
Identification of Male-Specific Molecular Markers by Recombination of RhoGEF10 Gene in Spotted Knifejaw (Oplegnathus punctatus)

 , ,
, ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
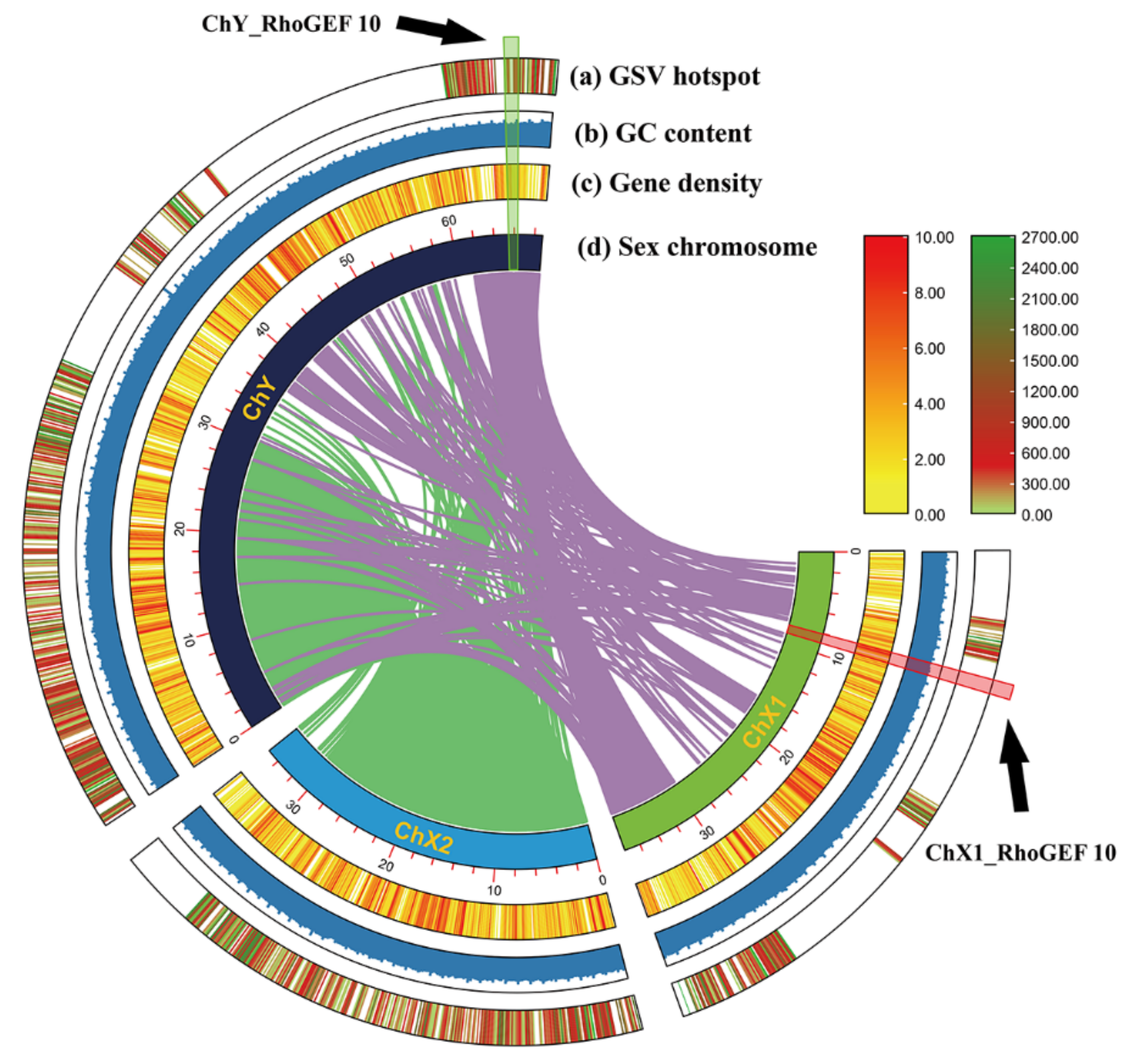
2.1. Genome-Wide Identification and Characterization of RhoGEF10
2.2. Source of Experimental Samples
2.3. Genomic DNA Extraction
2.4. Gender Molecular Markers in Spotted Knifejaw
3. Results
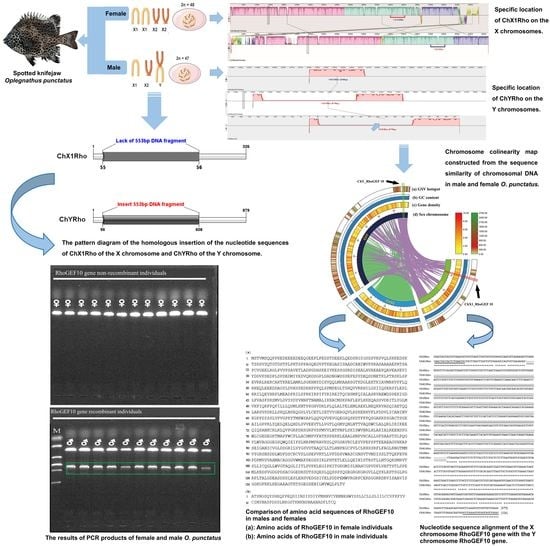
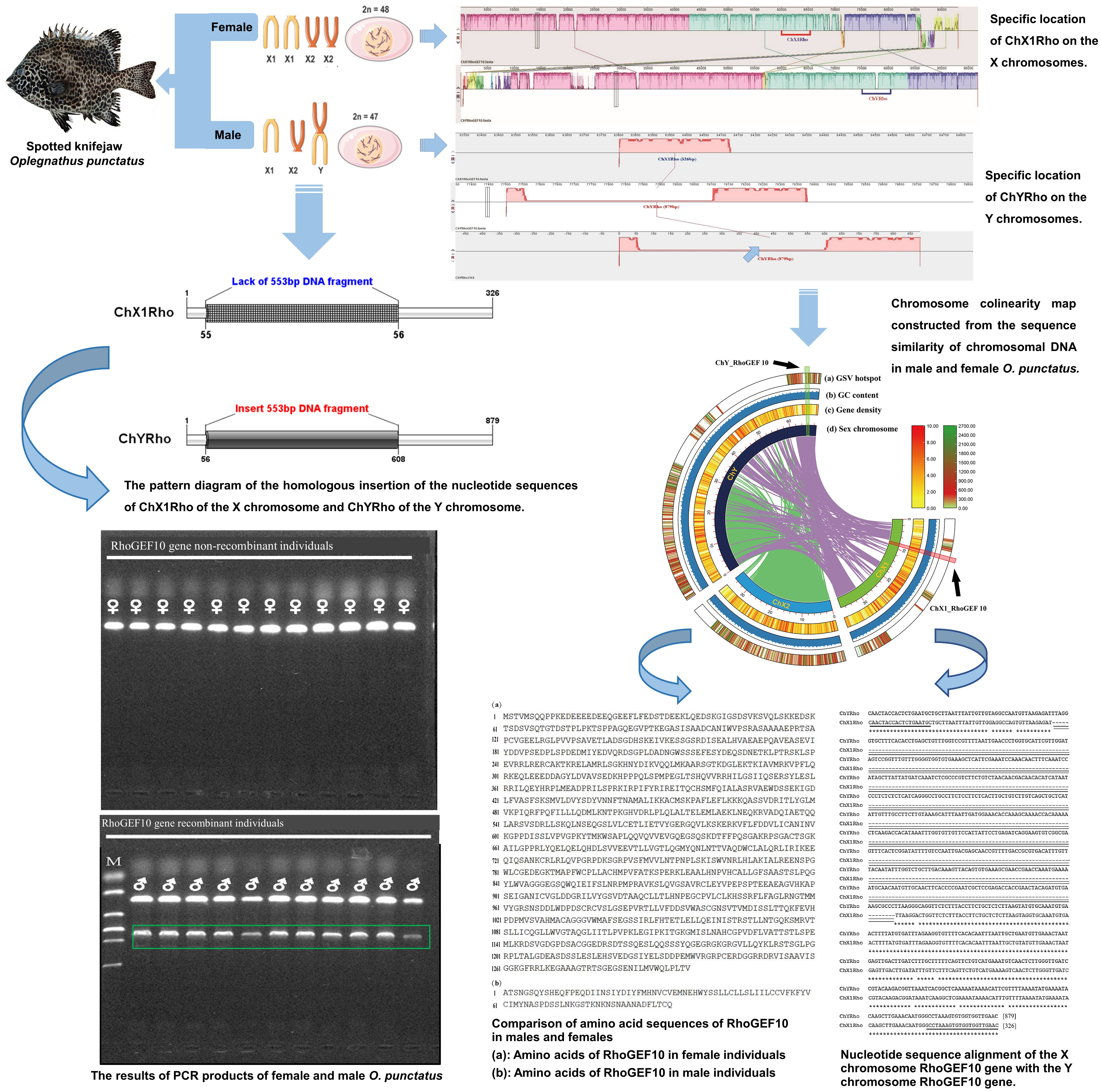
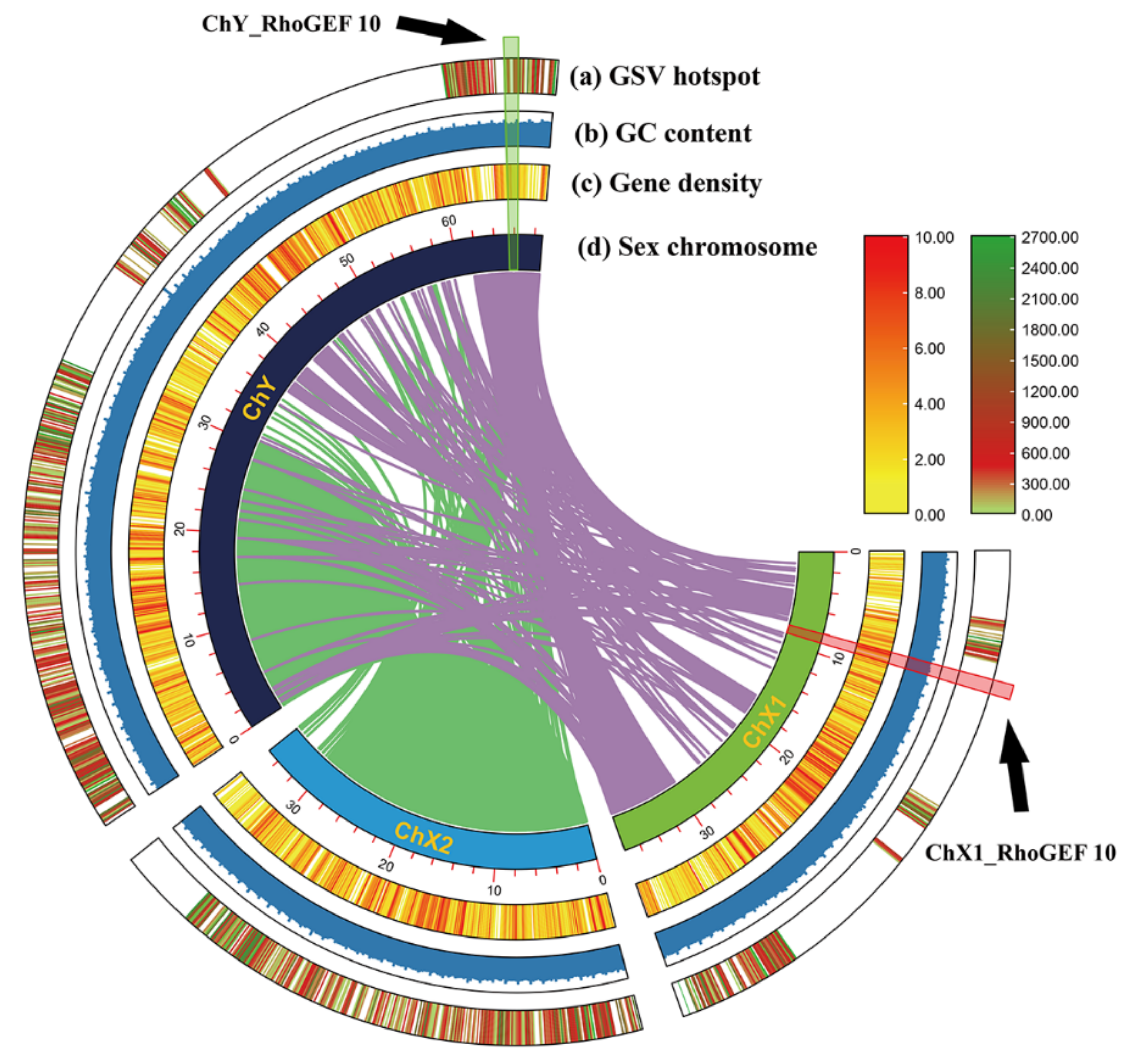
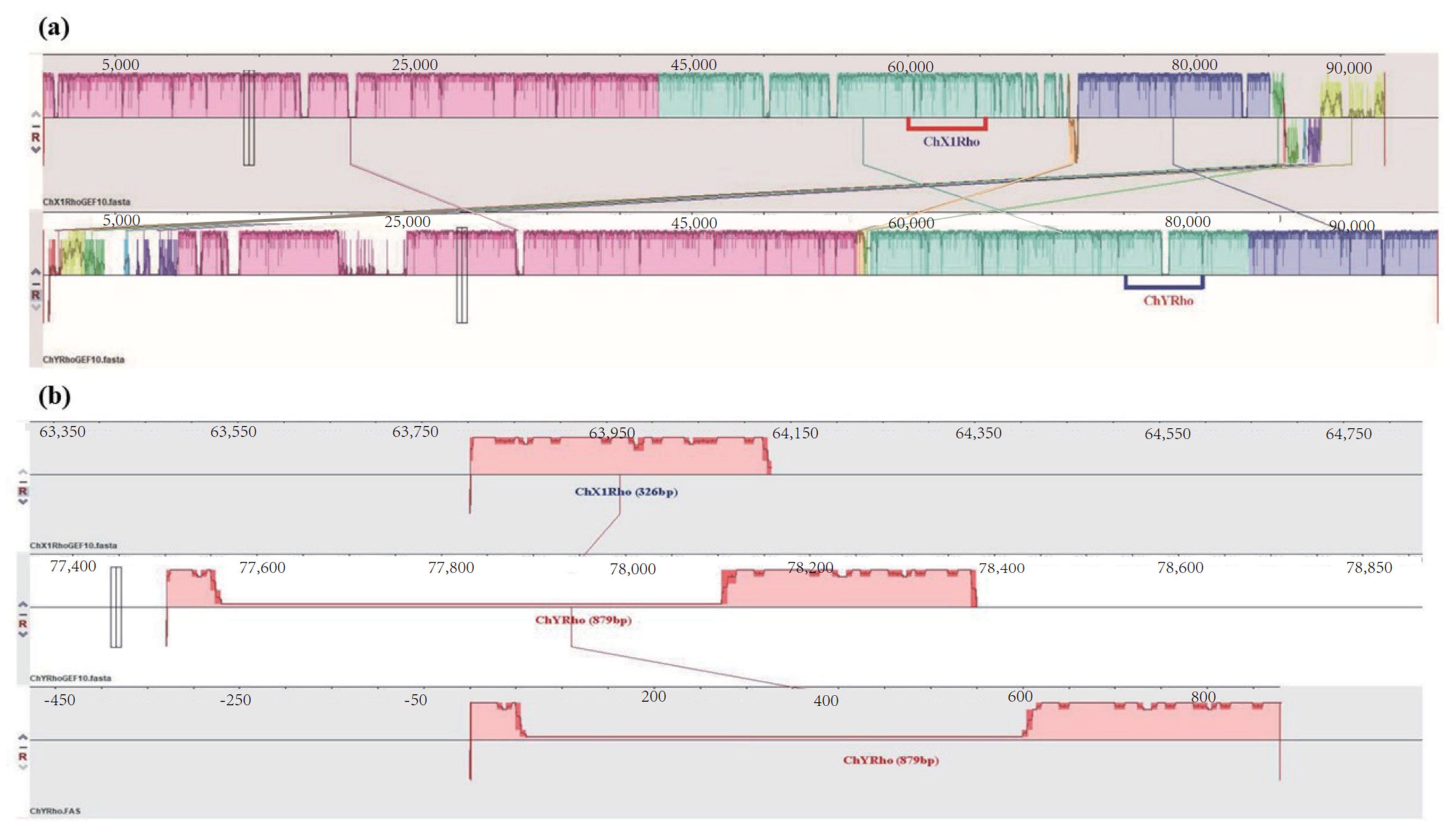
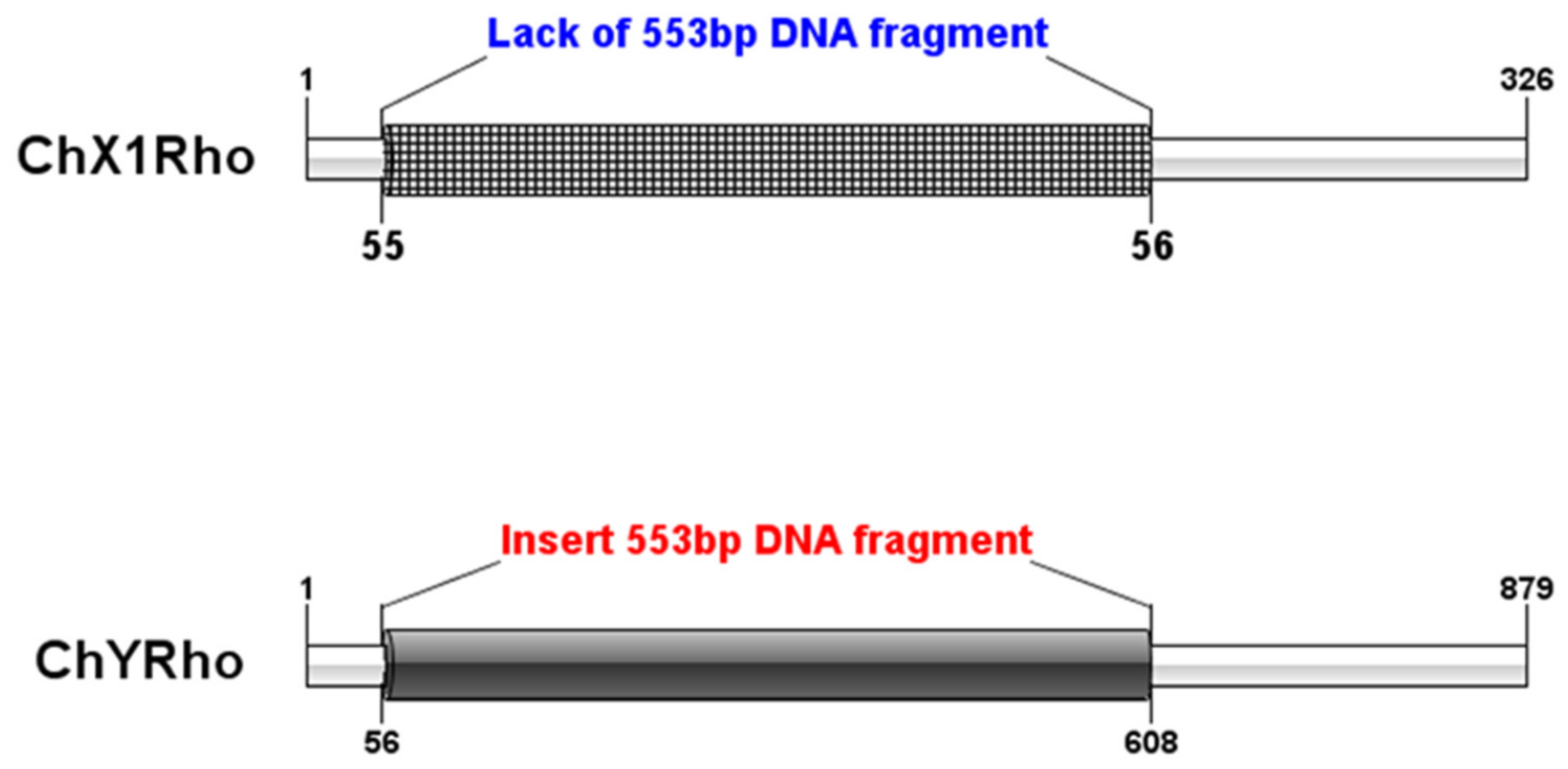
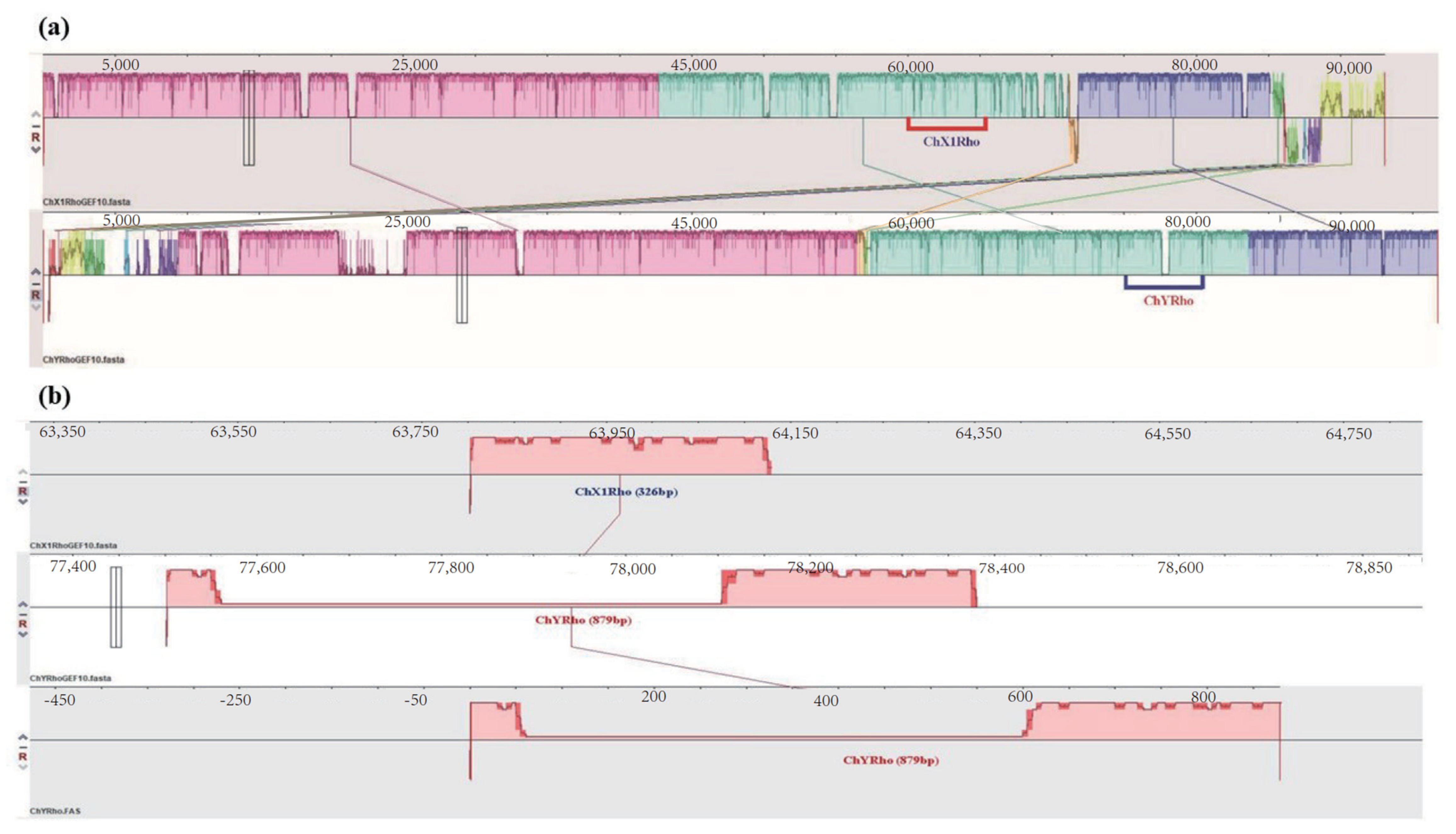
3.1. Sequence Analysis of RhoGEF10 Gene
3.2. Homologous Comparison of RhoGEF10 Gene in Males and Females
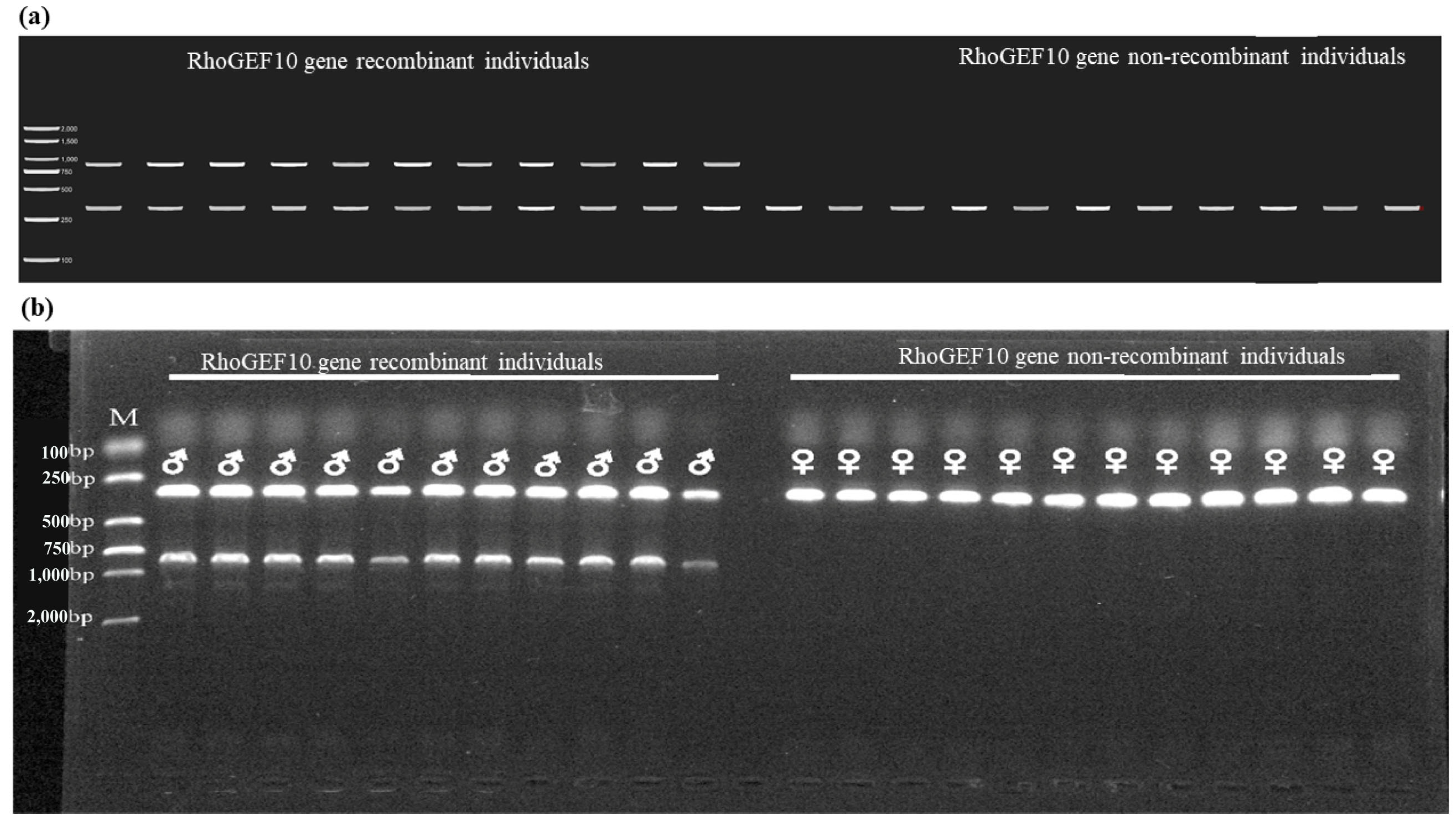
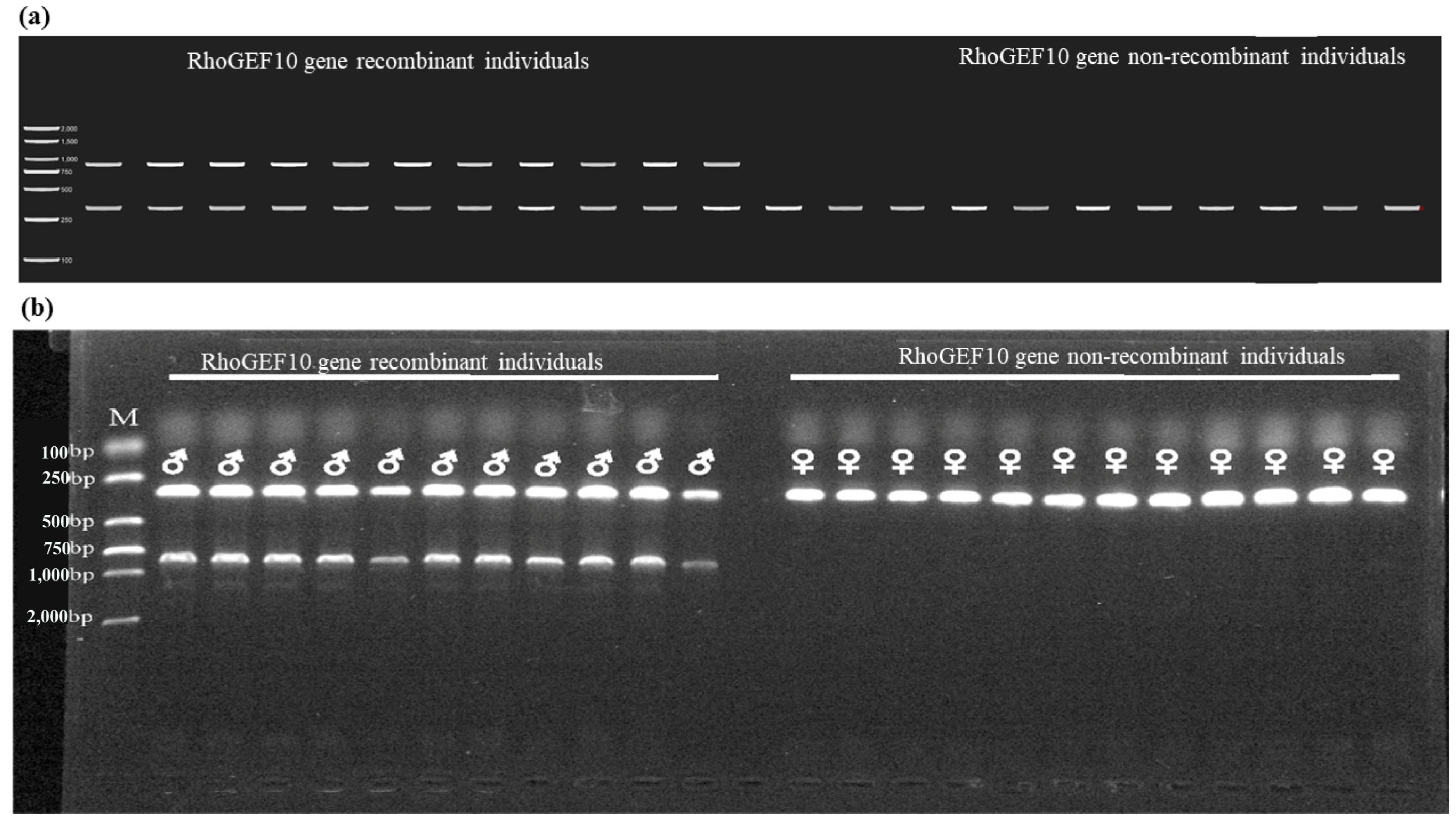
3.3. Genetic Sex Identification of New Markers in O. punctatus
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Du, X.X.; Wang, B.; Liu, X.M.; Liu, X.B.; He, Y.; Zhang, Q.Q.; Wang, X.B. Comparative transcriptome analysis of ovary and testis reveals potential sex-related genes and pathways in spotted knifejaw Oplegnathus punctatus. Gene 2017, 637, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.H.; Song, X.F.; Peng, L.; Hallerman, E.; Huang, Z.T. Effects