
Potential Misidentification of Natural Isomers and Mass-Analogs of Modified Nucleosides by Liquid Chromatography–Triple Quadrupole Mass Spectrometry

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
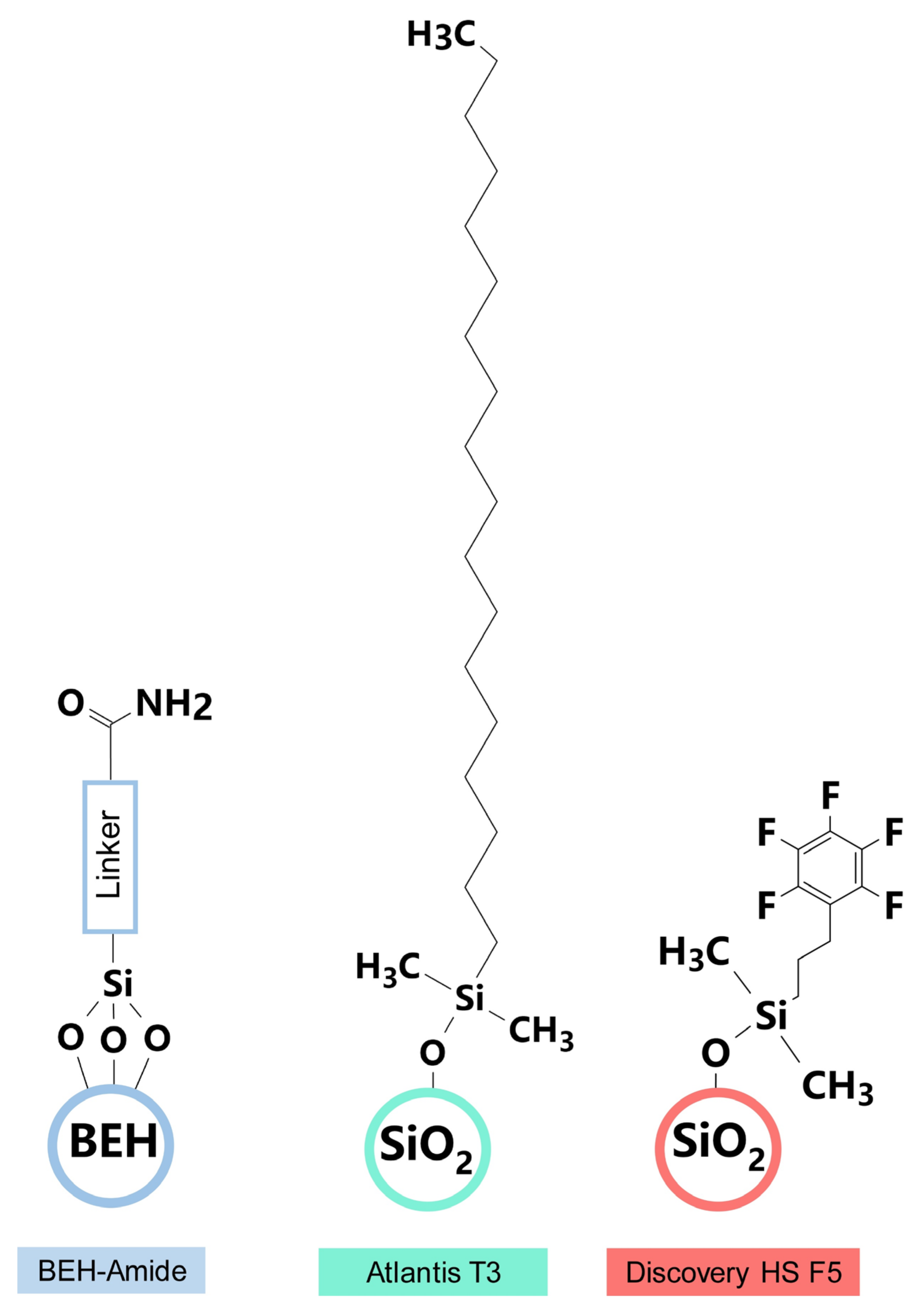
3.1. Analytical Chromatography Columns
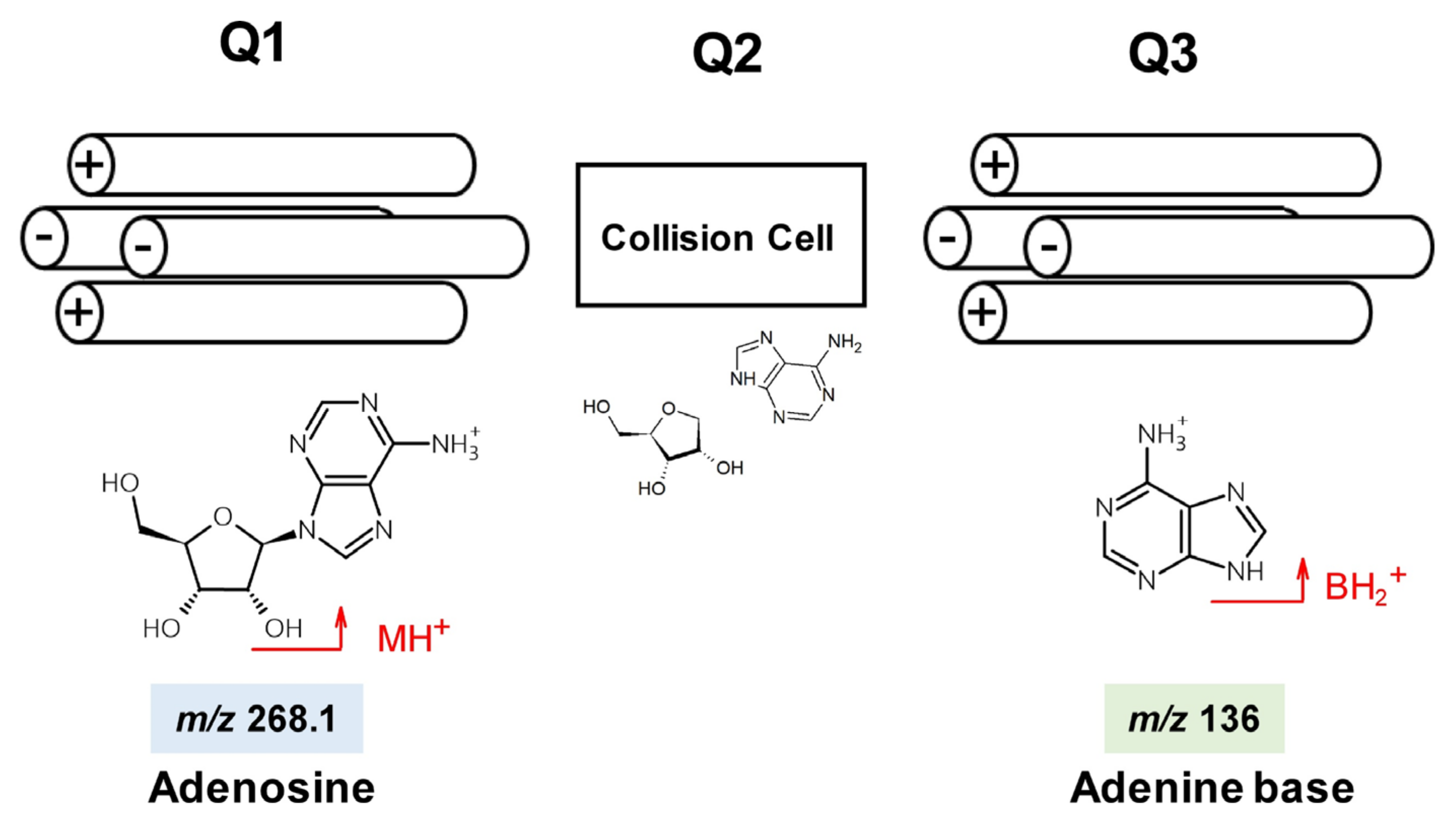
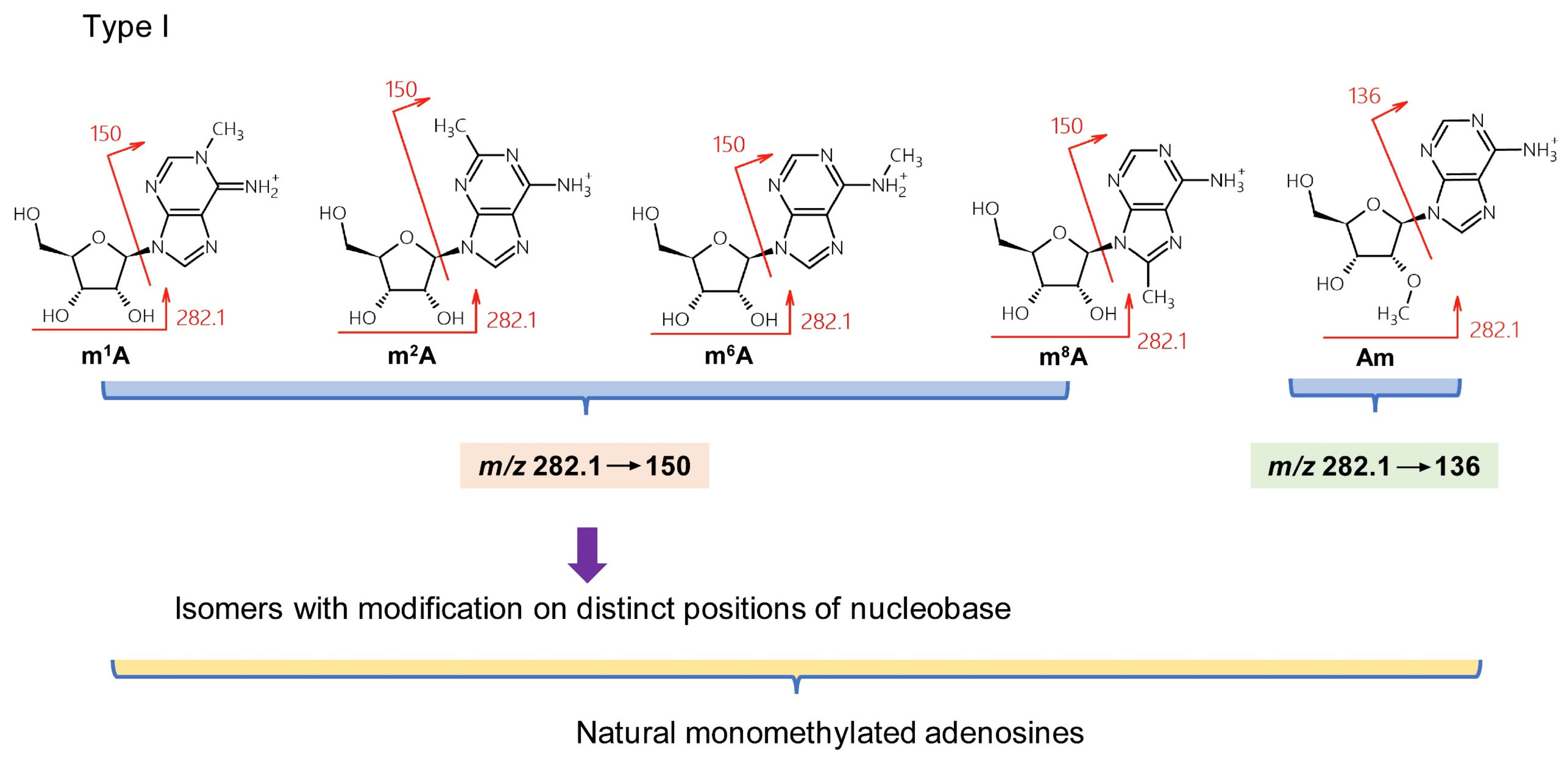
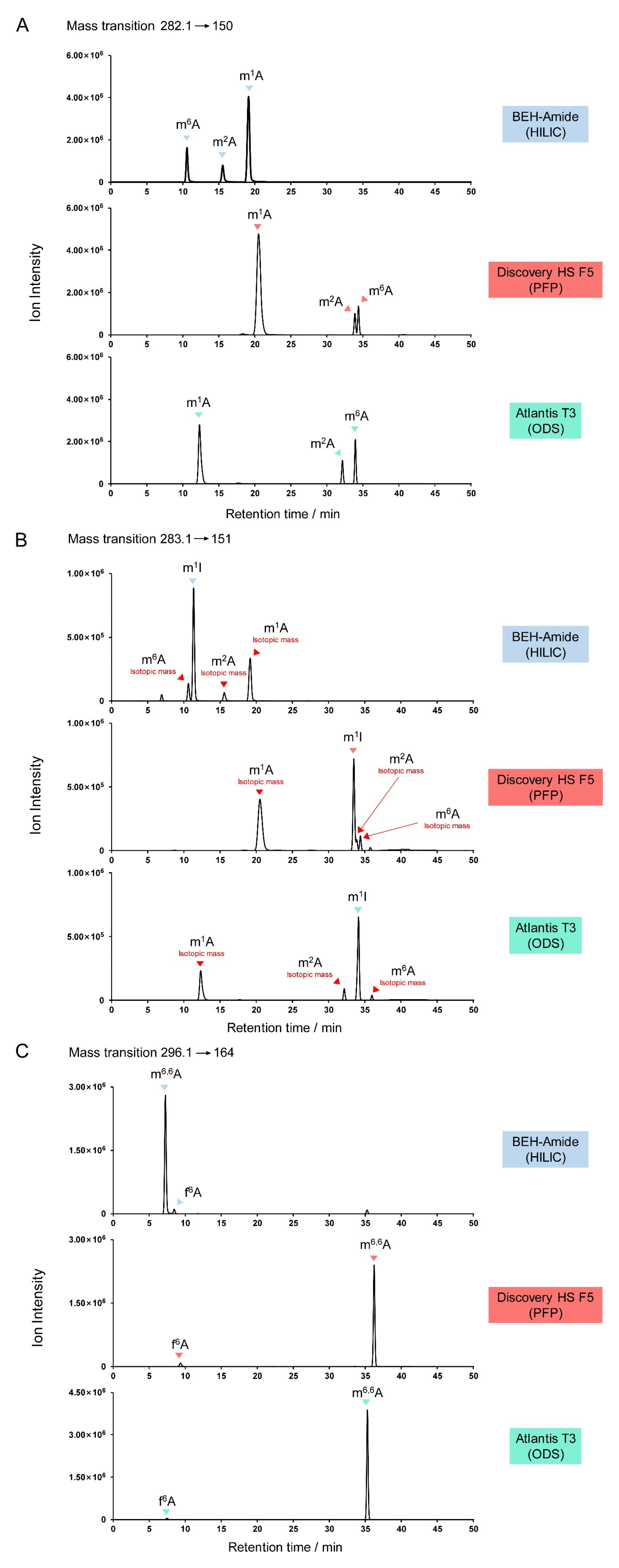
3.2. Adenosine Derivatives
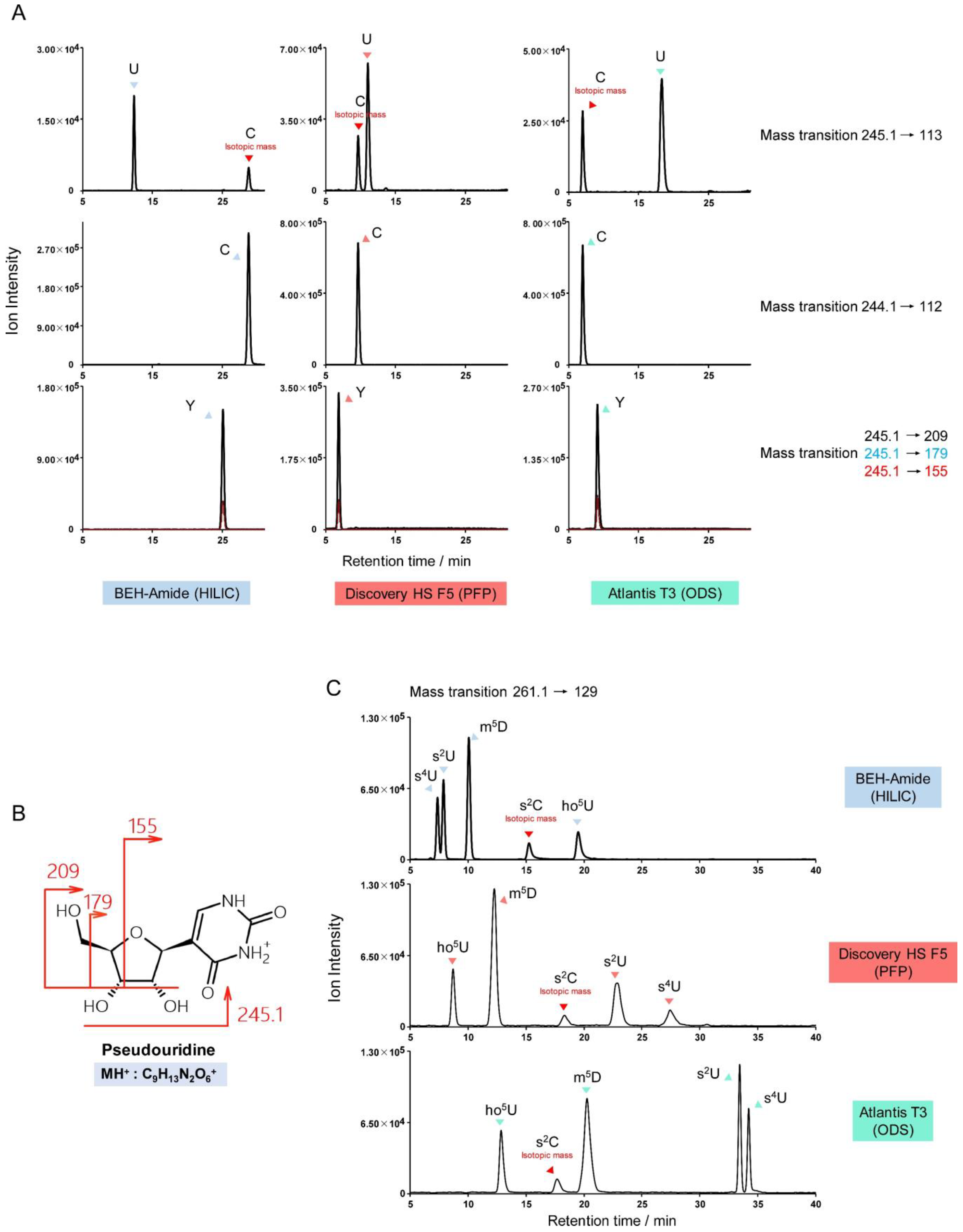
3.3. Uridine and Cytidine Derivatives
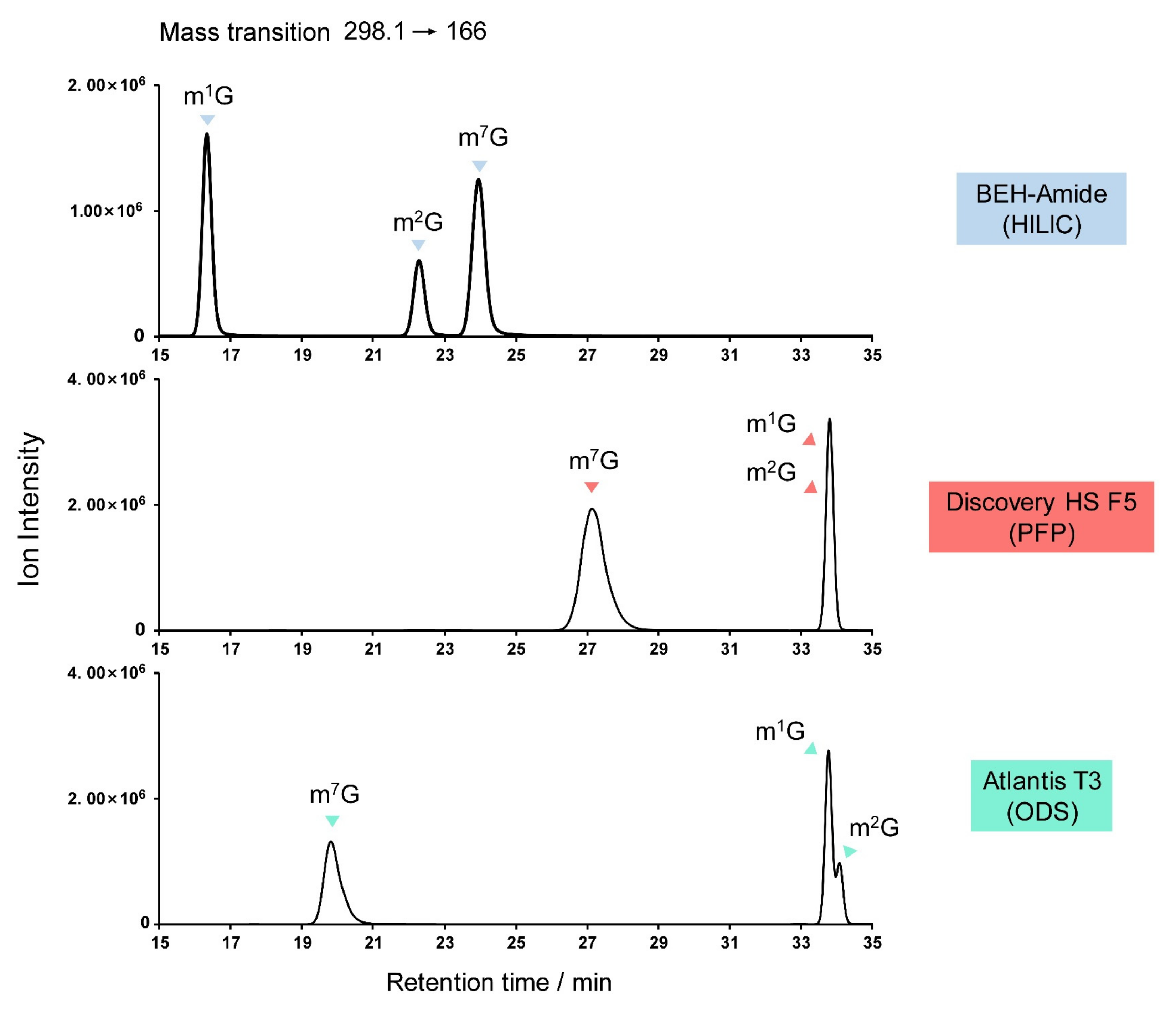
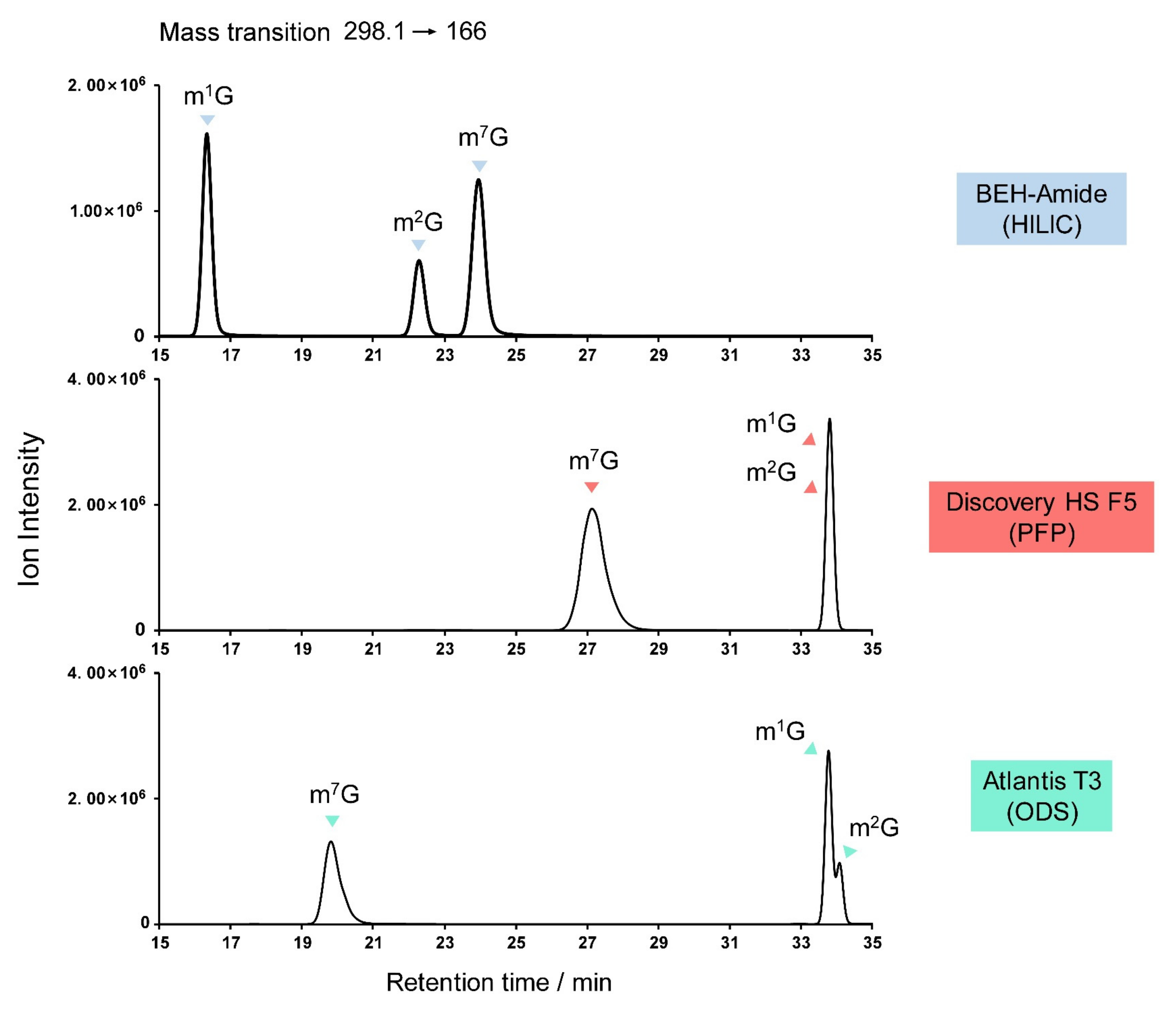
3.4. Guanosine Derivatives
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nachtergaele, S.; He, C. The emerging biology of RNA post-transcriptional modifications. RNA Biol. 2017, 14, 156–163. [Google Scholar] [CrossRef] [PubMed]
- Zhao, B.S.; Roundtree, I.A.; He, C. Post-transcriptional gene regulation by mRNA modifications. Nat. Rev. Mol. Cell Biol. 2017, 18, 31–42. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T. The expanding world of tRNA modifications and their disease relevance. Nat. Rev. Mol. Cell Biol. 2021, 22, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Roundtree, I.A.; Evans, M.E.; Pan, T.; He, C. Dynamic RNA modifications in gene expression regulation. Cell 2017, 169, 1187–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dannfald, A.; Favory, J.-J.; Deragon, J.-M. Variations in transfer and ribosomal RNA epitranscriptomic status can adapt eukaryote translation to changing physiological and environmental conditions. RNA Biol. 2021, 18, 4–18. [Google Scholar] [CrossRef]
