
Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma

 , , , and
, , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Analysis of miRNAs and miRNA Target Genes in HNSCC Patients
2.2. HNSCC Cell Lines
2.3. RNA Extraction and Quantitative Real-Time Reverse-Transcription PCR (qRT-PCR)
2.4. Transfection of miRNAs and siRNAs into HNSCC Cells
2.5. RIP Assay
2.6. Functional Assays of HNSCC Cells (Cell Proliferation, Migration, and Invasion Assays)
2.7. Plasmid Construction and Dual-Luciferase Reporter Assays
2.8. Immunohistochemistry
2.9. Gene Set Enrichment Analysis (GSEA)
2.10. Statistical Analysis
3. Results
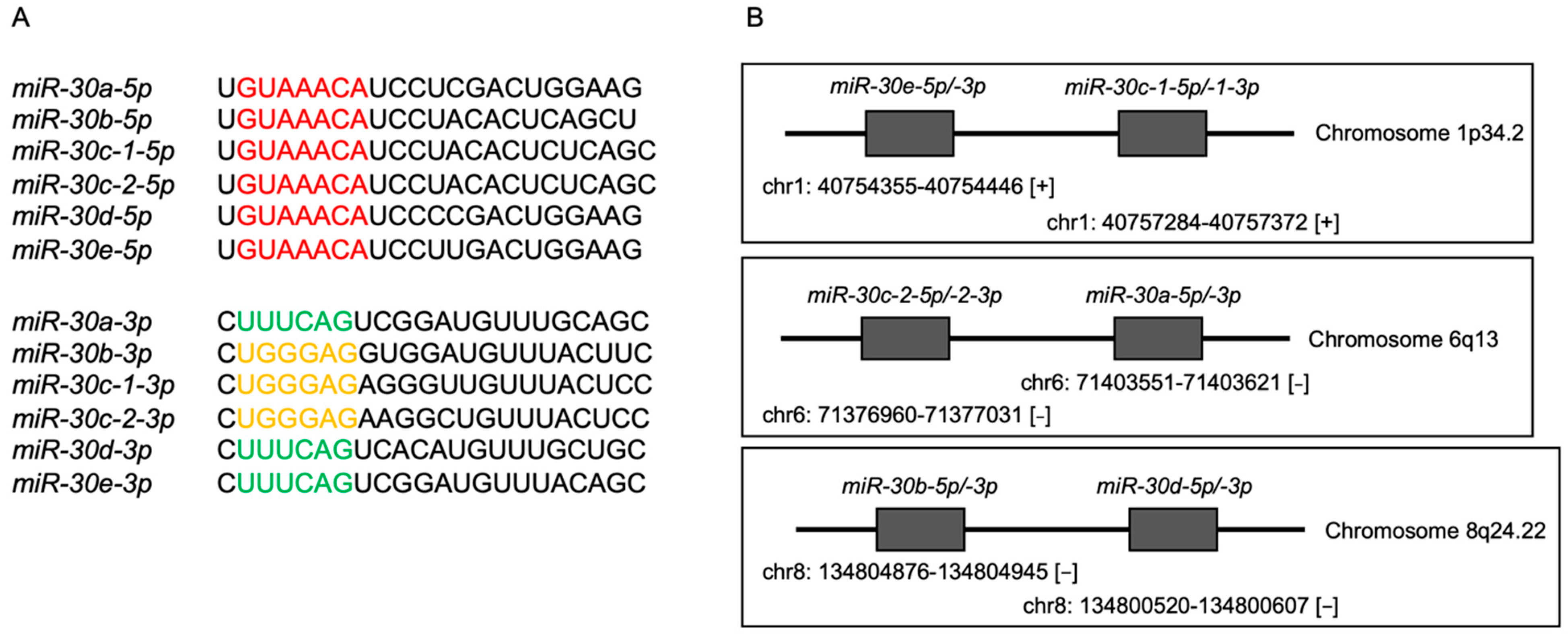
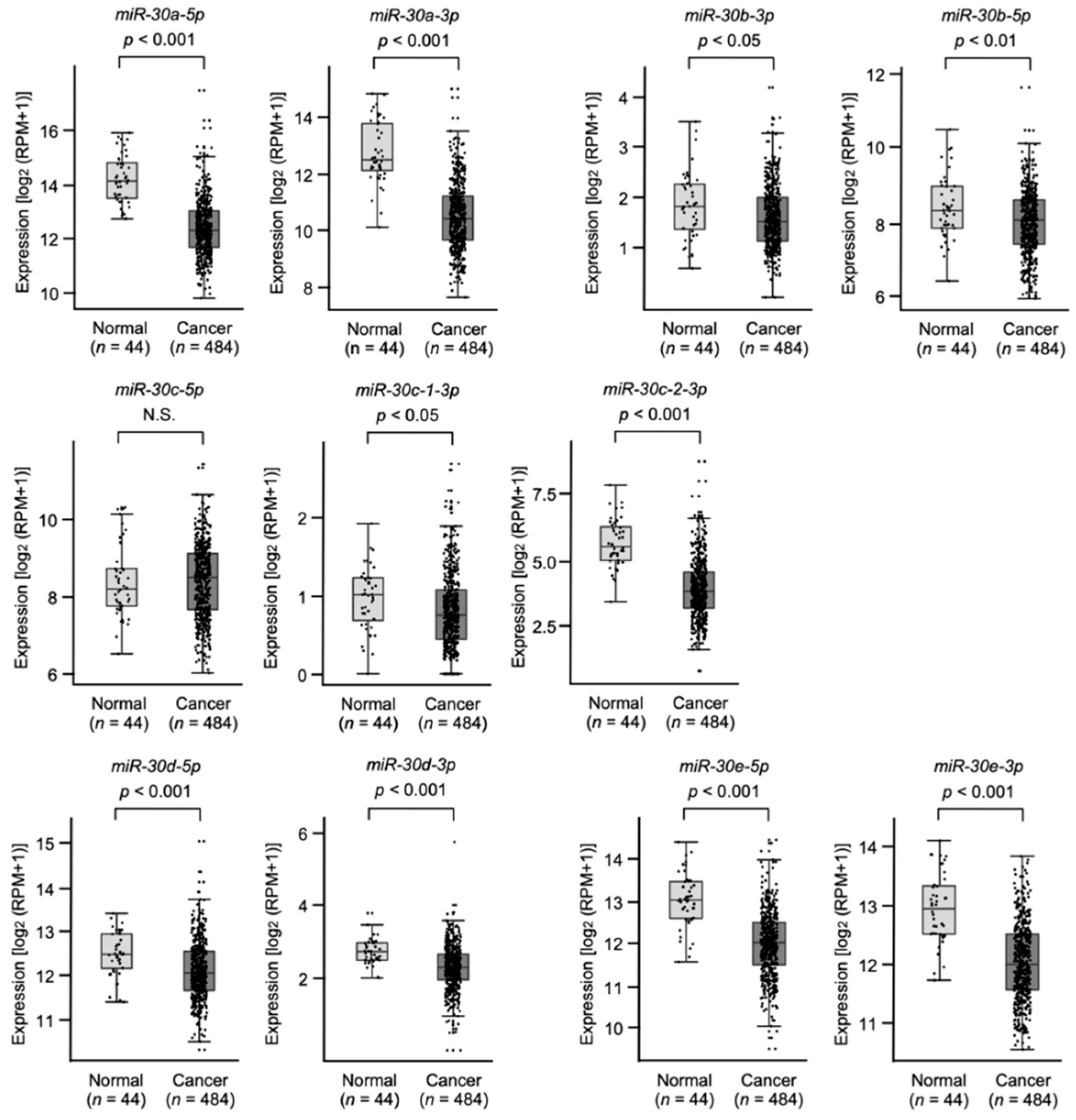
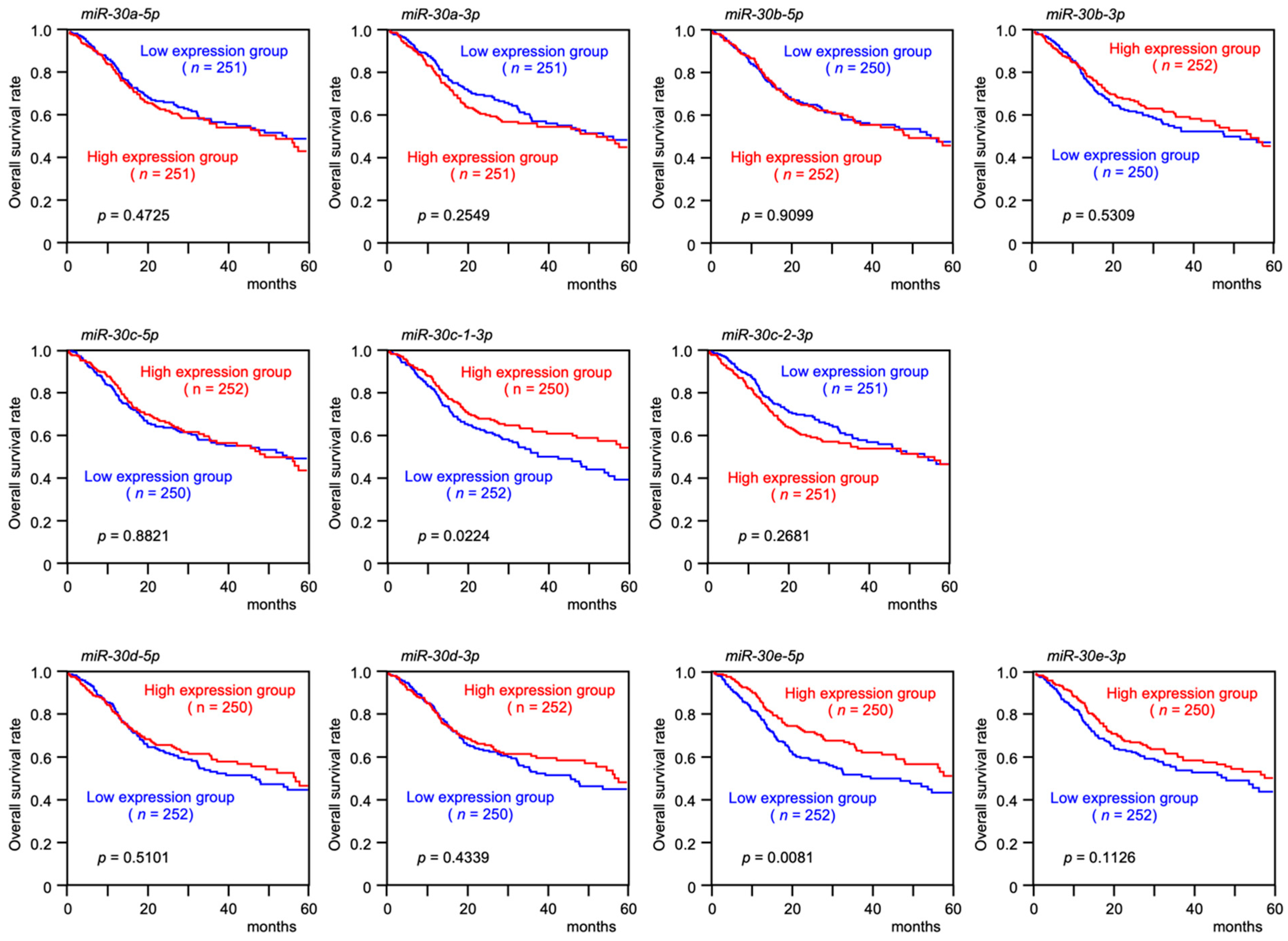
3.1. Expression Levels and the Clinical Significance of the miR-30 Family in HNSCC Clinical Specimens Assessed by TCGA Analysis
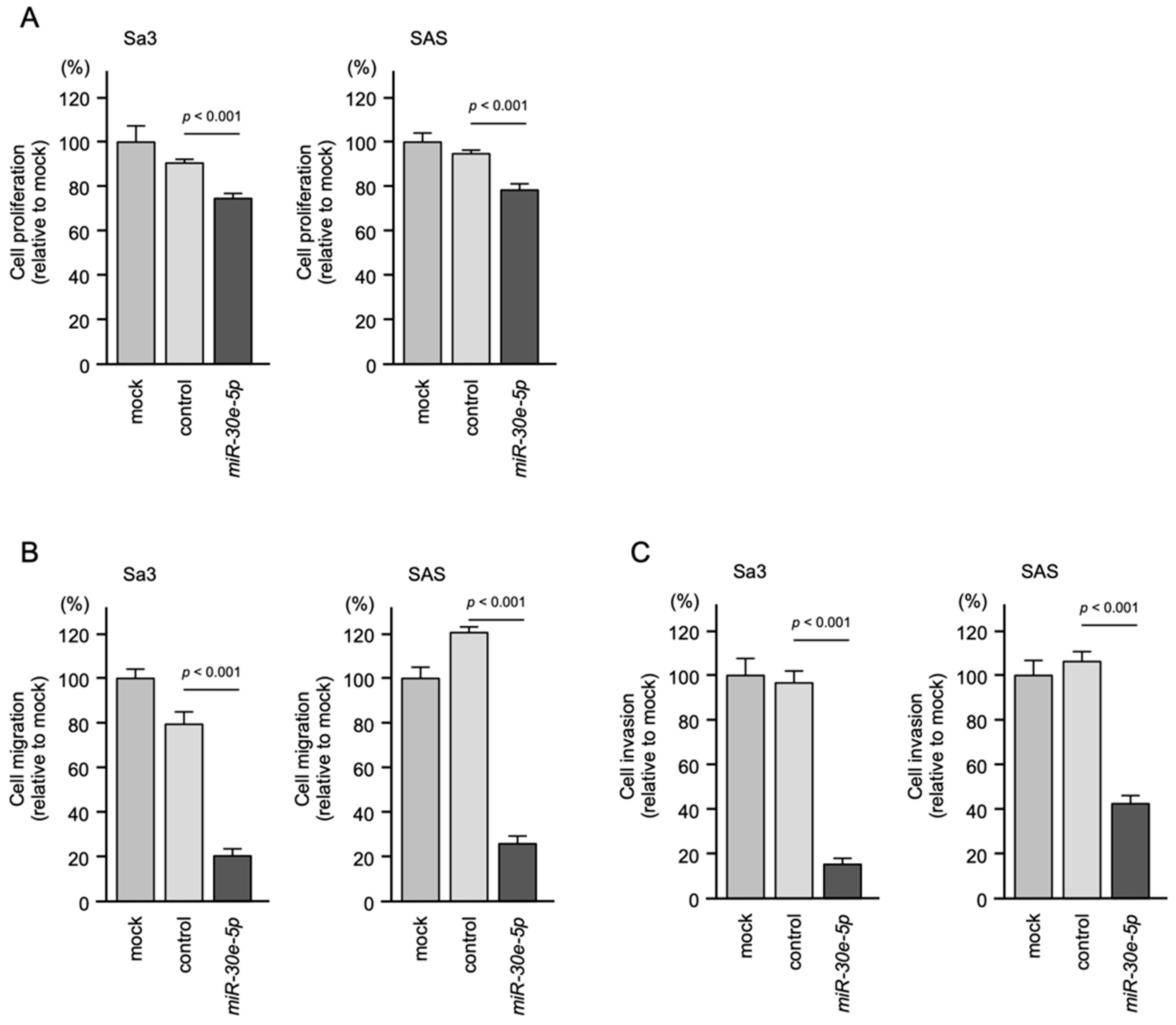
3.2. Effect of Transient Transfection of miR-30e-5p on HNSCC Cell Proliferation, Migration and Invasion
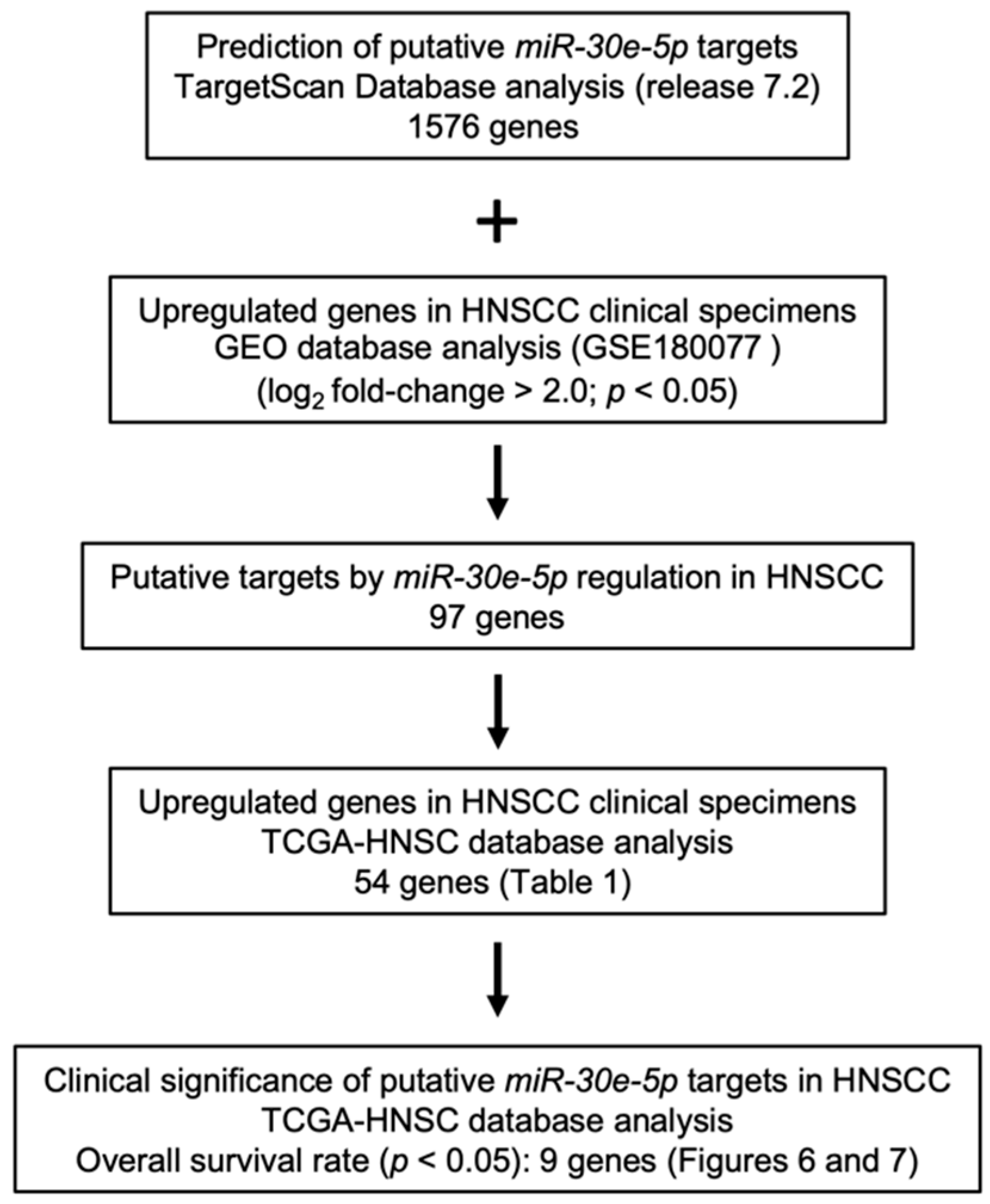
3.3. Screening for Oncogenic Targets of miR-30e-5p in HNSCC
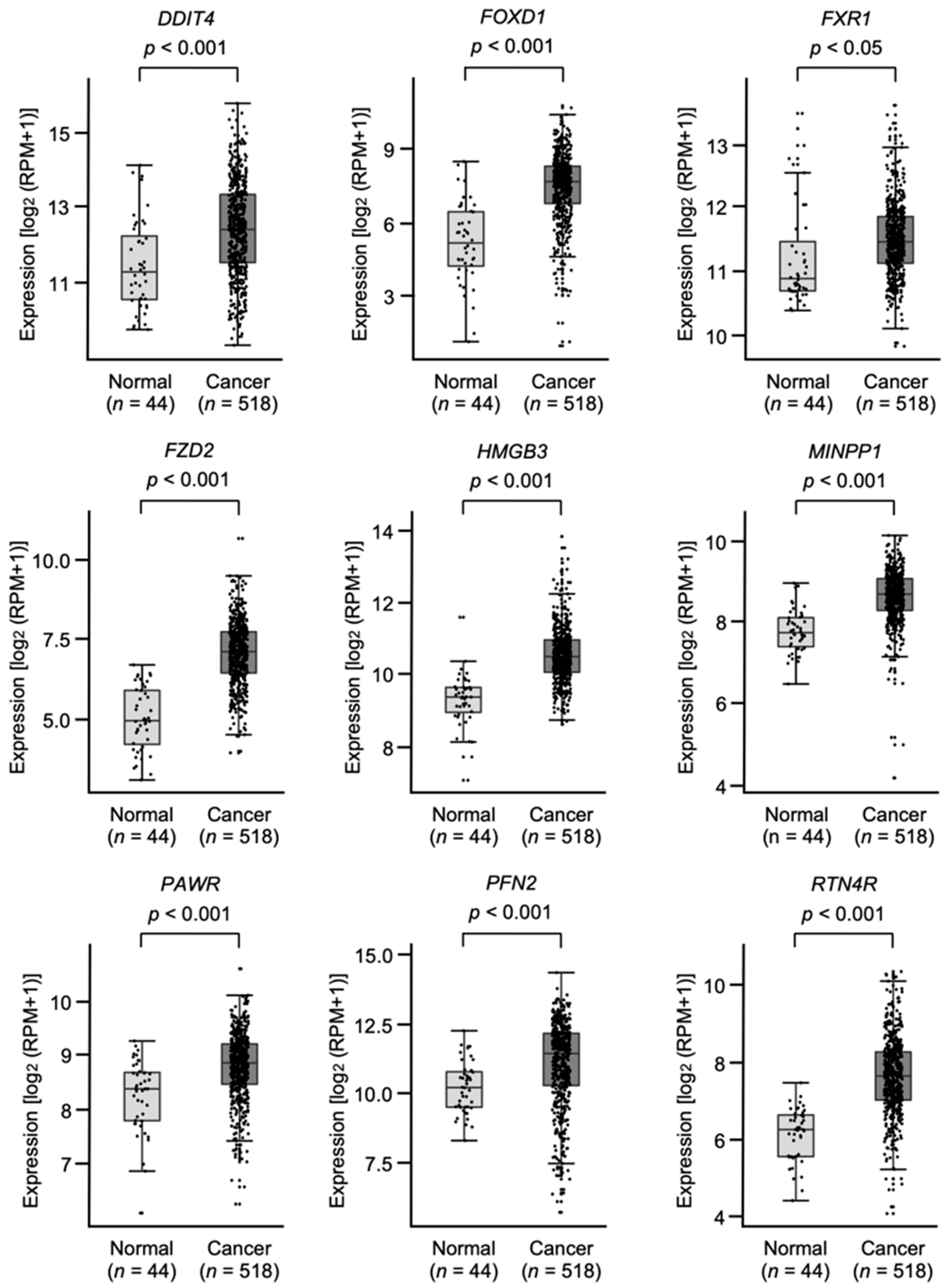
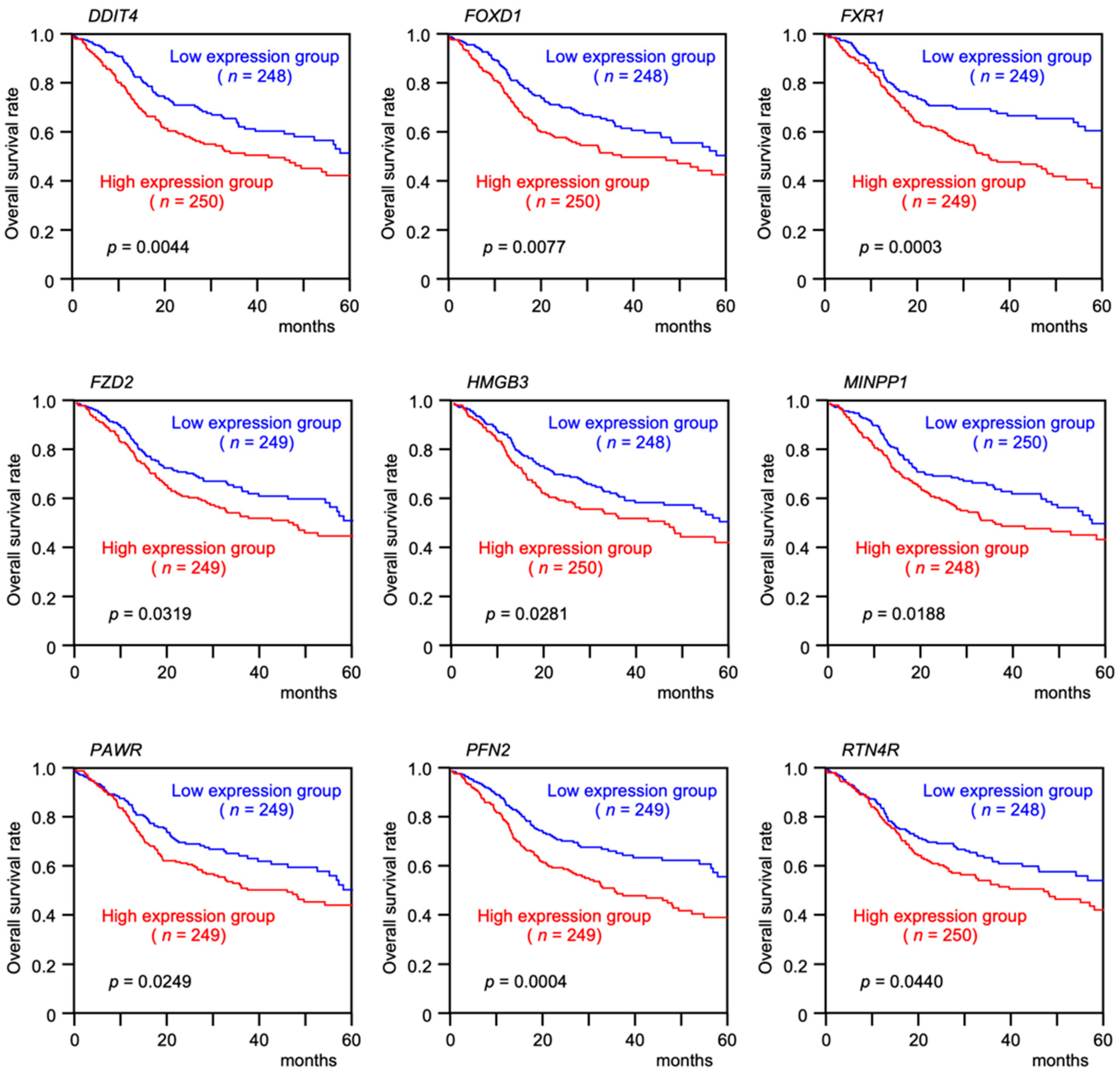
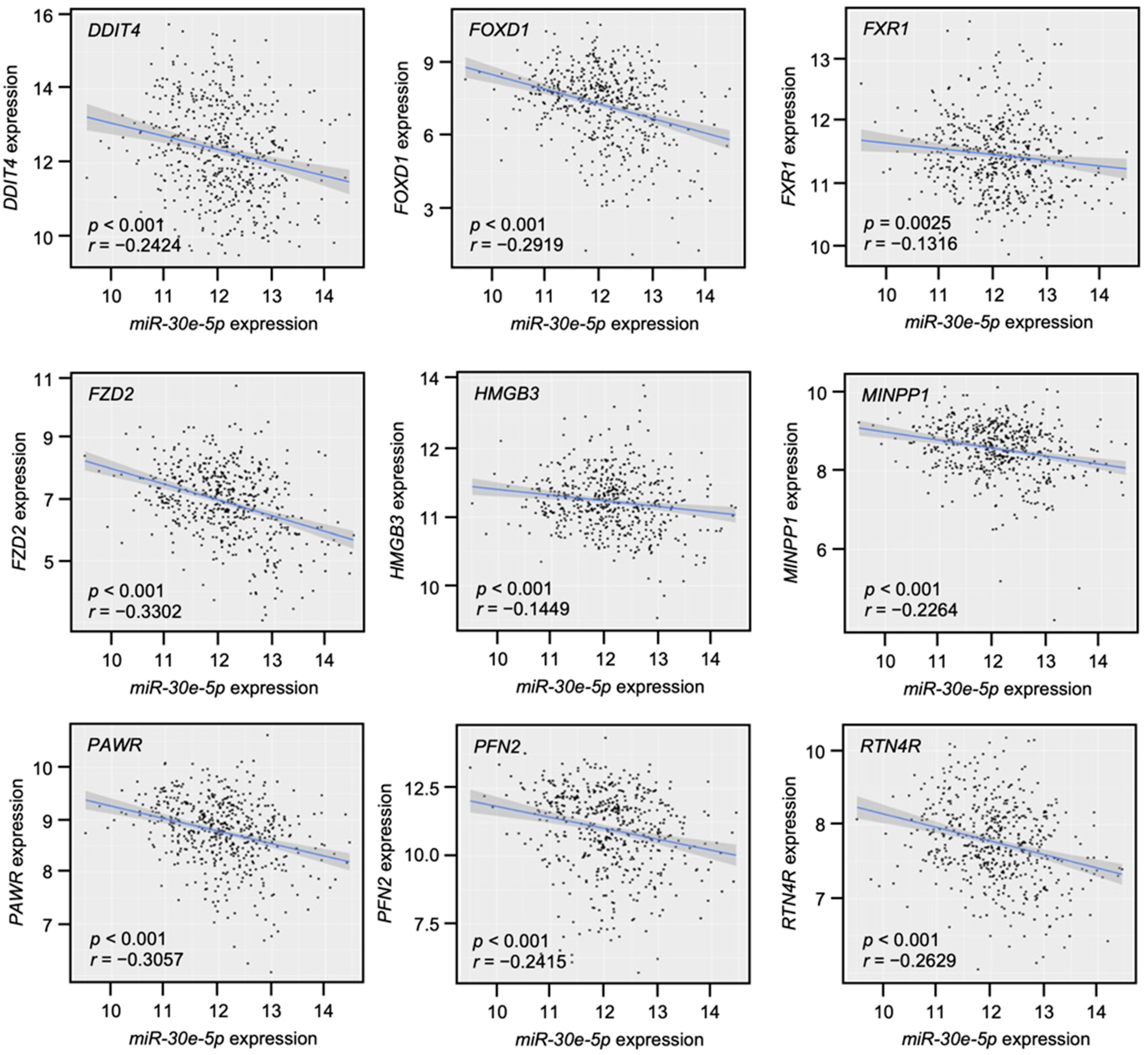
3.4. Clinical Significance of miR-30e-5p Targets in Patients with HNSCC Determined by TCGA Analysis
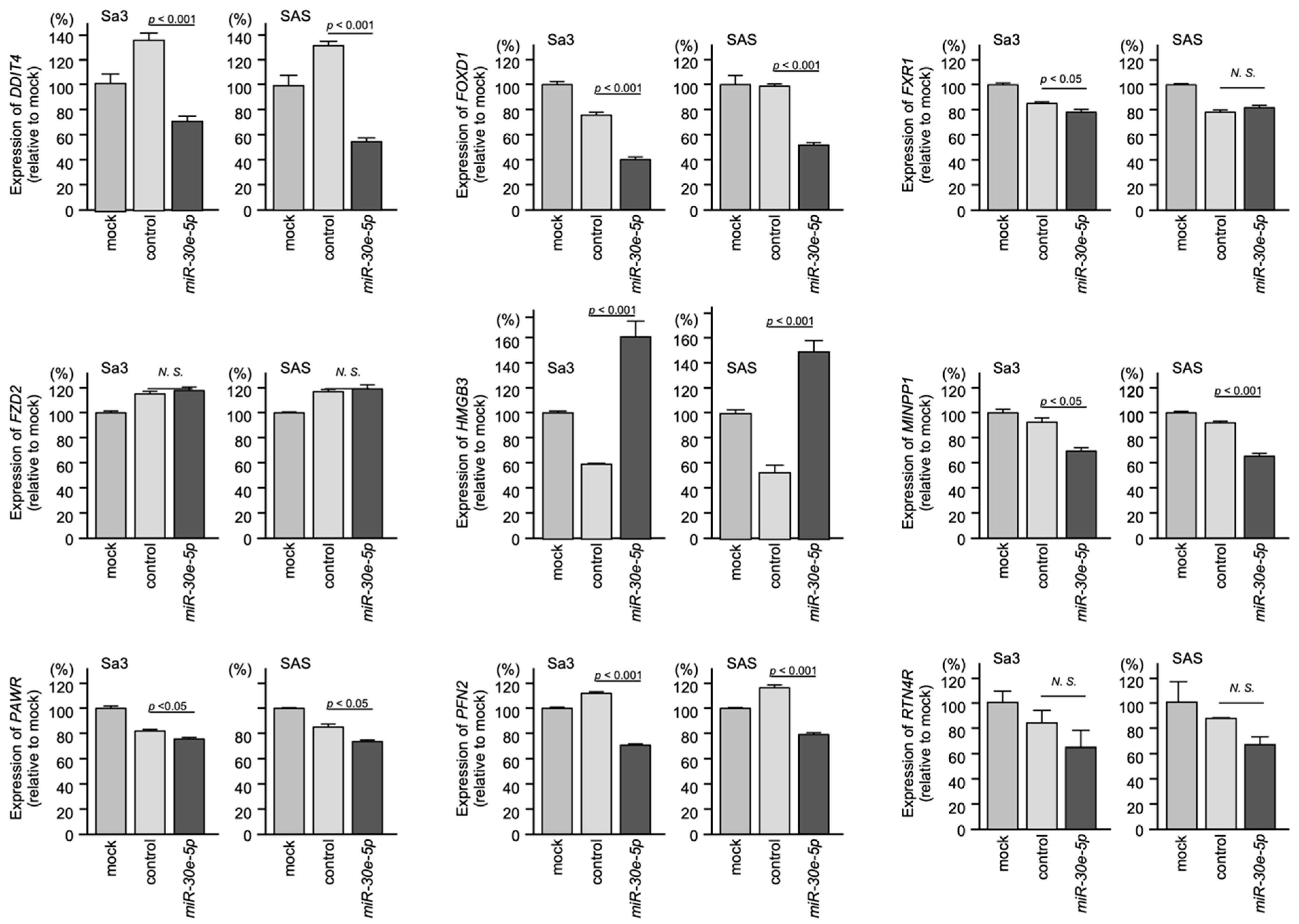
3.5. Regulated Expression of the Nine Identified Genes by miR-30e-5p in HNSCC Cells
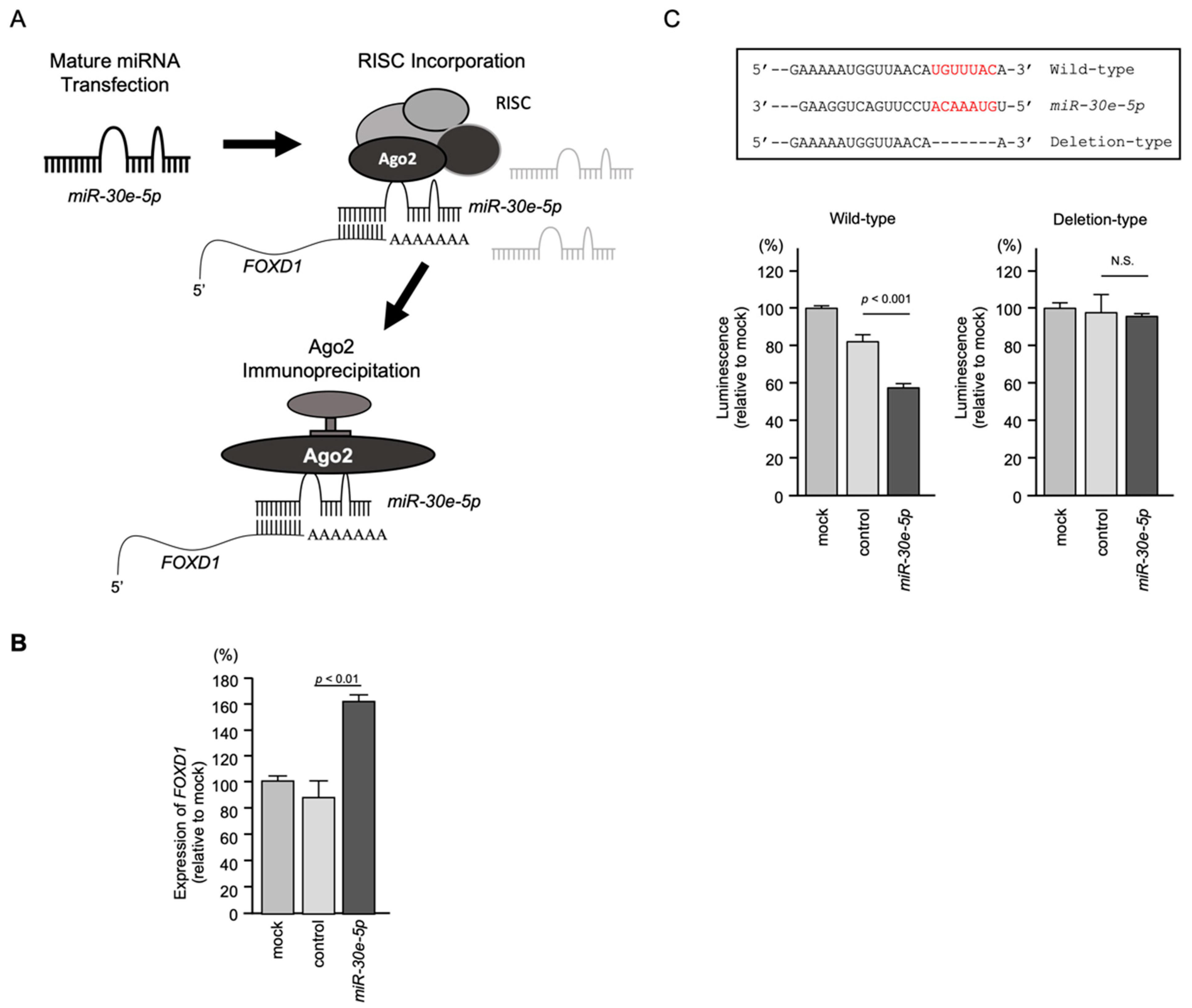
3.6. Incorporation of FOXD1 mRNA into the RNA-Induced Silencing Complex (RISC) and Direct Control of FOXD1 Expression by miR-30e-5p in HNSCC Cells
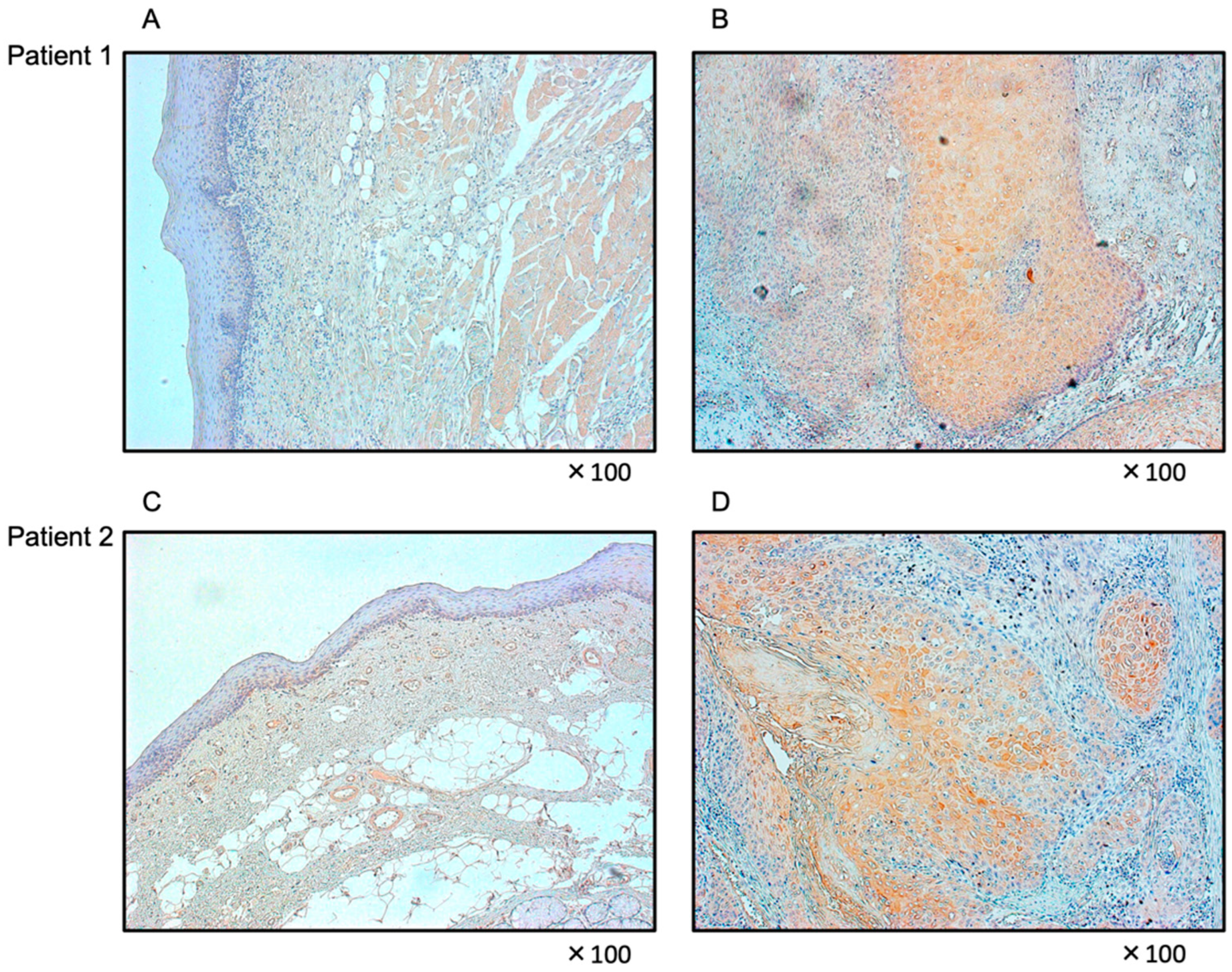
3.7. Expression of FOXD1 in HNSCC Clinical Specimens
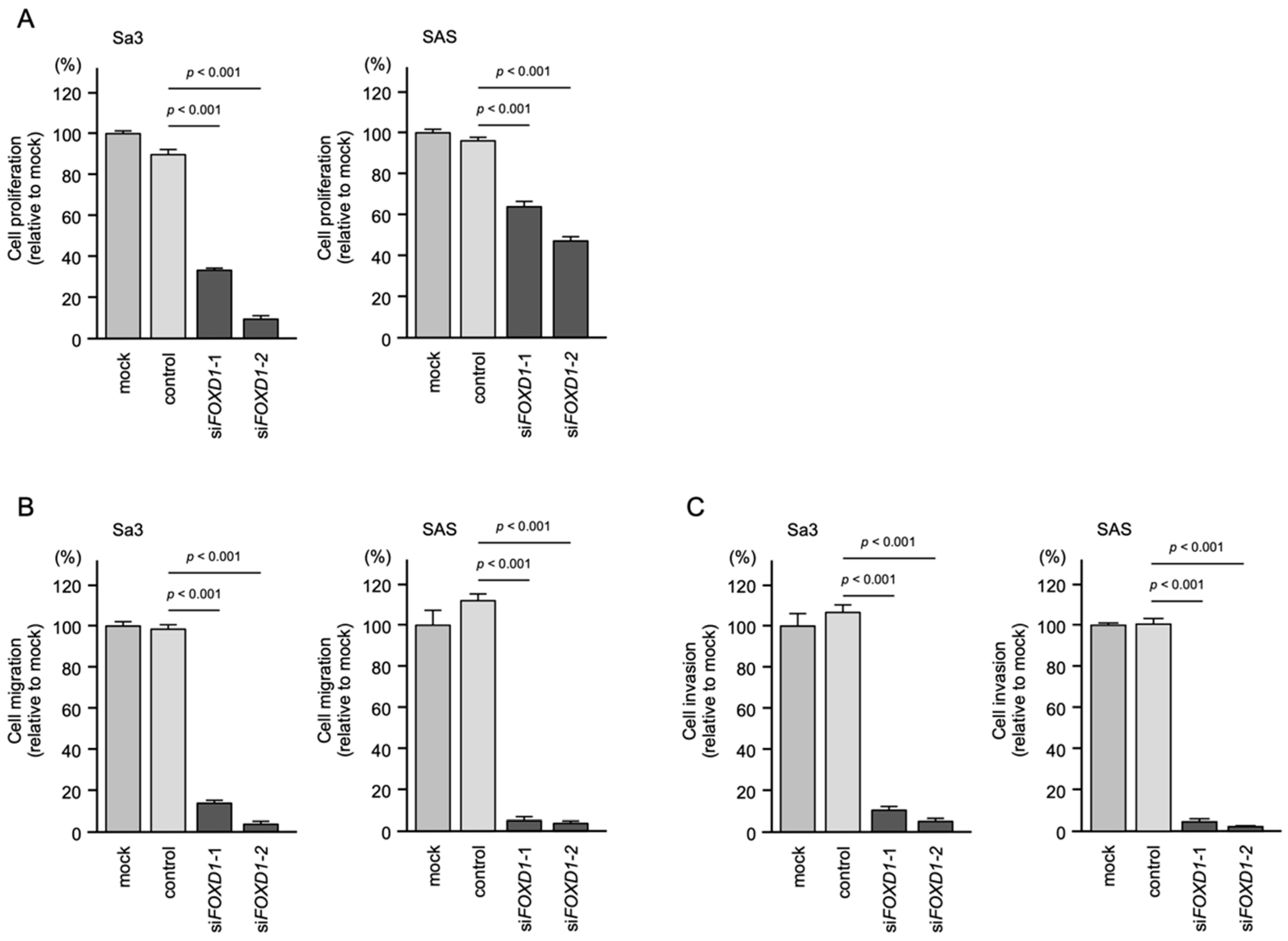
3.8. Effects of FOXD1 Knockdown on the Proliferation, Migration, and Invasion of HNSCC Cells
3.9. FOXD1-Mediated Molecular Pathways in HNSCC Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [PubMed]
- Johnson, D.E.; Burtness, B.; Leemans, C.R.; Lui, V.W.Y.; Bauman, J.E.; Grandis, J.R. Head and neck squamous cell carcinoma. Nat. Rev. Dis. Primers 2020, 6, 92. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.E.W.; Bell, R.B.; Bifulco, C.B.; Burtness, B.; Gillison, M.L.; Harrington, K.J.; Le, Q.T.; Lee, N.Y.; Leidner, R.; Lewis, R.L.; et al. The Society for Immunotherapy of Cancer consensus statement on immunotherapy for the treatment of squamous cell carcinoma of the head and neck (HNSCC). J. Immunother. Cancer 2019, 7, 184. [Google Scholar] [CrossRef] [PubMed]
- Chow, L.Q.M. Head and Neck Cancer. N. Engl. J. Med. 2020, 382, 60–72. [Google Scholar] [CrossRef]
