
ASPN Is a Potential Biomarker and Associated with Immune Infiltration in Endometriosis


Abstract
:1. Introduction
2. Materials and Methods
2.1. Patients and Tissue Samples
2.2. Data Download
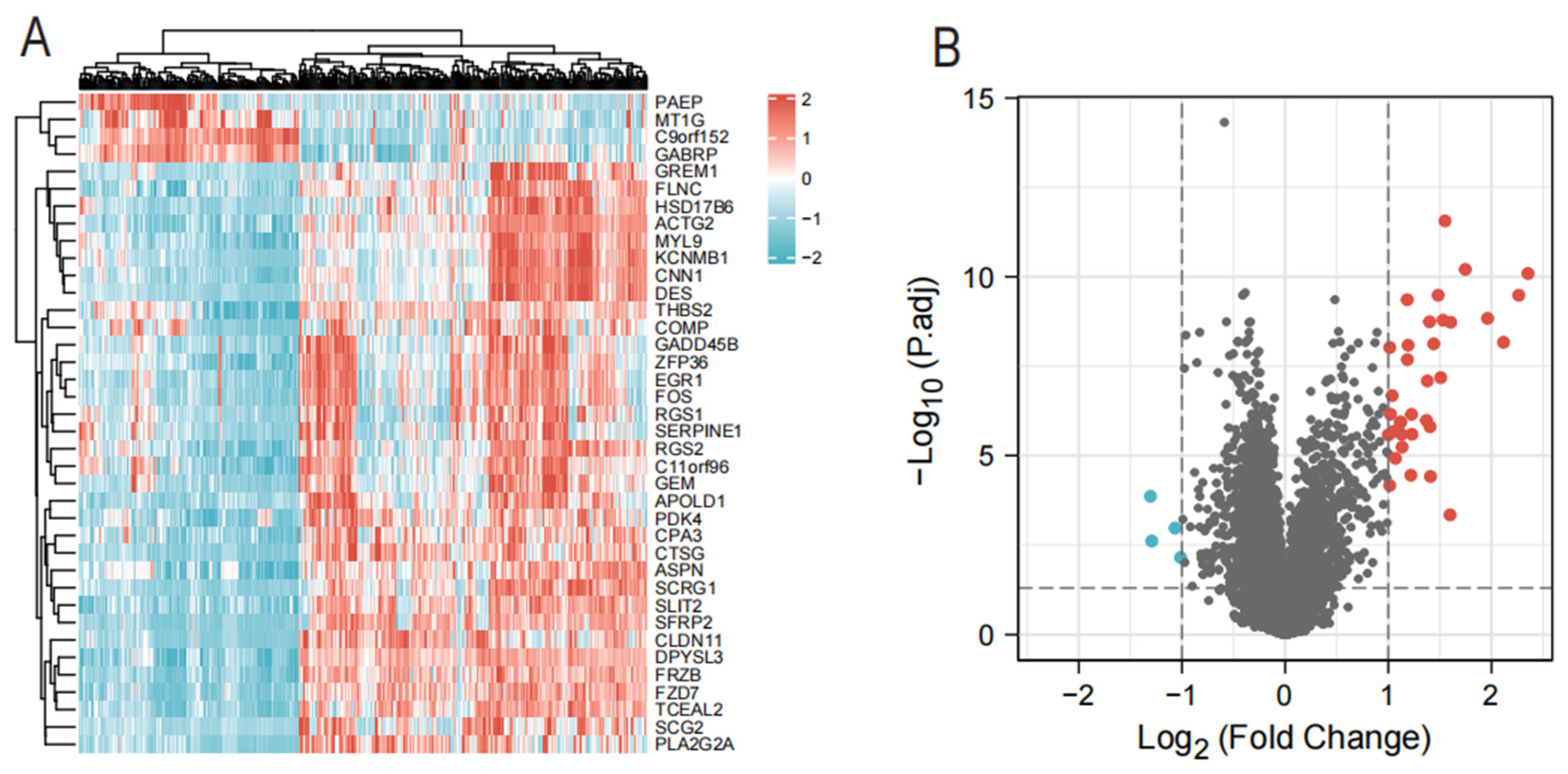
2.3. Identification of Differentially Expressed Endometriosis Genes
2.4. Candidate Diagnostic Biomarker Screening
2.5. Quantitative Real-Time PCR (qRT-PCR)
2.6. Western Blotting Experiments
2.7. Functional Enrichment Analysis
2.8. Immune Infiltration Level Analysis
2.9. Statistical Analysis
3. Results
3.1. Identification of DEGs in Endometriosis
3.2. Screening Validation of Diagnostic Markers
3.3. Sensitivity and Specificity of ASPN in Internal and External Datasets
3.4. Correlation of ASPN Expression with Clinical Features in Patients with Endometriosis
3.5. Functional Annotation of DEGs and GSEA
3.6. ASPN Is Associated with Immune Infiltration in Endometriosis
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Christ, J.P.; Yu, O.; Schulze-Rath, R.; Grafton, J.; Hansen, K.; Reed, S.D. Incidence, prevalence, and trends in endometriosis diagnosis: A United States population-based study from 2006 to 2015. Am. J. Obstet. Gynecol. 2021, 225, 500.e501–500.e509. [Google Scholar] [CrossRef] [PubMed]
- Koninckx, P.R.; Ussia, A.; Adamyan, L.; Tahlak, M.; Keckstein, J.; Wattiez, A.; Martin, D.C. The epidemiology of endometriosis is poorly known as the pathophysiology and diagnosis are unclear. Best Pract. Res. Clin. Obstet. Gynaecol. 2021, 71, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Gratton, S.M.; Choudhry, A.J.; Vilos, G.A.; Vilos, A.; Baier, K.; Holubeshen, S.; Medor, M.C.; Mercier, S.; Nguyen, V.; Chen, I. Diagnosis of Endometriosis at Laparoscopy: A Validation Study Comparing Surgeon Visualization with Histologic Findings. J. Obstet. Gynaecol. Can. JOGC = J. D’obstetrique Et Gynecol. Du Can. JOGC 2022, 44, 135–141. [Google Scholar] [CrossRef] [PubMed]
- Pascoal, E.; Wessels, J.M.; Aas-Eng, M.K.; Abrao, M.S.; Condous, G.; Jurkovic, D.; Espada, M.; Exacoustos, C.; Ferrero, S.; Guerriero, S.; et al. Strengths and limitations of diagnostic tools for endometriosis and relevance in diagnostic test accuracy research. Ultrasound Obstet. Gynecol. Off. J. Int. Soc. Ultrasound Obstet. Gynecol. 2022; online ahead of print. [Google Scholar]
- Kido, A.; Himoto, Y.; Moribata, Y.; Kurata, Y.; Nakamoto, Y. MRI in the Diagnosis of Endometriosis and Related Diseases. Korean J. Radiol. 2022, 23, 426–445. [Google Scholar] [CrossRef]
- Wu, X.G.; Chen, J.J.; Zhou, H.L.; Wu, Y.; Lin, F.; Shi, J.; Wu, H.Z.; Xiao, H.Q.; Wang, W. Identification and Validation of the Signatures of Infiltrating Immune Cells in the Eutopic Endometrium Endometria of Women with Endometriosis. Front. Immunol. 2021, 12, 671201. [Google Scholar] [CrossRef] [PubMed]