- Schwartz, S.; Agarwala, S.D.; Mumbach, M.R.; Jovanovic, M.; Mertins, P.; Shishkin, A.; Tabach, Y.; Mikkelsen, T.S.; Satija, R.; Ruvkun, G. High-resolution mapping reveals a conserved, widespread, dynamic mRNA methylation program in yeast meiosis. Cell 2013, 155, 1409–1421. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Zhao, B.S.; Roundtree, I.A.; Lu, Z.; Han, D.; Ma, H.; Weng, X.; Chen, K.; Shi, H.; He, C. N6-methyladenosine modulates messenger RNA translation efficiency. Cell 2015, 161, 1388–1399. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.; Wan, J.; Gao, X.; Zhang, X.; Jaffrey, S.R.; Qian, S.-B. Dynamic m6A mRNA methylation directs translational control of heat shock response. Nature 2015, 526, 591–594. [Google Scholar] [CrossRef] [Green Version]
- Asano, K.; Suzuki, T.; Saito, A.; Wei, F.-Y.; Ikeuchi, Y.; Numata, T.; Tanaka, R.; Yamane, Y.; Yamamoto, T.; Goto, T.; et al. Metabolic and chemical regulation of tRNA modification associated with taurine deficiency and human disease. Nucleic Acids Res. 2018, 46, 1565–1583. [Google Scholar] [CrossRef] [Green Version]
- Lin, H.; Miyauchi, K.; Harada, T.; Okita, R.; Takeshita, E.; Komaki, H.; Fujioka, K.; Yagasaki, H.; Goto, Y.-i.; Yanaka, K.; et al. CO2-sensitive tRNA modification associated with human mitochondrial disease. Nat. Commun. 2018, 9, 1875. [Google Scholar] [CrossRef]
