborsky, J.; Bednar, D. Caver Web 1.0: Identification of tunnels and channels in proteins and analysis of ligand transport. Nucleic Acids Res. 2019, 47, W414–W422. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Vihinen, M. Spectrum of disease-causing mutations in protein secondary structures. BMC Struct. Biol. 2007, 7, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Agnihotry, S.; Pathak, R.K.; Singh, D.B.; Tiwari, A.; Hussain, I. Protein structure prediction. In Bioinformatics; Elsevier: Amsterdam, The Netherlands, 2022; pp. 177–188. [Google Scholar]
- Kwa, M.; Plottel, C.S.; Blaser, M.J.; Adams, S. The intestinal microbiome and estrogen receptor–positive female breast cancer. J. Natl. Cancer Inst. 2016, 108. [Google Scholar] [CrossRef]
- Wang, P.; Jia, Y.; Wu, R.; Chen, Z.; Yan, R. Human gut bacterial β-glucuronidase inhibition: An emerging approach to manage medication therapy. Biochem. Pharm. 2021, 190, 114566. [Google Scholar] [CrossRef]
- Beaud, D.; Tailliez, P.; Anba-Mondoloni, J. Genetic characterization of the β-glucuronidase enzyme from a human intestinal bacterium, Ruminococcus gnavus. Microbiology 2005, 151, 2323–2330. [Google Scholar] [CrossRef]
- Kim, D.; Jin, Y.; Jung, E.; Han, M.; Kobashi, K. Purification and characterization of β-glucuronidase from Escherichia coli HGU-3, a human intestinal bacterium. Biol. Pharm. Bull. 1995, 18, 1184–1188. [Google Scholar] [CrossRef]
- Kim, D.; Kang, H.; Park, S.; Kobashi, K. Characterization of β-glucosidase and β-glucuronidase of alkalotolerant intestinal bacteria. Biol Pharm. Bull. 1994, 17, 423–426. [Google Scholar] [CrossRef]
- Morya, V.K.; Yadav, S.; Kim, E.K.; Yadav, D. In silico characterization of alkaline proteases from different species of Aspergillus. Appl. Biochem. Biotechnol. 2012, 166, 243–257. [Google Scholar] [CrossRef]
- Gamage, D.G.; Gunaratne, A.; Periyannan, G.R.; Russell, T.G. Applicability of instability index for in vitro protein stability prediction. Protein Pept. Lett. 2019, 26, 339–347. [Google Scholar] [CrossRef]
- Barril, X. Druggability predictions: Methods, limitations, and applications. Wiley Interdiscip. Rev. Comput. Mol. Sci 2013, 3, 327–338. [Google Scholar] [CrossRef]
- Michel, M.; Homan, E.J.; Wiita, E.; Pedersen, K.; Almlöf, I.; Gustavsson, A.L.; Lundbäck, T.; Helleday, T.; Warpman Berglund, U. In silico druggability assessment of the NUDIX hydrolase protein family as a workflow for target prioritization. Front. Chem. 2020, 8, 443. [Google Scholar] [CrossRef] [PubMed]
- Kingsley, L.J.; Lill, M.A. Substrate tunnels in enzymes: Structure–function relationships and computational methodology. Proteins Struct. Funct. Bioinform. 2015, 83, 599–611. [Google Scholar] [CrossRef]
- Wong, A.; Gehring, C.; Irving, H.R. Conserved functional motifs and homology modeling to predict hidden moonlighting functional sites. Front. Bioeng. Biotechnol. 2015, 3, 82. [Google Scholar] [CrossRef]