- Vallvé-Juanico, J.; Santamaria, X.; Vo, K.C.; Houshdaran, S.; Giudice, L.C. Macrophages display proinflammatory phenotypes in the eutopic endometrium of women with endometriosis with relevance to an infectious etiology of the disease. Fertil. Steril. 2019, 112, 1118–1128. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Bhandari, R.; Lei, Q.; Lu, M.; Zhang, L.; Zhang, M.; Sun, F.; Feng, L.; Zhao, S. Identification and Exploration of Novel Macrophage M2-Related Biomarkers and Potential Therapeutic Agents in Endometriosis. Front. Mol. Biosci. 2021, 8, 656145. [Google Scholar] [CrossRef] [PubMed]
- Vallve-Juanico, J.; Houshdaran, S.; Giudice, L.C. The endometrial immune environment of women with endometriosis. Hum. Reprod. Update 2019, 25, 564–591. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Q.; Yang, F.; Chen, X.; Li, J.; Zhong, C.; Chen, S. Patterns of Immune Infiltration in Endometriosis and Their Relationship to r-AFS Stages. Front. Genet. 2021, 12, 631715. [Google Scholar] [CrossRef] [PubMed]
- Yuan, W.H.; Xie, Q.Q.; Wang, K.P.; Shen, W.; Feng, X.F.; Liu, Z.; Shi, J.T.; Zhang, X.B.; Zhang, K.; Deng, Y.J.; et al. Screening of osteoarthritis diagnostic markers based on immune-related genes and immune infiltration. Sci. Rep. 2021, 11, 7032. [Google Scholar] [CrossRef]
- Su, R.; Jin, C.; Zhou, L.; Cao, Y.; Kuang, M.; Li, L.; Xiang, J. Construction of a ceRNA network of hub genes affecting immune infiltration in ovarian cancer identified by WGCNA. BMC Cancer 2021, 21, 970. [Google Scholar] [CrossRef] [PubMed]
- Hever, A.; Roth, R.B.; Hevezi, P.; Marin, M.E.; Acosta, J.A.; Acosta, H.; Rojas, J.; Herrera, R.; Grigoriadis, D.; White, E.; et al. Human endometriosis is associated with plasma cells and overexpression of B lymphocyte stimulator. Proc. Natl. Acad. Sci. USA 2007, 104, 12451–12456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gabriel, M.; Fey, V.; Heinosalo, T.; Adhikari, P.; Rytkönen, K.; Komulainen, T.; Huhtinen, K.; Laajala, T.D.; Siitari, H.; Virkki, A.; et al. A relational database to identify differentially expressed genes in the endometrium and endometriosis lesions. Sci. Data 2020, 7, 284. [Google Scholar] [CrossRef]
- Barrett, T.; Wilhite, S.E.; Ledoux, P.; Evangelista, C.; Kim, I.F.; Tomashevsky, M.; Marshall, K.A.; Phillippy, K.H.; Sherman, P.M.; Holko, M.; et al. NCBI GEO: Archive for functional genomics data sets--update. Nucleic Acids Res. 2013, 41, D991–D995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ritchie, M.E.; Phipson, B.; Wu, D.; Hu, Y.; Law, C.W.; Shi, W.; Smyth, G.K. limma powers differential expression analyses for RNA-sequencing and microarray studies. Nucleic Acids Res. 2015, 43, e47. [Google Scholar] [CrossRef] [PubMed]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Becht, E.; Giraldo, N.A.; Lacroix, L.; Buttard, B.; Elarouci, N.; Petitprez, F.; Selves, J.; Laurent-Puig, P.; Sautès-Fridman, C.; Fridman, W.H.; et al. Estimating the population abundance of tissue-infiltrating immune and stromal cell populations using gene expression. Genome Biol. 2016, 17, 218. [Google Scholar] [CrossRef]