- Bonner, J.A.; Harari, P.M.; Giralt, J.; Cohen, R.B.; Jones, C.U.; Sur, R.K.; Raben, D.; Baselga, J.; Spencer, S.A.; Zhu, J.; et al. Radiotherapy plus cetuximab for locoregionally advanced head and neck cancer: 5-year survival data from a phase 3 randomised trial, and relation between cetuximab-induced rash and survival. Lancet Oncol. 2010, 11, 21–28. [Google Scholar] [CrossRef]
- Betel, D.; Wilson, M.; Gabow, A.; Marks, D.S.; Sander, C. The microRNA.org resource: Targets and expression. Nucleic Acids Res. 2008, 36, D149–D153. [Google Scholar] [CrossRef]
- Anfossi, S.; Babayan, A.; Pantel, K.; Calin, G.A. Clinical utility of circulating non-coding RNAs—An update. Nat. Rev. Clin. Oncol. 2018, 15, 541–563. [Google Scholar] [CrossRef]
- Krek, A.; Grun, D.; Poy, M.N.; Wolf, R.; Rosenberg, L.; Epstein, E.J.; MacMenamin, P.; da Piedade, I.; Gunsalus, K.C.; Stoffel, M.; et al. Combinatorial microRNA target predictions. Nat. Genet. 2005, 37, 495–500. [Google Scholar] [CrossRef]
- Gebert, L.F.R.; MacRae, I.J. Regulation of microRNA function in animals. Nat. Rev. Mol. Cell Biol. 2019, 20, 21–37. [Google Scholar] [CrossRef]
- Rupaimoole, R.; Slack, F.J. MicroRNA therapeutics: Towards a new era for the management of cancer and other diseases. Nat. Rev. Drug Discov. 2017, 16, 203–222. [Google Scholar] [CrossRef]
- Goodall, G.J.; Wickramasinghe, V.O. RNA in cancer. Nat. Rev. Cancer 2021, 21, 22–36. [Google Scholar] [CrossRef]
- Mitra, R.; Adams, C.M.; Jiang, W.; Greenawalt, E.; Eischen, C.M. Pan-cancer analysis reveals cooperativity of both strands of microRNA that regulate tumorigenesis and patient survival. Nat. Commun. 2020, 11, 968. [Google Scholar] [CrossRef]
- Koshizuka, K.; Nohata, N.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Fukumoto, I.; Katada, K.; Okamoto, Y.; Seki, N. Deep sequencing-based microRNA expression signatures in head and neck squamous cell carcinoma: Dual strands of pre-miR-150 as antitumor miRNAs. Oncotarget 2017, 8, 30288–30304. [Google Scholar] [CrossRef]
- Yonemori, K.; Seki, N.; Idichi, T.; Kurahara, H.; Osako, Y.; Koshizuka, K.; Arai, T.; Okato, A.; Kita, Y.; Arigami, T.; et al. The microRNA expression signature of pancreatic ductal adenocarcinoma by RNA sequencing: Anti-tumour functions of the microRNA-216 cluster. Oncotarget 2017, 8, 70097–70115. [Google Scholar] [CrossRef]
- Yang, S.J.; Yang, S.Y.; Wang, D.D.; Chen, X.; Shen, H.Y.; Zhang, X.H.; Zhong, S.L.; Tang, J.H.; Zhao, J.H. The miR-30 family: Versatile players in breast cancer. Tumor Biol. 2017, 39, 1010428317692204. [Google Scholar] [CrossRef]
- Saleh, A.D.; Cheng, H.; Martin, S.E.; Si, H.; Ormanoglu, P.; Carlson, S.; Clavijo, P.E.; Yang, X.; Das, R.; Cornelius, S.; et al. Integrated Genomic and Functional microRNA Analysis Identifies miR-30-5p as a Tumor Suppressor and Potential Therapeutic Nanomedicine in Head and Neck Cancer. Clin. Cancer Res. 2019, 25, 2860–2873. [Google Scholar] [CrossRef]