of nitrate on aquaculture production, blood, and histological markers and liver transcriptome of Oplegnathus punctatus. Aquaculture 2019, 501, 387–396. [Google Scholar] [CrossRef]
- Kawabe, K.; Kimura, J. Rearing of spotted parrot fish Oplegnathus punctatus to market size in Chichi-jima, Ogasawara Islands, Southern Japan. Aquac. Sci. 2007, 55, 65–69. [Google Scholar]
- Xue, R.; An, H.; Liu, Q.H.; Xiao, Z.Z.; Wang, Y.F.; Li, J. Karyotype and Ag-nors in male and female of Oplegnathus punctatus. Oceanol. Limnol. Sin. 2016, 47, 626–632. [Google Scholar]
- Li, M.; Zhang, R.; Fan, G.Y.; Xu, W.T.; Zhou, Q.; Wang, L.; Li, W.S.; Pang, Z.F.; Yu, M.J.; Liu, Q.; et al. Reconstruction of the Origin of a Neo-Y Sex Chromosome and Its Evolution in the Spotted Knifejaw, Oplegnathus punctatus. Mol. Biol. Evol. 2021, 38, 2615–2626. [Google Scholar] [CrossRef]
- Xu, D.D.; Sember, A.; Zhu, Q.H.; de Oliveira, E.A.; Liehr, T.; Al-Rikabi, A.B.H.; Xiao, Z.Z.; Song, H.B.; Cioffi, M.D. Deciphering the Origin and Evolution of the X1X2Y System in Two Closely-Related Oplegnathus Species (Oplegnathidae and Centrarchiformes). Int. J. Mol. Sci. 2019, 20, 16. [Google Scholar] [CrossRef] [Green Version]