- Yang, Z.; Zi, Q.; Xu, K.; Wang, C.; Chi, Q. Development of a macrophages-related 4-gene signature and nomogram for the overall survival prediction of hepatocellular carcinoma based on WGCNA and LASSO algorithm. Int. Immunopharmacol. 2021, 90, 107238. [Google Scholar] [CrossRef]
- Wu, Z.; Lu, Z.; Li, L.; Ma, M.; Long, F.; Wu, R.; Huang, L.; Chou, J.; Yang, K.; Zhang, Y.; et al. Identification and Validation of Ferroptosis-Related LncRNA Signatures as a Novel Prognostic Model for Colon Cancer. Front. Immunol. 2021, 12, 783362. [Google Scholar] [CrossRef]
- Venkatesh, S.S.; Ferreira, T.; Benonisdottir, S.; Rahmioglu, N.; Becker, C.M.; Granne, I.; Zondervan, K.T.; Holmes, M.V.; Lindgren, C.M.; Wittemans, L.B.L. Obesity and risk of female reproductive conditions: A Mendelian randomisation study. PLoS Med. 2022, 19, e1003679. [Google Scholar] [CrossRef]
- Smolarz, B.; Szyllo, K.; Romanowicz, H. Endometriosis: Epidemiology, Classification, Pathogenesis, Treatment and Genetics (Review of Literature). Int. J. Mol. Sci. 2021, 22, 19. [Google Scholar]
- Ma, J.; Zhang, L.; Zhan, H.; Mo, Y.; Ren, Z.; Shao, A.; Lin, J. Single-cell transcriptomic analysis of endometriosis provides insights into fibroblast fates and immune cell heterogeneity. Cell Biosci. 2021, 11, 125. [Google Scholar] [CrossRef] [PubMed]
- Hughes, R.M.; Simons, B.W.; Khan, H.; Miller, R.; Kugler, V.; Torquato, S.; Theodros, D.; Haffner, M.C.; Lotan, T.; Huang, J.; et al. Asporin Restricts Mesenchymal Stromal Cell Differentiation, Alters the Tumor Microenvironment, and Drives Metastatic Progression. Cancer Res. 2019, 79, 3636–3650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castellana, B.; Escuin, D.; Peiró, G.; Garcia-Valdecasas, B.; Vázquez, T.; Pons, C.; Pérez-Olabarria, M.; Barnadas, A.; Lerma, E. ASPN and GJB2 Are Implicated in the Mechanisms of Invasion of Ductal Breast Carcinomas. J. Cancer 2012, 3, 175–183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rochette, A.; Boufaied, N.; Scarlata, E.; Hamel, L.; Brimo, F.; Whitaker, H.C.; Ramos-Montoya, A.; Neal, D.E.; Dragomir, A.; Aprikian, A.; et al. Asporin is a stromally expressed marker associated with prostate cancer progression. Br. J. Cancer 2017, 116, 775–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basak, D.; Jamal, Z.; Ghosh, A.; Mondal, P.K.; Dey Talukdar, P.; Ghosh, S.; Ghosh Roy, B.; Ghosh, R.; Halder, A.; Chowdhury, A.; et al. Reciprocal interplay between asporin and decorin: Implications in gastric cancer prognosis. PLoS ONE 2021, 16, e0255915. [Google Scholar] [CrossRef]
- Mishra, A.; Awasthi, S.; Raj, S.; Mishra, P.; Srivastava, R.N. Identifying the role of ASPN and COMP genes in knee osteoarthritis development. J. Orthop. Surg. Res. 2019, 14, 337. [Google Scholar] [CrossRef]