- Mao, L.; Liu, S.; Hu, L.; Jia, L.; Wang, H.; Guo, M.; Chen, C.; Liu, Y.; Xu, L. miR-30 Family: A Promising Regulator in Development and Disease. BioMed Res. Int. 2018, 2018, 9623412. [Google Scholar] [CrossRef]
- Kozomara, A.; Birgaoanu, M.; Griffiths-Jones, S. miRBase: From microRNA sequences to function. Nucleic Acids Res. 2019, 47, D155–D162. [Google Scholar] [CrossRef]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef]
- Anaya, J. Linking TCGA survival data to mRNAs, miRNAs, and lncRNAs. PeerJ Comput. Sci. 2016, 2, e67. [Google Scholar] [CrossRef]
- Agarwal, V.; Bell, G.W.; Nam, J.W.; Bartel, D.P. Predicting effective microRNA target sites in mammalian mRNAs. eLife 2015, 4, e05005. [Google Scholar] [CrossRef]
- Koshizuka, K.; Hanazawa, T.; Kikkawa, N.; Arai, T.; Okato, A.; Kurozumi, A.; Kato, M.; Katada, K.; Okamoto, Y.; Seki, N. Regulation of ITGA3 by the anti-tumor miR-199 family inhibits cancer cell migration and invasion in head and neck cancer. Cancer Sci. 2017, 108, 1681–1692. [Google Scholar] [CrossRef]
- Koma, A.; Asai, S.; Minemura, C.; Oshima, S.; Kinoshita, T.; Kikkawa, N.; Koshizuka, K.; Moriya, S.; Kasamatsu, A.; Hanazawa, T.; et al. Impact of Oncogenic Targets by Tumor-Suppressive miR-139-5p and miR-139-3p Regulation in Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2021, 22, 9947. [Google Scholar] [CrossRef]
- Asai, S.; Koma, A.; Nohata, N.; Kinoshita, T.; Kikkawa, N.; Kato, M.; Minemura, C.; Uzawa, K.; Hanazawa, T.; Seki, N. Impact of miR-1/miR-133 Clustered miRNAs: PFN2 Facilitates Malignant Phenotypes in Head and Neck Squamous Cell Carcinoma. Biomedicines 2022, 10, 663. [Google Scholar] [CrossRef]
- Minemura, C.; Asai, S.; Koma, A.; Kase-Kato, I.; Tanaka, N.; Kikkawa, N.; Kasamatsu, A.; Yokoe, H.; Hanazawa, T.; Uzawa, K.; et al. Identification of Tumor-Suppressive miR-30e-3p Targets: Involvement of SERPINE1 in the Molecular Pathogenesis of Head and Neck Squamous Cell Carcinoma. Int. J. Mol. Sci. 2022, 23, 3808. [Google Scholar] [CrossRef]
- Mootha, V.K.; Lindgren, C.M.; Eriksson, K.F.; Subramanian, A.; Sihag, S.; Lehar, J.; Puigserver, P.; Carlsson, E.; Ridderstråle, M.; Laurila, E.; et al. PGC-1alpha-responsive genes involved in oxidative phosphorylation are coordinately downregulated in human diabetes. Nat. Genet. 2003, 34, 267–273. [Google Scholar] [CrossRef]
- Subramanian, A.; Tamayo, P.; Mootha, V.K.; Mukherjee, S.; Ebert, B.L.; Gillette, M.A.; Paulovich, A.; Pomeroy, S.L.; Golub, T.R.; Lander, E.S.; et al. Gene set enrichment analysis: A knowledge-based approach for interpreting genome-wide expression profiles. Proc. Natl. Acad. Sci. USA 2005, 102, 15545–15550. [Google Scholar] [CrossRef]
- Liberzon, A.; Subramanian, A.; Pinchback, R.; Thorvaldsdóttir, H.; Tamayo, P.; Mesirov, J.P. Molecular signatures database (MSigDB) 3.0. Bioinformatics 2011, 27, 1739–1740. [Google Scholar] [CrossRef]