- Sun, Z.; Fu, P.; Wei, K.; Zhang, H.; Zhang, Y.; Xu, J.; Jiang, F.; Liu, X.; Xu, W.; Wu, W. Identification of novel immunogenic proteins from Mycoplasma bovis and establishment of an indirect ELISA based on recombinant E1 β subunit of the pyruvate dehydrogenase complex. PLoS ONE 2014, 9, e88328. [Google Scholar] [CrossRef]
- Sperker, B.; Backman, J.T.; Kroemer, H.K. The role of β-glucuronidase in drug disposition and drug targeting in humans. Clin. Pharm. 1997, 33, 18–31. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| # | Probiotics | No. of Amino Acids | Mol. Wt. | pI | Half-Life (Hours) | Extinction Coefficient | Instability Index | Aliphatic Index |
|---|---|---|---|---|---|---|---|---|
| 1 | L. rhamnosus | 603 | 68,570.92 | 5.35 | 30 | 101,760 | 30.86 | 71.01 |
| 2 | R. intestinalis | 600 | 68,337.66 | 5.06 | 30 | 109,140 | 33.60 | 73.75 |
| 3 | C. comes | 795 | 87,709.17 | 5.85 | 30 | 126,920 | 26.87 | 78.06 |
| 4 | F. prausnitzii 1 | 598 | 67,535.00 | 5.34 | 30 | 119,345 | 35.75 | 74.03 |
| 5 | F. prausnitzii 2 | 608 | 69,345.36 | 6.13 | 30 | 118,970 | 33.77 | 75.41 |
| 6 | F. prausnitzii 3 | 639 | 72,500.39 | 5.92 | 30 | 131,960 | 32.82 | 71.85 |
| 7 | P. acnes | 594 | 67,236.34 | 5.12 | 30 | 125,040 | 33.75 | 78.94 |
| 8 | M. bacterium | 586 | 66,604.06 | 5.91 | 30 | 131,125 | 36.17 | 81.98 |
| 9 | S. xylosus | 597 | 67,983.63 | 5.21 | 30 | 89,980 | 31.77 | 82.84 |
| 10 | S. suis | 599 | 68,912.07 | 5.11 | 30 | 95,355 | 29.20 | 88.10 |
| 11 | Bacillus sp. M4U3P1 | 604 | 68,912.10 | 5.17 | 30 | 100,855 | 37.19 | 80.98 |
| 12 | E. coli (strain K12) | 603 | 68,447.00 | 5.24 | 30 | 140,760 | 26.68 | 77.74 |
| 13 | S. aquatilis NBRC 16722 | 631 | 69,237.47 | 8.92 | 30 | 124,455 | 37.69 | 83.88 |
| 14 | C. amalonaticus | 601 | 69,674.51 | 5.33 | 30 | 119,805 | 40.93 | 80.80 |
| 15 | E. gallinarum 1 | 596 | 69,632.94 | 5.01 | 30 | 155,075 | 25.88 | 75.87 |
| 16 | E. gallinarum 2 | 602 | 69,494.74 | 5.05 | 30 | 106,480 | 30.79 | 85.12 |
| 17 | S. enterica | 603 | 68,922.61 | 5.45 | 30 | 136,750 | 30.39 | 77.78 |
| 18 | S. caeli | 597 | 67,893.91 | 4.91 | 30 | 92,960 | 30.87 | 81.76 |
| 19 | S. haemolyticus | 601 | 68,750.54 | 5.46 | 30 | 92,835 | 32.41 | 81.20 |
| Bacteria | Sub-Cellular Localization | ||||
|---|---|---|---|---|---|
| Extracellular | Outer Membrane | Periplasmic Space | Inner Membrane | Cytoplasm | |
| Bacteria associated with normal individuals | |||||
| L. rhamnosus | 0.176 | 0.779 | 1.619 | 0.433 | 3.993 |
| R. intestinalis | 0.091 | 0.057 | 0.525 | 0.148 | 6.180 |
| C. comes | 1.146 | 1.249 | 0.448 | 0.396 | 3.761 |
| F. prausnitzii 1 | 0.111 | 0.038 | 2.304 | 0.365 | 4.183 |
| F. prausnitzii 2 | 0.084 | 0.126 | 0.579 | 1.541 | 4.669 |
| F. prausnitzii 3 | 0.857 | 0.311 | 0.713 | 0.530 | 4.589 |