- Xiao, Z.Z. Study on Popultion Genetics and Culture Biology of Oplegandthus fasciatus. Ph.D. Thesis, Chinese Academy of Sciences University, Beijing, China, 30 November 2014. [Google Scholar]
- Mosaddeghzadeh, N.; Ahmadian, M.R. The RHO Family GTPases: Mechanisms of Regulation and Signaling. Cells 2021, 10, 24. [Google Scholar] [CrossRef]
- Chang, Y.J.; Pownall, S.; Jensen, T.E.; Mouaaz, S.; Foltz, W.; Zhou, L.; Liadis, N.; Woo, M.; Hao, Z.Y.; Dutt, P.; et al. The Rho-guanine nucleotide exchange factor PDZ-RhoGEF governs susceptibility to diet-induced obesity and type 2 diabetes. eLife 2015, 4, 24. [Google Scholar] [CrossRef]
- Felline, A.; Belmonte, L.; Raimondi, F.; Bellucci, L.; Fanelli, F. Interconnecting Flexibility, Structural Communication, and Function in RhoGEF Oncoproteins. J. Chem. Inf. Model. 2019, 59, 4300–4313. [Google Scholar] [CrossRef]
- Gai, Z.C.; Zhao, J.Q. Genome-wide analysis reveals the functional and expressional correlation between RhoGAP and RhoGEF in mouse. Genomics 2020, 112, 1694–1706. [Google Scholar] [CrossRef]
- Rossman, K.L.; Der, C.J.; Sondek, J. GEF means go: Turning on Rho GTPases with guanine nucleotide-exchange factors. Nat. Rev. Mol. Cell Biol. 2005, 6, 167–180. [Google Scholar] [CrossRef]
- Liu, M.T.; Yang, J.; Wang, Z.G. RhoGEF Family and the Role in Occurrence and Development of Diseases. Biol. Bull. 2014, 34, 63–67. [Google Scholar]