- Yamada, Y.; Sugawara, S.; Arai, T.; Kojima, S.; Kato, M.; Okato, A.; Yamazaki, K.; Naya, Y.; Ichikawa, T.; Seki, N. Molecular pathogenesis of renal cell carcinoma: Impact of the anti-tumor miR-29 family on gene regulation. Int. J. Urol. 2018, 25, 953–965. [Google Scholar] [CrossRef]
- Kanthaje, S.; Baikunje, N.; Kandal, I.; Ratnacaram, C.K. Repertoires of MicroRNA-30 family as gate-keepers in lung cancer. Front. Biosci. 2021, 13, 141–156. [Google Scholar] [CrossRef]
- Xu, G.; Cai, J.; Wang, L.; Jiang, L.; Huang, J.; Hu, R.; Ding, F. MicroRNA-30e-5p suppresses non-small cell lung cancer tumorigenesis by regulating USP22-mediated Sirt1/JAK/STAT3 signaling. Exp. Cell Res. 2018, 362, 268–278. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.X.; Zhang, H.; Li, X.H.; Liu, Y.H. MiR-30e-5p inhibits proliferation and metastasis of nasopharyngeal carcinoma cells by target-ing USP22. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 6342–6349. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Qin, H.; Jiang, B.; Chen, W.; Cao, W.; Zhao, X.; Yuan, H.; Qi, W.; Zhuo, D.; Guo, H. miR-30e-5p suppresses cell proliferation and migration in bladder cancer through regulating metadherin. J. Cell. Biochem. 2019, 120, 15924–15932. [Google Scholar] [CrossRef]
- Zhang, S.; Li, G.; Liu, C.; Lu, S.; Jing, Q.; Chen, X.; Zheng, H.; Ma, H.; Zhang, D.; Ren, S.; et al. miR-30e-5p represses angiogenesis and metastasis by directly targeting AEG-1 in squamous cell carcinoma of the head and neck. Cancer Sci. 2020, 111, 356–368. [Google Scholar] [CrossRef]
- Laudato, S.; Patil, N.; Abba, M.L.; Leupold, J.H.; Benner, A.; Gaiser, T.; Marx, A.; Allgayer, H. P53-induced miR-30e-5p inhibits colorectal cancer invasion and metastasis by targeting ITGA6 and ITGB1. Int. J. Cancer 2017, 141, 1879–1890. [Google Scholar] [CrossRef]
- Majumder, M.; Johnson, R.H.; Palanisamy, V. Fragile X-related protein family: A double-edged sword in neurodevelopmental disorders and cancer. Crit. Rev. Biochem. Mol. Biol. 2020, 55, 409–424. [Google Scholar] [CrossRef]
- Qian, J.; Hassanein, M.; Hoeksema, M.D.; Harris, B.K.; Zou, Y.; Chen, H.; Lu, P.; Eisenberg, R.; Wang, J.; Espinosa, A.; et al. The RNA binding protein FXR1 is a new driver in the 3q26-29 amplicon and predicts poor prognosis in human cancers. Proc. Natl. Acad. Sci. USA 2015, 112, 3469–3474. [Google Scholar] [CrossRef]
- Majumder, M.; Palanisamy, V. RNA binding protein FXR1-miR301a-3p axis contributes to p21WAF1 degradation in oral cancer. PLoS Genet. 2020, 16, e1008580. [Google Scholar] [CrossRef]
- Qie, S.; Majumder, M.; Mackiewicz, K.; Howley, B.V.; Peterson, Y.K.; Howe, P.H.; Palanisamy, V.; Diehl, J.A. Fbxo4-mediated degradation of Fxr1 suppresses tumorigenesis in head and neck squamous cell carcinoma. Nat. Commun. 2017, 8, 1534. [Google Scholar] [CrossRef]