- Li, H.; Yang, H.H.; Sun, Z.G.; Tang, H.B.; Min, J.K. Whole-transcriptome sequencing of knee joint cartilage from osteoarthritis patients. Bone Jt. Res. 2019, 8, 290–303. [Google Scholar] [CrossRef]
- Li, H.; Zhang, Z.; Chen, L.; Sun, X.; Zhao, Y.; Guo, Q.; Zhu, S.; Li, P.; Min, L.; Zhang, S. Cytoplasmic Asporin promotes cell migration by regulating TGF-β/Smad2/3 pathway and indicates a poor prognosis in colorectal cancer. Cell Death Dis. 2019, 10, 109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.; Wang, L.; Guan, X.; Yue, C. MiR-4303 relieves chondrocyte inflammation by targeting ASPN in osteoarthritis. J. Orthop. Surg. Res. 2021, 16, 618. [Google Scholar] [CrossRef] [PubMed]
- Sekimoto, T.; Ishii, M.; Emi, M.; Kurogi, S.; Funamoto, T.; Yonezawa, Y.; Tajima, T.; Sakamoto, T.; Hamada, H.; Chosa, E. Copy number loss in the region of the ASPN gene in patients with acetabular dysplasia: ASPN CNV in acetabular dysplasia. Bone Jt. Res. 2017, 6, 439–445. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Lv, X.; Yu, H.; Xu, P.; Ma, R.; Zou, K. In search of key genes associated with endometriosis using bioinformatics approach. Eur. J. Obstet. Gynecol. Reprod. Biol. 2015, 194, 119–124. [Google Scholar] [CrossRef]
- Wu, T.; Zhang, R.; Jiang, Q.; Li, Z.; Wu, R. Expression of cellular adherent and invasive molecules in recurrent ovarian endometriosis. J. Int. Med. Res. 2020, 48, 300060520971993. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jiang, R.; Yao, Y.; Qian, L.; Zhao, Y.; Huang, X. Identification of endometriosis-associated genes and pathways based on bioinformatic analysis. Medicine 2021, 100, e26530. [Google Scholar] [CrossRef]
- Khan, K.N.; Yamamoto, K.; Fujishita, A.; Muto, H.; Koshiba, A.; Kuroboshi, H.; Saito, S.; Teramukai, S.; Nakashima, M.; Kitawaki, J. Differential Levels of Regulatory T Cells and T-Helper-17 Cells in Women with Early and Advanced Endometriosis. J. Clin. Endocrinol. Metab. 2019, 104, 4715–4729. [Google Scholar] [CrossRef]
- Chen, S.; Chai, X.; Wu, X. Bioinformatical analysis of the key differentially expressed genes and associations with immune cell infiltration in development of endometriosis. BMC Genom. Data 2022, 23, 20. [Google Scholar] [CrossRef] [PubMed]
- Riccio, L.G.C.; Baracat, E.C.; Chapron, C.; Batteux, F.; Abrão, M.S. The role of the B lymphocytes in endometriosis: A systematic review. J. Reprod. Immunol. 2017, 123, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Freitag, N.; Pour, S.J.; Fehm, T.N.; Toth, B.; Markert, U.R.; Weber, M.; Togawa, R.; Kruessel, J.S.; Baston-Buest, D.M.; Bielfeld, A.P. Are uterine natural killer and plasma cells in infertility patients associated with endometriosis, repeated implantation failure, or recurrent pregnancy loss? Arch. Gynecol. Obs. 2020, 302, 1487–1494. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Characteristics | Number of Cases | |
|---|---|---|
| Age (years) | ≤35 | 241 |
| >35 | 160 | |
| Stage | ||
| I–II | 85 | |
| III–IV | 250 | |