- Zheng, Y. Dbl family guanine nucleotide exchange factors. Trends Biochem. Sci. 2001, 26, 724–732. [Google Scholar] [CrossRef]
- Verhoeven, K.; De Jonghe, P.; Van de Putte, T.; Nelis, E.; Zwijsen, A.; Verpoorten, N.; De Vriendt, E.; Jacobs, A.; Van Gerwen, V.; Francis, A.; et al. Slowed conduction and thin myelination of peripheral nerves associated with mutant Rho guanine-nucleotide exchange factor 10. Am. J. Hum. Genet. 2003, 73, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Sadok, A.; Marshall, C.J. Rho GTPases: Masters of cell migration. Small GTPases 2014, 5, e29710. [Google Scholar] [CrossRef] [Green Version]
- Sandrine, E.-M.; Alan, H. Rho GTPases in cell biology. Nature 2002, 420, 629–635. [Google Scholar]
- Al-Snan, N.R.; Shabbir, S.; Baksh, S.S.; AlQerainees, M.; Haidar, M.; Messaoudi, S.A.; Bakhiet, M. Population genetics of 30 insertion/deletion polymorphisms in the Bahraini population. Sci. Rep. 2021, 11, 7. [Google Scholar] [CrossRef]
- Zhou, L.; Wang, Y.; Gui, J.F. Genetic evidence for gonochoristic reproduction in gynogenetic silver crucian carp (Carassius auratus gibelio bloch) as revealed by RAPD assays. J. Mol. Evol. 2000, 51, 498–506. [Google Scholar] [CrossRef]