- Pinto-Costa, R.; Sousa, M.M. Profilin as a dual regulator of actin and microtubule dynamics. Cytoskeleton 2020, 77, 76–83. [Google Scholar] [CrossRef]
- Zhou, K.; Chen, J.; Wu, J.; Xu, Y.; Wu, Q.; Yue, J.; Song, Y.; Li, S.; Zhou, P.; Tu, W.; et al. Profilin 2 Promotes Proliferation and Metastasis of Head and Neck Cancer Cells by Regulating PI3K/AKT/β-Catenin Signaling Pathway. Oncol. Res. 2019, 27, 1079–1088. [Google Scholar] [CrossRef]
- Cao, Q.; Liu, Y.; Wu, Y.; Hu, C.; Sun, L.; Wang, J.; Li, C.; Guo, M.; Liu, X.; Lv, J.; et al. Profilin 2 promotes growth, metastasis, and angiogenesis of small cell lung cancer through cancer-derived exosomes. Aging 2020, 12, 25981–25999. [Google Scholar] [CrossRef]
- Yan, J.; Ma, C.; Gao, Y. MicroRNA-30a-5p suppresses epithelial-mesenchymal transition by targeting profilin-2 in high invasive non-small cell lung cancer cell lines. Oncol. Rep. 2017, 37, 3146–3154. [Google Scholar] [CrossRef][Green Version]
- Katoh, M.; Katoh, M. Human FOX gene family (Review). Int. J. Oncol. 2004, 25, 1495–1500. [Google Scholar] [CrossRef]
- Quintero-Ronderos, P.; Laissue, P. The multisystemic functions of FOXD1 in development and disease. J. Mol. Med. 2018, 96, 725–739. [Google Scholar] [CrossRef]
- Lin, C.H.; Lee, H.H.; Chang, W.M.; Lee, F.P.; Chen, L.C.; Lu, L.S.; Lin, Y.F. FOXD1 Repression Potentiates Radiation Effectiveness by Downregulating G3BP2 Expression and Promoting the Activation of TXNIP-Related Pathways in Oral Cancer. Cancers 2020, 12, 2690. [Google Scholar] [CrossRef]
- Sun, Q.; Novak, D.; Hüser, L.; Poelchen, J.; Wu, H.; Granados, K.; Federico, A.; Liu, K.; Steinfass, T.; Vierthaler, M.; et al. FOXD1 promotes dedifferentiation and targeted therapy resistance in melanoma by regulating the expression of connective tissue growth factor. Int. J. Cancer 2021, 149, 657–674. [Google Scholar] [CrossRef]
- Wu, Q.; Ma, J.; Wei, J.; Meng, W.; Wang, Y.; Shi, M. FOXD1-AS1 regulates FOXD1 translation and promotes gastric cancer progression and chemoresistance by activating the PI3K/AKT/mTOR pathway. Mol. Oncol. 2021, 15, 299–316. [Google Scholar] [CrossRef]
- Chen, C.; Tang, J.; Xu, S.; Zhang, W.; Jiang, H. miR-30a-5p Inhibits Proliferation and Migration of Lung Squamous Cell Carcinoma Cells by Targeting FOXD1. BioMed Res. Int. 2020, 2020, 2547902. [Google Scholar] [CrossRef]
- Zhou, L.; Jia, S.; Ding, G.; Zhang, M.; Yu, W.; Wu, Z.; Cao, L. Down-regulation of miR-30a-5p is Associated with Poor Prognosis and Promotes Chemoresistance of Gemcitabine in Pancreatic Ductal Adenocarcinoma. J. Cancer 2019, 10, 5031–5040. [Google Scholar] [CrossRef] [PubMed]