| Healthy | 66 | |
| Cycle Phase | ||
| menstruation | 20 | |
| proliferative | 60 | |
| secretory | 97 | |
| medication | 175 | |
| unknown | 49 | |
| Site | ||
| endometrium | 146 | |
| endometriosis | 166 | |
| DIE | 89 |
| Gene Names | HR (95% CI) | p-Value |
|---|---|---|
| SCG2 | 7.89 (4.80–11.74) | <0.001 |
| EGR1 | 3.70 (2.10–5.82) | <0.001 |
| ASPN | 1.58 (1.04–2.32) | 0.038 |
| SLIT2 | 3.07 (1.80–4.76) | <0.001 |
| GEM | 0.17 (0.03–0.63) | 0.008 |
| SFRP2 | 0.34 (0.15–0.65) | 0.001 |
| FOS | 0.24 (0.08–0.56) | 0.001 |
| GREM1 | 0.32 (0.09–0.90) | 0.035 |
| C9orf152 | 2.01 (1.31–2.94) | 0.003 |
| PAEP | 0.69 (0.51–0.89) | 0.007 |
| Dataset | Number | AUC (%) | Yoden Index | Sensitivity (%) | Specificity (%) |
|---|---|---|---|---|---|
| GSE141549 | 401 | 73.8 | 0.43 | 77.3 | 65.2 |
| GSE7305 | 20 | 93.0 | 0.80 | 90.0 | 90.0 |
| Clinical Data | 24 | 83.3 | 0.58 | 91.7 | 66.7 |
| ASPN | ||||
|---|---|---|---|---|
| Clinical Parameters | n (%) | High Group (%) | Low Group (%) | p-Value |
| Age (years) | 32.24 ± 6.694 | 33.60 ± 7.387 | 0.030 | |
| Stage | ||||
| I–II | 85 (21.2) | 45 (11.2) | 40 (10.0) | 0.006 |
| III–IV | 250 (62.3) | 134 (33.4) | 116 (28.9) | |
| Healthy | 66 (16.5) | 21 (5.2) | 45 (11.2) | |
| Cycle Phase | ||||
| menstruation | 60 (15.0) | 29 (7.3) | 31 (7.7) | 0.227 |
| proliferative | 97 (24.2) | 42 (10.5) | 55 (13.7) | |
| secretory | 20 (5.0) | 14 (3.5) | 6 (1.5) | |
| medication | 175 (43.6) | 92 (22.9) | 83 (20.7) | |
| unknown | 49 (12.2) | 23 (5.7) | 26 (6.5) | |
| Site | ||||
| Endometrium | 146 (36.4) | 22 (5.5) | 124 (30.9) | <0.001 |
| Peritoneum | 27 (6.7) | 4 (1.0) | 23 (5.7) | |
| Endometriosis | 139 (34.7) | 100 (24.9) | 39 (9.7) | |
| DIE | 89 (22.2) | 74 (18.5) | 15 (3.7) | |
| Univariate-Logistic | Multivariate-Logistic | |||
|---|---|---|---|---|
| Clinical Parameters | HR (95% CI) | p-Value | HR (95% CI) | p-Value |
| Age (years) | 0.842 (0.802–0.884) | <0.001 | 0.848 (0.805–0.894) | <0.001 |
| Cycle Phase | 1.077 (0.884–1.313) | 0.462 | / | / |
| Site | 1.895 (1.472–2.438) | <0.001 | 1.256 (0.867–1.819) | 0.229 |
| ASPN | 1.870 (1.544–2.266) | <0.001 | 1.574 (1.199–2.068) | 0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, L.; Sun, J. ASPN Is a Potential Biomarker and Associated with Immune Infiltration in Endometriosis. Genes 2022, 13, 1352. https://doi.org/10.3390/genes13081352
Wang L, Sun J. ASPN Is a Potential Biomarker and Associated with Immune Infiltration in Endometriosis. Genes. 2022; 13(8):1352. https://doi.org/10.3390/genes13081352
Chicago/Turabian StyleWang, Li, and Jing Sun. 2022. "ASPN Is a Potential Biomarker and Associated with Immune Infiltration in Endometriosis" Genes 13, no. 8: 1352. https://doi.org/10.3390/genes13081352
APA StyleWang, L., & Sun, J. (2022). ASPN Is a Potential Biomarker and Associated with Immune Infiltration in Endometriosis. Genes, 13(8), 1352. https://doi.org/10.3390/genes13081352