| P. acnes | 0.030 | 0.010 | 0.232 | 0.230 | 6.498 |
| Bacteria associated with breast cancer patients | |||||
| M. bacterium | 0.298 | 0.112 | 0.360 | 3.839 | 2.390 |
| S. xylosus | 0.050 | 0.054 | 0.174 | 0.117 | 6.604 |
| S. suis | 0.063 | 0.089 | 0.169 | 0.229 | 6.449 |
| Bacillus sp. M4U3P1 | 0.004 | 0.008 | 0.038 | 0.049 | 6.901 |
| E. coli (strain K12) | 0.019 | 0.012 | 0.353 | 0.090 | 6.526 |
| S. aquatilis NBRC 16722 | 0.161 | 0.389 | 0.440 | 3.271 | 2.738 |
| C. amalonaticus | 0.043 | 0.036 | 0.083 | 0.857 | 5.982 |
| E. gallinarum 1 | 0.052 | 0.041 | 0.026 | 0.066 | 6.815 |
| E. gallinarum 2 | 0.011 | 0.008 | 0.021 | 0.074 | 6.885 |
| S. enterica | 0.008 | 0.007 | 0.081 | 0.038 | 6.865 |
| S. caeli | 0.108 | 0.152 | 0.296 | 0.102 | 6.343 |
| S. haemolyticus | 0.082 | 0.044 | 0.236 | 0.131 | 6.508 |
| Probiotics | α Helix (%) | Extended Strand (%) | β Turn (%) | Random Coil (%) |
|---|---|---|---|---|
| Normal tissue associated bacteria | ||||
| L. rhamnosus | 26.37 | 23.55 | 7.13 | 42.95 |
| R. intestinalis | 27.33 | 22.67 | 6.50 | 43.50 |
| C. comes | 23.02 | 25.91 | 8.30 | 42.77 |
| F. prausnitzii 1 | 27.42 | 22.74 | 6.69 | 43.14 |
| F. prausnitzii 2 | 26.15 | 22.04 | 6.25 | 45.56 |
| F. prausnitzii 3 | 23.32 | 25.82 | 8.29 | 42.57 |
| P. acnes | 26.60 | 24.75 | 6.90 | 41.75 |
| Breast cancer patients associated bacteria | ||||
| M. bacterium | 26.45 | 23.21 | 6.31 | 44.03 |
| S. xylosus | 26.80 | 23.95 | 6.87 | 42.38 |
| S. suis | 26.88 | 22.37 | 6.84 | 43.91 |
| Bacillus sp. M4U3P1 | 27.32 | 22.52 | 6.62 | 43.54 |
| E. coli (strain K12) | 25.70 | 23.88 | 7.13 | 43.28 |
| S. aquatilis NBRC 16722 | 22.66 | 24.25 | 5.39 | 47.70 |
| C. amalonaticus | 26.79 | 24.13 | 5.49 | 43.59 |
| E. gallinarum 1 | 25.17 | 23.49 | 6.21 | 45.13 |
| E. gallinarum 2 | 27.24 | 23.59 | 7.14 | 42.03 |
| S. enterica | 26.37 | 24.21 | 6.63 | 42.79 |
| S. caeli | 27.81 | 23.62 | 6.70 | 41.88 |
| S. haemolyticus | 26.46 | 24.13 | 6.99 | 42.43 |
| Bacteria | Relevance Score (%) | Druggability | Bottleneck Radius (Angstrom) | Length (Angstrom) | Curvature |
|---|---|---|---|---|---|
| Normal tissue associated microbiota | |||||
| L. rhamnosus | 100 | 0.14 | 2.3 | 1.4 | 1.1 |
| R. intestinalis | 100 | 0.17 | 1.6 | 4.3 | 1.2 |
| C. comes | 100 | 0.28 | 1.9 | 2.7 | 1.3 |
| F. prausnitzii 1 | 100 | 0.07 | 2.1 | 2.5 | 1.0 |
| F. prausnitzii 2 | 100 | 0.07 | 1.6 | 8.8 | 1.1 |
| F. prausnitzii 3 | 100 | 0.05 | 1.2 | 10.7 | 1.4 |
| P. acnes | 100 | 0.45 | 1.2 | 10.7 | 1.4 |
| Breast cancer patients associated bacteria | |||||
| Methylococcaceae bacterium | 100 | 0.80 | 2.3 | 6.9 | 1.1 |
| S. xylosus | 100 | 0.60 | 2.0 | 5.7 | 1.1 |
| S. suis | 100 | 0.07 | 2.1 | 1.5 | 1.0 |
| Bacillus sp. M4U3P1 | 100 | 0.07 | 2.2 | 4.4 | 1.2 |
| E. coli (strain K12) | 100 | 0.06 | 2.2 | 1.9 | 1.2 |
| S. aquatilis NBRC 16722 | 100 | 0.06 | 1.1 | 8.7 | 0.65 |