- Tao, J.; Cong, H.; Wang, H.; Zhang, D.; Liu, C.; Chu, H.; Qing, Q.; Wang, K. MiR-30a-5p inhibits osteosarcoma cell proliferation and migration by targeting FOXD1. Biochem. Biophys. Res. Commun. 2018, 503, 1092–1097. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qiu, C.; Lu, N.; Liu, Z.; Jin, C.; Sun, C.; Bu, H.; Yu, H.; Dongol, S.; Kong, B. FOXD1 is targeted by miR-30a-5p and miR-200a-5p and suppresses the proliferation of human ovarian carcinoma cells by promoting p21 expression in a p53-independent manner. Int. J. Oncol. 2018, 52, 2130–2142. [Google Scholar] [CrossRef] [PubMed]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Entrez Gene ID | Gene Symbol | Gene Name | Total Binding Sites | GEO 1 p Value | GEO log2(FC 2) | 5y OS 3 p Value |
|---|---|---|---|---|---|---|
| 8087 | FXR1 | Fragile X mental retardation, autosomal homolog 1 | 1 | 0.014 | 3.06 | <0.001 |
| 5217 | PFN2 | Profilin 2 | 1 | 0.004 | 2.67 | <0.001 |
| 54541 | DDIT4 | DNA-damage-inducible transcript 4 | 1 | 0.017 | 2.70 | 0.004 |
| 2297 | FOXD1 | Forkhead box D1 | 1 | 0.008 | 3.79 | 0.008 |
| 9562 | MINPP1 | Multiple inositol-polyphosphate phosphatase 1 | 1 | 0.013 | 2.02 | 0.019 |
| 5074 | PAWR | PRKC, apoptosis, WT1, regulator | 2 | 0.003 | 2.04 | 0.025 |
| 3149 | HMGB3 | High mobility group box 3 | 1 | 0.004 | 2.17 | 0.028 |
| 2535 | FZD2 | Frizzled class receptor 2 | 1 | 0.006 | 2.36 | 0.032 |
| 65078 | RTN4R | Reticulon 4 receptor | 1 | 0.008 | 2.20 | 0.044 |
| 115908 | CTHRC1 | Collagen triple helix repeat containing 1 | 1 | 0.010 | 2.96 | 0.059 |
| 3218 | HOXB8 | Homeobox B8 | 1 | 0.046 | 3.29 | 0.077 |
| 9143 | SYNGR3 | Synaptogyrin 3 | 1 | 0.042 | 2.12 | 0.077 |
| 1012 | CDH13 | Cadherin 13 | 1 | 0.013 | 2.13 | 0.091 |
| 6683 | SPAST | Spastin | 2 | 0.012 | 2.10 | 0.092 |
| 79718 | TBL1XR1 | Transducin (β)-like 1 X-linked receptor 1 | 2 | 0.002 | 2.63 | 0.104 |
| 114088 | TRIM9 | Tripartite motif containing 9 | 1 | 0.005 | 4.27 | 0.113 |
| 84733 | CBX2 | Chromobox homolog 2 | 1 | 0.005 | 3.00 | 0.133 |
| 27 | ABL2 | ABL proto-oncogene 2, non-receptor tyrosine kinase | 1 | 0.008 | 2.19 | 0.155 |
| 23657 | SLC7A11 | solute carrier family 7 (Anionic amino acid transporter light chain, xc- system), member 11 | 1 | 0.007 | 4.00 | 0.160 |
| 154214 | RNF217 | Ring finger protein 217 | 1 | 0.016 | 2.36 | 0.186 |
| 79712 | GTDC1 | Glycosyltransferase-like domain containing 1 | 1 | 0.004 | 4.31 | 0.193 |
| 26059 | ERC2 | ELKS/RAB6-interacting/CAST family member 2 | 1 | 0.027 | 3.54 | 0.210 |
| 3237 | HOXD11 | Homeobox D11 | 1 | 0.033 | 4.24 | 0.214 |
| 89796 | NAV1 | Neuron navigator 1 | 1 | 0.007 | 2.88 | 0.234 |
| 6659 | SOX4 | SRY (sex determining region Y)-box 4 | 1 | 0.005 | 2.30 | 0.258 |
| 54434 | SSH1 | Slingshot protein phosphatase 1 | 1 | 0.016 | 2.05 | 0.280 |
| 2048 | EPHB2 | EPH receptor B2 | 1 | 0.013 | 2.53 | 0.303 |