- Liu, F.; Clark, W.; Luo, G.; Wang, X.; Fu, Y.; Wei, J.; Wang, X.; Hao, Z.; Dai, Q.; Zheng, G.; et al. ALKBH1-mediated tRNA demethylation regulates translation. Cell 2016, 167, 816–828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helm, M.; Motorin, Y. Detecting RNA modifications in the epitranscriptome: Predict and validate. Nat. Rev. Genet. 2017, 18, 275–291. [Google Scholar] [CrossRef] [PubMed]
- Hartstock, K.; Rentmeister, A. Mapping m6A in RNA: Established methods, remaining challenges and emerging approaches. Chemistry 2019, 25, 3455–3464. [Google Scholar] [CrossRef]
- Zhao, L.-Y.; Song, J.; Liu, Y.; Song, C.-X.; Yi, C. Mapping the epigenetic modifications of DNA and RNA. Protein Cell 2020, 11, 792–808. [Google Scholar] [CrossRef]
- Schneeberger, E.M.; Breuker, K. Native top-down mass spectrometry of TAR RNA in complexes with a wild-type tat peptide for binding site mapping. Angew. Chem. Int. Ed. 2017, 129, 1274–1278. [Google Scholar] [CrossRef]
- Schürch, S. Characterization of nucleic acids by tandem mass spectrometry-The second decade (2004–2013): From DNA to RNA and modified sequences. Mass Spectrom. Rev. 2016, 35, 483–523. [Google Scholar] [CrossRef] [PubMed]
- Yoluç, Y.; Ammann, G.; Barraud, P.; Jora, M.; Limbach, P.A.; Motorin, Y.; Marchand, V.; Tisné, C.; Borland, K.; Kellner, S. Instrumental analysis of RNA modifications. Crit. Rev. Biochem. Mol. Biol. 2021, 56, 178–204. [Google Scholar] [CrossRef]