| C. amalonaticus | 100 | 0.14 | 1.3 | 14.6 | 1.2 |
| E. gallinarum 1 | 100 | 0.13 | 1.2 | 15.9 | 1.3 |
| E. gallinarum 2 | 100 | 0.23 | 2.8 | 2.0 | 1.2 |
| S. enterica | 100 | 0.51 | 2.5 | 1.5 | 1.0 |
| S. caeli | 100 | 0.39 | 2.3 | 4.0 | 1.1 |
| S. haemolyticus | 100 | 0.52 | 1.9 | 3.4 | 1.0 |
| Bacteria | Residues in Most Favored Region (%) | Residues in Additional Allowed Regions (%) | Residues in Generously Allowed Regions (%) | Residues in Disallowed Regions (%) | G-Factors | ||
|---|---|---|---|---|---|---|---|
| Dihedrals | Covalent | Overall | |||||
| Normal tissue associated bacteria | |||||||
| L. rhamnosus | 88.4 | 10.6 | 0.8 | 0.2 | −0.13 | 0.47 | 0.11 |
| R. intestinalis | 85.2 | 14 | 0.2 | 0.6 | 0.07 | 0.42 | 0.22 |
| C. comes | 88.2 | 10.0 | 1.1 | 0.6 | −0.11 | 0.28 | 0.05 |
| F. prausnitzii 1 | 86.3 | 13.3 | 0.2 | 0.2 | 0.07 | 0.48 | 0.24 |
| F. prausnitzii 2 | 91.3 | 7.9 | 0.6 | 0.2 | −0.10 | 0.49 | 0.14 |
| F. prausnitzii 3 | 81.7 | 16.7 | 0.9 | 0.7 | −0.26 | 0.37 | 0.00 |
| P. acnes | 82.1 | 16.9 | 0.4 | 0.6 | −0.23 | 0.42 | 0.03 |
| Breast cancer patients associated bacteria | |||||||
| Methylococcaceae bacterium | 88.2 | 10 | 1.0 | 0.8 | −0.14 | 0.37 | 0.07 |
| S. xylosus | 86.8 | 11.5 | 0.9 | 0.8 | −0.05 | 0.57 | 0.20 |
| S. suis | 87.5 | 11.7 | 0.6 | 0.2 | −0.00 | 0.62 | 0.24 |
| Bacillus sp. M4U3P1 | 87.4 | 11.7 | 0.6 | 0.4 | −0.04 | 0.57 | 0.20 |
| E. coli (strain K12) | 87.3 | 11.3 | 0.9 | 0.4 | −0.11 | 0.24 | 0.04 |
| S. aquatilis NBRC 16722 | 83.6 | 14.6 | 1.4 | 0.4 | −0.32 | −0.06 | −0.21 |
| C. amalonaticus | 86.4 | 12.0 | 1.1 | 0.4 | −0.14 | 0.47 | 0.10 |
| E. gallinarum 1 | 84.9 | 10.2 | 3.1 | 1.8 | −0.08 | 0.31 | 0.08 |
| E. gallinarum 2 | 86.5 | 11.3 | 1.3 | 0.9 | −0.02 | 0.57 | 0.21 |
| S. enterica | 86.6 | 11.3 | 1.3 | 0.9 | −0.02 | 0.57 | 0.21 |
| S. caeli | 86.6 | 11.7 | 1.1 | 0.6 | −0.12 | 0.25 | 0.04 |
| S. haemolyticus | 85.9 | 12.1 | 0.9 | 1.1 | −0.05 | 0.55 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muccee, F.; Ghazanfar, S.; Ajmal, W.; Al-Zahrani, M. In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients. Genes 2022, 13, 1545. https://doi.org/10.3390/genes13091545
Muccee F, Ghazanfar S, Ajmal W, Al-Zahrani M. In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients. Genes. 2022; 13(9):1545. https://doi.org/10.3390/genes13091545
Chicago/Turabian StyleMuccee, Fatima, Shakira Ghazanfar, Wajya Ajmal, and Majid Al-Zahrani. 2022. "In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients" Genes 13, no. 9: 1545. https://doi.org/10.3390/genes13091545
APA StyleMuccee, F., Ghazanfar, S., Ajmal, W., & Al-Zahrani, M. (2022). In-Silico Characterization of Estrogen Reactivating β-Glucuronidase Enzyme in GIT Associated Microbiota of Normal Human and Breast Cancer Patients. Genes, 13(9), 1545. https://doi.org/10.3390/genes13091545