- Wan, Z.Y.; Xia, J.H.; Lin, G.; Wang, L.; Lin, V.C.L.; Yue, G.H. Genome-wide methylation analysis identified sexually dimorphic methylated regions in hybrid tilapia. Sci. Rep. 2016, 6, 14. [Google Scholar] [CrossRef] [Green Version]
- Gao, D.D.; Ou, M.; Wu, Y.D.; Chen, K.C.; Liu, H.Y.; Luo, Q.; Zhao, J. Gene Cloning and Expression Analysis of Growth Hormone Gene from Blotched Snakehead (Channa maculata). J. Agric. Biotechnol. 2021, 29, 2328–2341. [Google Scholar]
- Palaiokostas, C.; Bekaert, M.; Khan, M.G.Q.; Taggart, J.B.; Gharbi, K.; McAndrew, B.J.; Penman, D.J. Mapping and Validation of the Major Sex-Determining Region in Nile Tilapia (Oreochromis niloticus L.) Using RAD Sequencing. PLoS ONE 2013, 8, 9. [Google Scholar] [CrossRef]
- Degani, G.; Tzchori, I.; Yom-Din, S.; Goldberg, D.; Jackson, K. Growth differences and growth hormone expression in male and female European eels Anguilla anguilla. Gen. Comp. Endocrinol. 2003, 134, 88–93. [Google Scholar] [CrossRef]
- Ma, Q.; Liu, S.F.; Zhuang, Z.M.; Lin, L.; Sun, Z.Z.; Liu, C.L.; Ma, H.; Su, Y.Q.; Tang, Q.S. Genomic structure, polymorphism and expression analysis of the growth hormone (GH) gene in female and male Half-smooth tongue sole (Cynoglossus semilaevis). Gene 2012, 493, 92–104. [Google Scholar] [CrossRef]