- Catala, M.; Gato, A.; Tisné, C.; Barraud, P. 1 H, 15 N chemical shift assignments of the imino groups of yeast tRNA Phe: Influence of the post-transcriptional modifications. Biomol. NMR Assign. 2020, 14, 169–174. [Google Scholar] [CrossRef]
- Barraud, P.; Gato, A.; Heiss, M.; Catala, M.; Kellner, S.; Tisné, C. Time-resolved NMR monitoring of tRNA maturation. Nat. Commun. 2019, 10, 3373. [Google Scholar] [CrossRef] [Green Version]
- Su, D.; Chan, C.T.; Gu, C.; Lim, K.S.; Chionh, Y.H.; McBee, M.E.; Russell, B.S.; Babu, I.R.; Begley, T.J.; Dedon, P.C. Quantitative analysis of ribonucleoside modifications in tRNA by HPLC-coupled mass spectrometry. Nat. Protoc. 2014, 9, 828–841. [Google Scholar] [CrossRef]
- Miyauchi, K.; Kimura, S.; Suzuki, T. A cyclic form of N6-threonylcarbamoyladenosine as a widely distributed tRNA hypermodification. Nat. Chem. Biol. 2013, 9, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, Y.; Miyauchi, K.; Kang, B.-i.; Suzuki, T. Nucleoside analysis by hydrophilic interaction liquid chromatography coupled with mass spectrometry. Methods Enzymol. 2015, 560, 19–28. [Google Scholar] [PubMed]
- Basanta-Sanchez, M.; Temple, S.; Ansari, S.A.; D’Amico, A.; Agris, P.F. Attomole quantification and global profile of RNA modifications: Epitranscriptome of human neural stem cells. Nucleic Acids Res. 2016, 