| 9258 | MFHAS1 | Malignant fibrous histiocytoma amplified sequence 1 | 1 | 0.005 | 2.27 | 0.311 |
| 54566 | EPB41L4B | Erythrocyte membrane protein band 4.1 like 4B | 1 | 0.004 | 3.06 | 0.413 |
| 8448 | DOC2A | Double C2-like domains, α | 2 | 0.021 | 3.00 | 0.485 |
| 28982 | FLVCR1 | Feline leukemia virus subgroup C cellular receptor 1 | 1 | 0.005 | 2.43 | 0.492 |
| 55785 | FGD6 | FYVE, RhoGEF and PH domain containing 6 | 1 | 0.014 | 2.21 | 0.530 |
| 490 | ATP2B1 | ATPase, Ca++ transporting, plasma membrane 1 | 1 | 0.007 | 2.93 | 0.556 |
| 4644 | MYO5A | Myosin VA (heavy chain 12, myoxin) | 1 | 0.003 | 2.06 | 0.605 |
| 4015 | LOX | Lysyl oxidase | 1 | 0.006 | 4.46 | 0.613 |
| 50805 | IRX4 | Iroquois homeobox 4 | 1 | 0.039 | 2.67 | 0.652 |
| 23432 | GPR161 | G protein-coupled receptor 161 | 1 | 0.008 | 2.50 | 0.662 |
| 2729 | GCLC | Glutamate-cysteine ligase, catalytic subunit | 1 | 0.004 | 3.14 | 0.710 |
| 8038 | ADAM12 | ADAM metallopeptidase domain 12 | 2 | 0.003 | 4.29 | 0.721 |
| 3631 | INPP4A | Inositol polyphosphate-4-phosphatase, type I, 107kDa | 1 | 0.003 | 2.12 | 0.747 |
| 9832 | JAKMIP2 | Janus kinase and microtubule interacting protein 2 | 1 | 0.010 | 4.19 | 0.769 |
| 121268 | RHEBL1 | Ras homolog enriched in brain like 1 | 1 | 0.004 | 2.92 | 0.775 |
| 144455 | E2F7 | E2F transcription factor 7 | 3 | 0.021 | 2.47 | 0.786 |
| 94032 | CAMK2N2 | Calcium/calmodulin-dependent protein kinase II inhibitor 2 | 1 | 0.003 | 2.95 | 0.823 |
| 84206 | MEX3B | Mex-3 RNA binding family member B | 2 | 0.016 | 2.75 | 0.880 |
| 2887 | GRB10 | Growth factor receptor-bound protein 10 | 1 | 0.008 | 2.07 | 0.890 |
| 23333 | DPY19L1 | Dpy-19-like 1 (C. elegans) | 1 | 0.013 | 3.07 | 0.893 |
| 9435 | CHST2 | Carbohydrate (N-acetylglucosamine-6-O) sulfotransferase 2 | 1 | 0.006 | 3.30 | 0.917 |
| 54165 | DCUN1D1 | DCN1, defective in cullin neddylation 1, domain containing 1 | 2 | 0.009 | 2.50 | 0.938 |
| 54714 | CNGB3 | Cyclic nucleotide gated channel β 3 | 1 | 0.008 | 2.98 | 0.968 |
| 221002 | RASGEF1A | RasGEF domain family, member 1A | 1 | 0.005 | 2.81 | 0.968 |
| 55144 | LRRC8D | Leucine rich repeat containing 8 family, member D | 1 | 0.002 | 2.60 | 0.978 |
| 8534 | CHST1 | Carbohydrate (keratan sulfate Gal-6) sulfotransferase 1 | 1 | 0.005 | 4.41 | 0.980 |
| 8632 | DNAH17 | Dynein, axonemal, heavy chain 17 | 1 | 0.006 | 5.26 | 0.999 |
| Monovariate | Multivariate | |||||
|---|---|---|---|---|---|---|
| Variables | HR | 95% CI | p-Value | HR | 95% CI | p-Value |
| DDIT4 (High vs. Low expression) | 1.506 | 1.138–1.994 | 0.004 | 1.292 | 0.939–1.776 | 0.115 |
| FOXD1 (High vs. Low expression) | 1.526 | 1.153–2.019 | 0.003 | 1.374 | 1.002–1.890 | 0.049 |
| FXR1 (High vs. Low expression) | 1.651 | 1.242–2.193 | 0.001 | 1.303 | 0.930–1.825 | 0.124 |