- Gong, J.; Li, B.J.; Zhao, J.; Zhou, Z.X.; Ke, Q.Z.; Zhu, Q.H.; Xu, D.D.; Zhou, T.; Xu, P. Sex-Specific Genomic Region Identification and Molecular Sex Marker Development of Rock Bream (Oplegnathus fasciatus). Mar. Biotechnol. 2022, 24, 163–173. [Google Scholar] [CrossRef]
- Liu, H.Y.; Pang, M.X.; Yu, X.M.; Zhou, Y.; Tong, J.O.; Fu, B.D. Sex-specific markers developed by next-generation sequencing confirmed an XX/XY sex determination system in bighead carp (Hypophthalmichthys nobilis) and silver carp (Hypophthalmichthys molitrix). DNA Res. 2018, 25, 257–264. [Google Scholar] [CrossRef]
- Wang, L.; Xie, N.; Shen, Y.B.; Ye, B.Q.; Yue, G.H.; Feng, X.Y. Constructing High-Density Genetic Maps and Developing Sexing Markers in Northern Snakehead (Channa argus). Mar. Biotechnol. 2019, 21, 348–358. [Google Scholar] [CrossRef]
- Lin, A.Q.; Xiao, S.J.; Xu, S.B.; Ye, K.; Lin, X.Y.; Sun, S.; Wang, Z.Y. Identification of a male-specific DNA marker in the large yellow croaker (Larimichthys crocea). Aquaculture 2017, 480, 116–122. [Google Scholar] [CrossRef]
- Xu, D.D.; Lou, B.; Xu, H.X.; Li, S.L.; Geng, Z. Isolation and Characterization of Male-Specific DNA Markers in the Rock Bream Oplegnathus fasciatus. Mar. Biotechnol. 2013, 15, 221–229. [Google Scholar] [CrossRef]