44, e26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qin, Y.; Zhong, Q.; Zhang, Y.; Lin, X.; Fu, P.; Lin, H. Micro-flow hydrophilic interaction liquid chromatography coupled with triple quadrupole mass spectrometry detects modified nucleosides in the transfer RNA pool of cyanobacteria. J. Sep. Sci. 2021, 44, 3208–3218. [Google Scholar] [CrossRef] [PubMed]
- Chan, C.T.; Dyavaiah, M.; DeMott, M.S.; Taghizadeh, K.; Dedon, P.C.; Begley, T.J. A quantitative systems approach reveals dynamic control of tRNA modifications during cellular stress. PLoS Genet. 2010, 6, e1001247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boccaletto, P.; Machnicka, M.A.; Purta, E.; Piątkowski, P.; Bagiński, B.; Wirecki, T.K.; de Crécy-Lagard, V.; Ross, R.; Limbach, P.A.; Kotter, A. MODOMICS: A database of RNA modification pathways—2017 update. Nucleic Acids Res. 2018, 46, D303–D307. [Google Scholar] [CrossRef]
- Nakayama, H.; Yamauchi, Y.; Nobe, Y.; Sato, K.; Takahashi, N.; Shalev-Benami, M.; Isobe, T.; Taoka, M. Method for direct mass-spectrometry-based identification of monomethylated RNA nucleoside positional isomers and its application to the analysis of Leishmania rRNA. Anal. Chem. 2019, 91, 15634–15643. [Google Scholar] [CrossRef] [Green Version]
- Fu, D.; Brophy, J.A.; Chan, C.T.; Atmore, K.A.; Begley, U.; Paules, R.S.; Dedon, P.C.; Begley, T.J.; Samson, L.D. Human AlkB homolog ABH8 Is a tRNA methyltransferase required for wobble uridine modification and DNA damage survival. Mol. Cell. Biol. 2010, 30, 2449–2459. [Google Scholar] [CrossRef] [Green Version]