| FZD2 (High vs. Low expression) | 1.337 | 1.011–1.768 | 0.042 | 1.069 | 0.776–1.473 | 0.684 |
| HMGB3 (High vs. Low expression) | 1.413 | 1.069–1.867 | 0.015 | 1.186 | 0.861–1.633 | 0.297 |
| MINPP1 (High vs. Low expression) | 1.451 | 1.096–1.921 | 0.009 | 1.308 | 0.940–1.818 | 0.111 |
| PAWR (High vs. Low expression) | 1.438 | 1.084–1.908 | 0.012 | 1.255 | 0.913–1.727 | 0.162 |
| PFN2 (High vs. Low expression) | 1.642 | 1.238–2.178 | 0.001 | 1.238 | 0.885–1.731 | 0.213 |
| RTN4R (High vs. Low expression) | 1.326 | 1.003–1.752 | 0.048 | 0.962 | 0.699–1.323 | 0.810 |
| Age (≥70 vs. <70) | 1.628 | 1.197–2.213 | 0.002 | 1.922 | 1.369–2.698 | <0.001 |
| Disease Stage (III, IV vs. I, II) | 1.746 | 1.158–2.633 | 0.008 | 1.774 | 1.159–2.716 | 0.008 |
| Pathological Grade (3, 4 vs. 1, 2) | 0.901 | 0.653–1.245 | 0.529 | - | - | - |
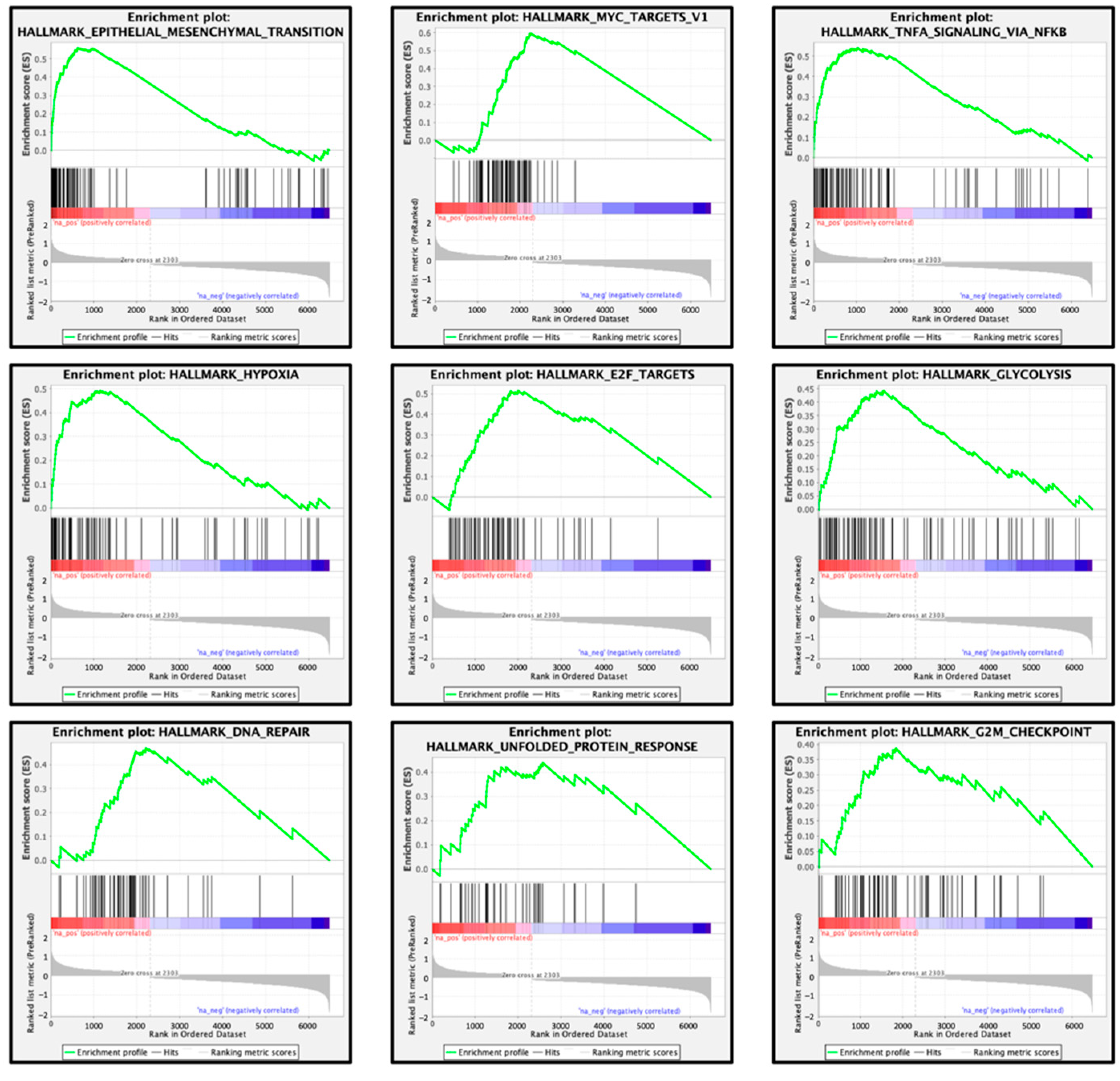
| Enriched Gene Sets in High FOXD1 Expression Group | ||
|---|---|---|
| Name | Normalized Enrichment Score | FDR q-Value |
| Epithelial mesenchymal transition | 3.674 | q < 0.001 |
| MYC targets V1 | 3.412 | q < 0.001 |
| TNFα signaling via NFκB | 3.262 | q < 0.001 |
| Hypoxia | 3.044 | q < 0.001 |
| E2F targets | 2.988 | q < 0.001 |
| Glycolysis | 2.797 | q < 0.001 |
| DNA repair | 2.602 | q < 0.001 |
| Unfolded protein response | 2.275 | 0.001 |
| G2M checkpoint | 2.248 | 0.001 |
| TGFβ signaling | 2.025 | 0.001 |
| KRAS signaling up | 1.946 | 0.005 |
| Coagulation | 1.848 | 0.010 |
| Inflammatory response | 1.753 | 0.014 |
| Apical junction | 1.739 | 0.015 |
| Oxidative phosphorylation | 1.713 | 0.015 |
| P53 pathway | 1.702 | 0.016 |
| IL6/JAK/STAT3 signaling | 1.617 | 0.027 |
| Apoptosis | 1.601 | 0.029 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Minemura, C.; Asai, S.; Koma, A.; Kikkawa, N.; Kato, M.; Kasamatsu, A.; Uzawa, K.; Hanazawa, T.; Seki, N. Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma. Genes 2022, 13, 1225. https://doi.org/10.3390/genes13071225
Minemura C, Asai S, Koma A, Kikkawa N, Kato M, Kasamatsu A, Uzawa K, Hanazawa T, Seki N. Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma. Genes. 2022; 13(7):1225. https://doi.org/10.3390/genes13071225
Chicago/Turabian StyleMinemura, Chikashi, Shunichi Asai, Ayaka Koma, Naoko Kikkawa, Mayuko Kato, Atsushi Kasamatsu, Katsuhiro Uzawa, Toyoyuki Hanazawa, and Naohiko Seki. 2022. "Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma" Genes 13, no. 7: 1225. https://doi.org/10.3390/genes13071225
APA StyleMinemura, C., Asai, S., Koma, A., Kikkawa, N., Kato, M., Kasamatsu, A., Uzawa, K., Hanazawa, T., & Seki, N. (2022). Identification of Antitumor miR-30e-5p Controlled Genes; Diagnostic and Prognostic Biomarkers for Head and Neck Squamous Cell Carcinoma. Genes, 13(7), 1225. https://doi.org/10.3390/genes13071225