- Tsuji, T.; Ohta, Y.; Kanno, Y.; Hirose, K.; Ohashi, K.; Mizuno, K. Involvement of p114-RhoGEF and Lfc in Wnt-3a-and Dishevelled-Induced RhoA Activation and Neurite Retraction in N1E-115 Mouse Neuroblastoma Cells. Mol. Biol. Cell 2010, 21, 3590–3600. [Google Scholar] [CrossRef] [Green Version]
- Van Sanden, S.; Murton, M.; Bobrowska, A.; Rahhali, N.; Sermon, J.; Rodrigues, B.; Goff-Leggett, D.; Chouaid, C.; Sebastian, M.; Greystoke, A. Prevalence of Epidermal Growth Factor Receptor Exon 20 Insertion Mutations in Non-small-Cell Lung Cancer in Europe: A Pragmatic Literature Review and Meta-analysis. Target. Oncol. 2022, 17, 153–166. [Google Scholar] [CrossRef]
- Koga, A.; Inagaki, H.; Bessho, Y.; Hori, H. Insertion of a novel transposable element in the tyrosinase gene is responsible for an albino mutation in the medaka fish, Oryzias latipes. Mol. Gen. Genet. 1995, 249, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Lindy, A.S.; Basehore, M.J.; Munisha, M.; Williams, A.L.; Friez, M.J.; Writzl, K.; Willems, P.; Dougan, S.T. Identification of a Novel Insertion Mutation in FGFR3 That Causes Thanatophoric Dysplasia Type 1. Am. J. Med. Genet. Part A 2016, 170, 1573–1579. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, R.; Banerjee, J.; Khalili, K.; Wedegaertner, P.B. Differences in Galpha12- and Galpha13-mediated plasma membrane recruitment of p115-RhoGEF. Cell Signal. 