- Pastore, C.; Topalidou, I.; Forouhar, F.; Yan, A.C.; Levy, M.; Hunt, J.F. Crystal structure and RNA binding properties of the RNA recognition motif (RRM) and AlkB domains in human AlkB homolog 8 (ABH8), an enzyme catalyzing tRNA hypermodification. J. Biol. Chem. 2012, 287, 2130–2143. [Google Scholar] [CrossRef] [Green Version]
- Patil, A.; Chan, C.; Dyavaiah, M.; Rooney, J.P.; Dedon, P. Translational infidelity-induced protein stress results from a deficiency in Trm9-catalyzed tRNA modifications. RNA Biol. 2012, 9, 990–1001. [Google Scholar] [CrossRef] [Green Version]
- Ghfar, A.A.; Wabaidur, S.M.; Ahmed, A.Y.B.H.; Alothman, Z.A.; Khan, M.R.; Al-Shaalan, N.H. Simultaneous determination of monosaccharides and oligosaccharides in dates using liquid chromatography–electrospray ionization mass spectrometry. Food Chem. 2015, 176, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Portillo-Castillo, O.J.; Castro-Ríos, R.; Chávez-Montes, A.; González-Horta, A.; Cavazos-Rocha, N.; Waksman De Torres, N.; Garza-Tapia, M. A new RP-HPLC method as an auxiliary tool for optimization of sample preparation procedures for tracing of PPCPs of different hydrophilicities. Acta Pharm. 2021, 71, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.; Herold, N.; Keppler, O.T.; Geisslinger, G.; Ferreirós, N. Quantitation of endogenous nucleoside triphosphates and nucleosides in human cells by liquid chromatography tandem mass spectrometry. Anal. Bioanal. Chem. 2015, 407, 3693–3704. [Google Scholar] [CrossRef] [PubMed]
- Dudley, E.; Tuytten, R.; Bond, A.; Lemière, F.; Brenton, A.G.; Esmans, E.L.; Newton, R.P. Study of the mass spectrometric fragmentation of pseudouridine: Comparison of fragmentation data obtained by matrix-assisted laser desorption/ionisation post-source decay, electrospray ion trap multistage mass spectrometry, and by a method utilising electrospray quadrupole time-of-flight tandem mass spectrometry and in-source fragmentation. Rapid Commun. Mass Spectrom. 2005, 19, 3075–3085. [Google Scholar]
- Kellner, S.; Neumann, J.; Rosenkranz, D.; Lebedeva, S.; Ketting, R.F.; Zischler, H.; Schneider, D.; Helm, M. Profiling of RNA modifications by multiplexed stable isotope labelling. Chem. Comm. 2014, 50, 3516–3518. [Google Scholar] [CrossRef]
- Kellner, S.; Ochel, A.; Thüring, K.; Spenkuch, F.; Neumann, J.; Sharma, S.; Entian, K.-D.; Schneider, D.; Helm, M. Absolute and relative quantification of RNA modifications via biosynthetic isotopomers. Nucleic Acids Res. 2014, 42, e142. [Google Scholar] [CrossRef] [Green Version]
- Kaiser, S.; Byrne, S.R.; Ammann, G.; Asadi Atoi, P.; Borland, K.; Brecheisen, R.; DeMott, M.S.; Gehrke, T.; Hagelskamp, F.; Heiss, M.; et al. Strategies to Avoid Artifacts in Mass Spectrometry-Based Epitranscriptome Analyses. Angew. Chem. Int. Ed. 2021, 60, 23885–23893. [Google Scholar] [CrossRef]