2009, 21, 996–1006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swiercz, J.M.; Worzfeld, T.; Offermanns, S. Semaphorin 4D Signaling Requires the Recruitment of Phospholipase C gamma into the Plexin-B1 Receptor Complex. Mol. Cell. Biol. 2009, 29, 6321–6334. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Xiao, Y.; Xiao, Z.; Ma, Y.; Zhao, H.; Li, J. Identification of Male-Specific Molecular Markers by Recombination of RhoGEF10 Gene in Spotted Knifejaw (Oplegnathus punctatus). Genes 2022, 13, 1262. https://doi.org/10.3390/genes13071262
Wu Y, Xiao Y, Xiao Z, Ma Y, Zhao H, Li J. Identification of Male-Specific Molecular Markers by Recombination of RhoGEF10 Gene in Spotted Knifejaw (Oplegnathus punctatus). Genes. 2022; 13(7):1262. https://doi.org/10.3390/genes13071262
Chicago/Turabian StyleWu, Yanduo, Yongshuang Xiao, Zhizhong Xiao, Yuting Ma, Haixia Zhao, and Jun Li. 2022. "Identification of Male-Specific Molecular Markers by Recombination of RhoGEF10 Gene in Spotted Knifejaw (Oplegnathus punctatus)" Genes 13, no. 7: 1262. https://doi.org/10.3390/genes13071262
APA StyleWu, Y., Xiao, Y., Xiao, Z., Ma, Y., Zhao, H., & Li, J. (2022). Identification of Male-Specific Molecular Markers by Recombination of RhoGEF10 Gene in Spotted Knifejaw (Oplegnathus punctatus). Genes, 13(7), 1262. https://doi.org/10.3390/genes13071262