- Lopez Torres, A.; Yanez Barrientos, E.; Wrobel, K.; Wrobel, K. Selective derivatization of cytosine and methylcytosine moieties with 2-bromoacetophenone for submicrogram DNA methylation analysis by reversed phase HPLC with spectrofluorimetric detection. Anal. Chem. 2011, 83, 7999–8005. [Google Scholar] [CrossRef]
- Estevez, M.; Valesyan, S.; Jora, M.; Limbach, P.A.; Addepalli, B. Oxidative damage to RNA is altered by the presence of interacting proteins or modified nucleosides. Front. Mol. Biosci. 2021, 8, 631. [Google Scholar] [CrossRef]
- Jora, M.; Borland, K.; Abernathy, S.; Zhao, R.; Kelley, M.; Kellner, S.; Addepalli, B.; Limbach, P.A. Chemical amination/imination of carbonothiolated nucleosides during RNA hydrolysis. Angew. Chem. Int. Ed. 2021, 60, 3961–3966. [Google Scholar] [CrossRef]
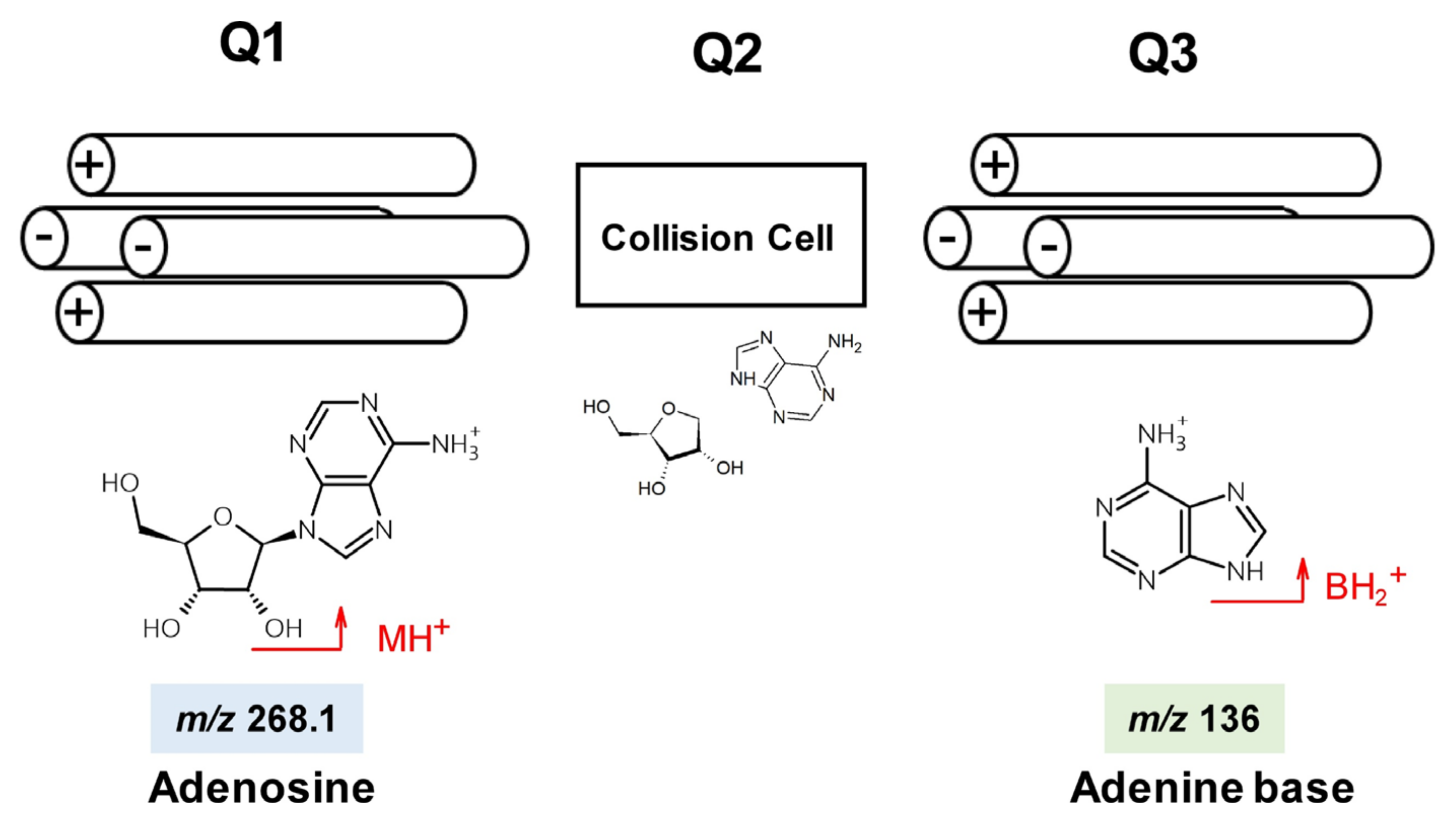
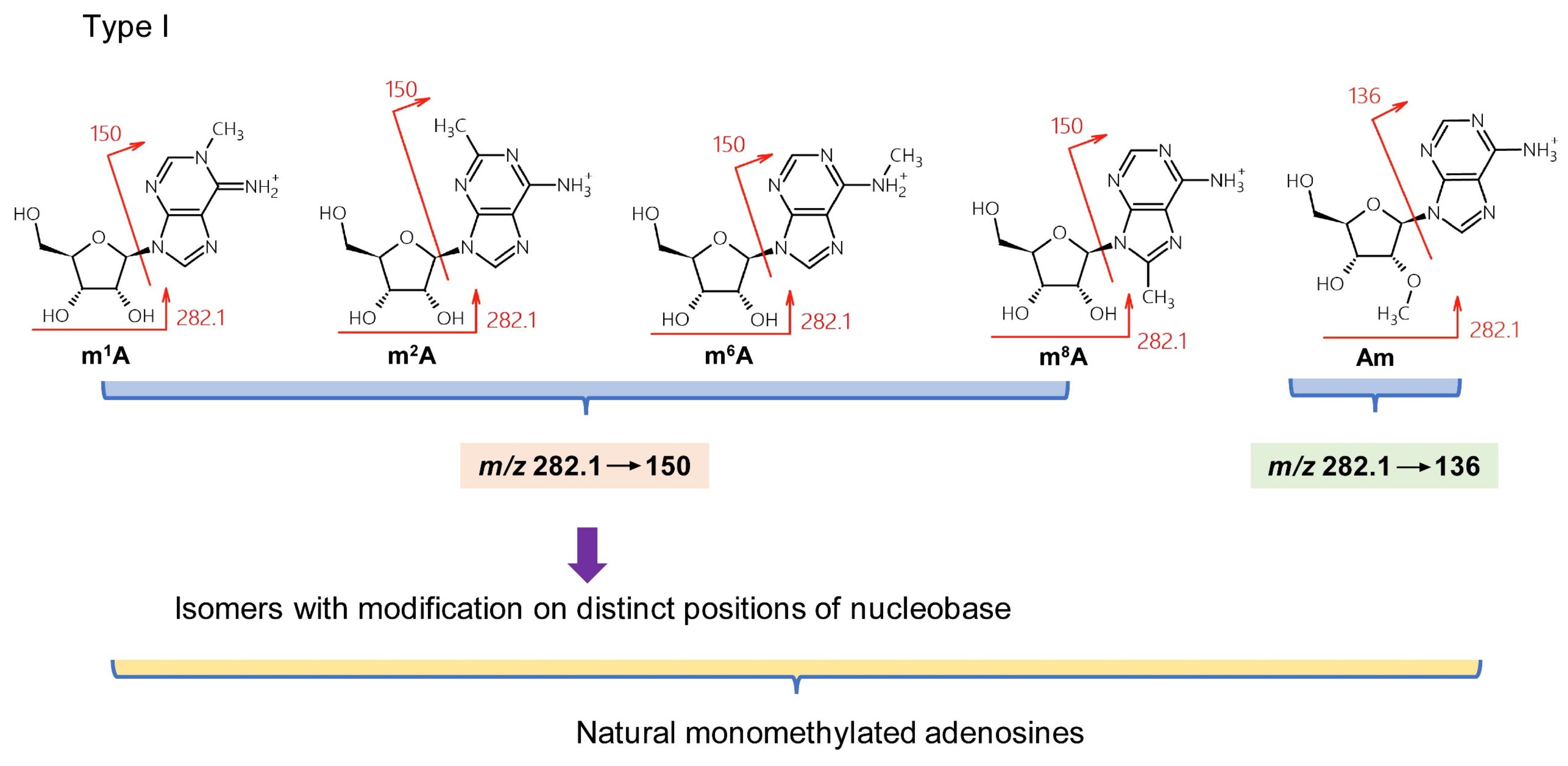
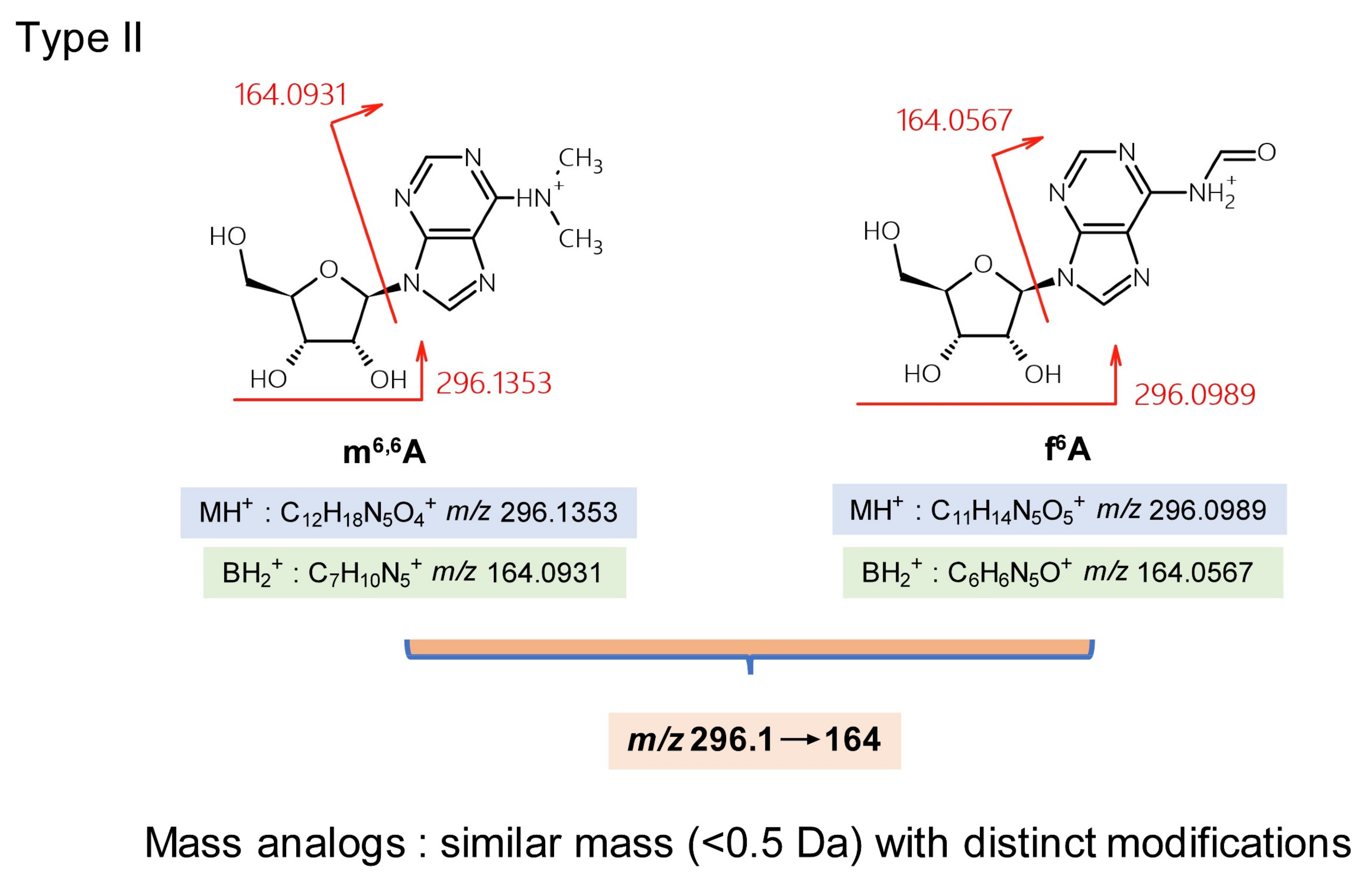
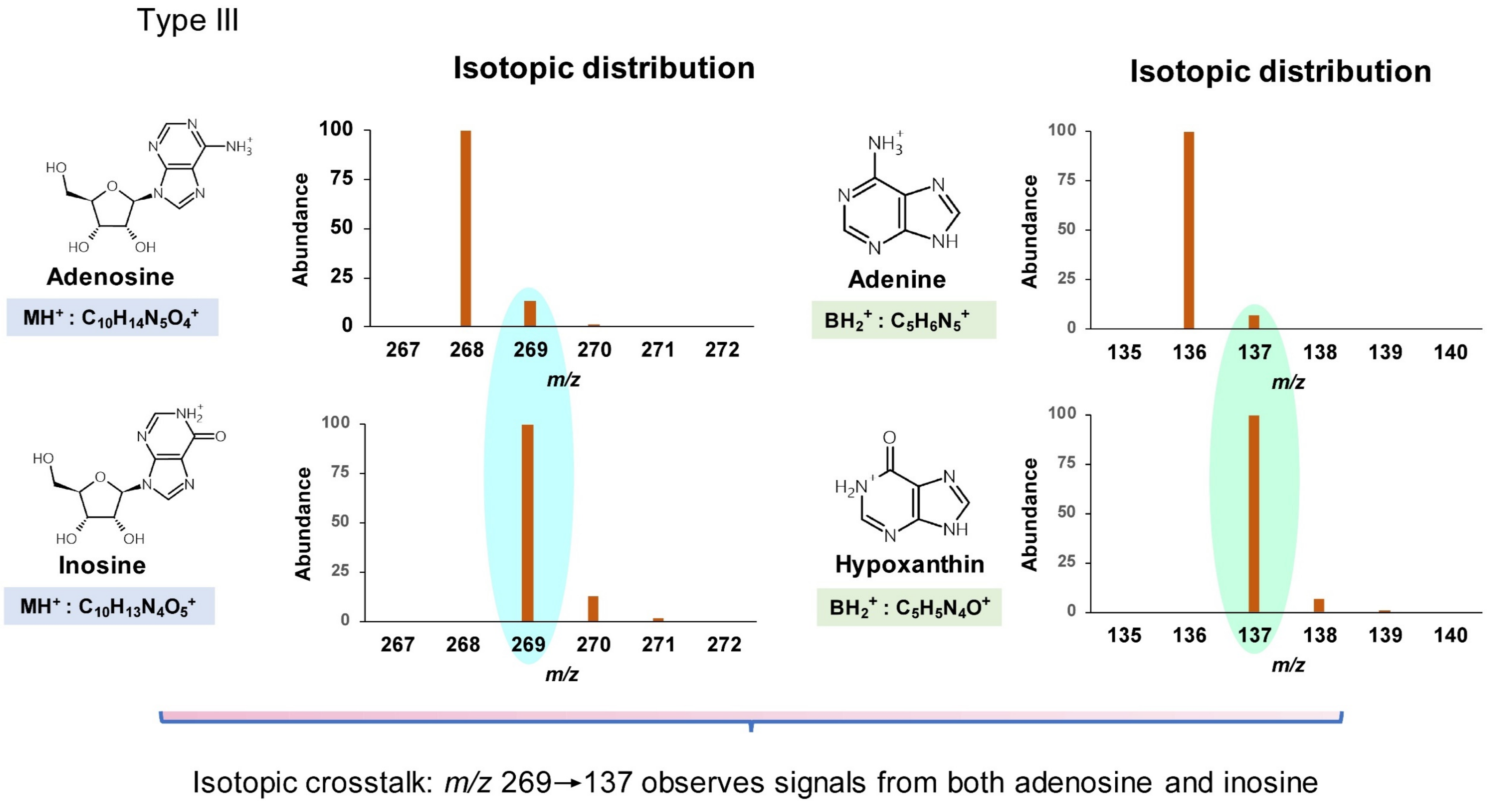




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Abbreviation | Chemical Formula | Precursor Ion | Product Ion |
|---|---|---|---|---|
| Adenosine derivatives | ||||
| 1-Methyladenosine | m1A | C11H15N5O4 | 282.1 | 150 |
| 2-Methyladenosine | m2A | C11H15N5O4 | 282.1 | 150 |
| N6-Methyladenosine | m6A | C11H15N5O4 | 282.1 | 150 |
| 8-Methyladenosine | m8A | C11H15N5O4 | 282.1 | 150 |
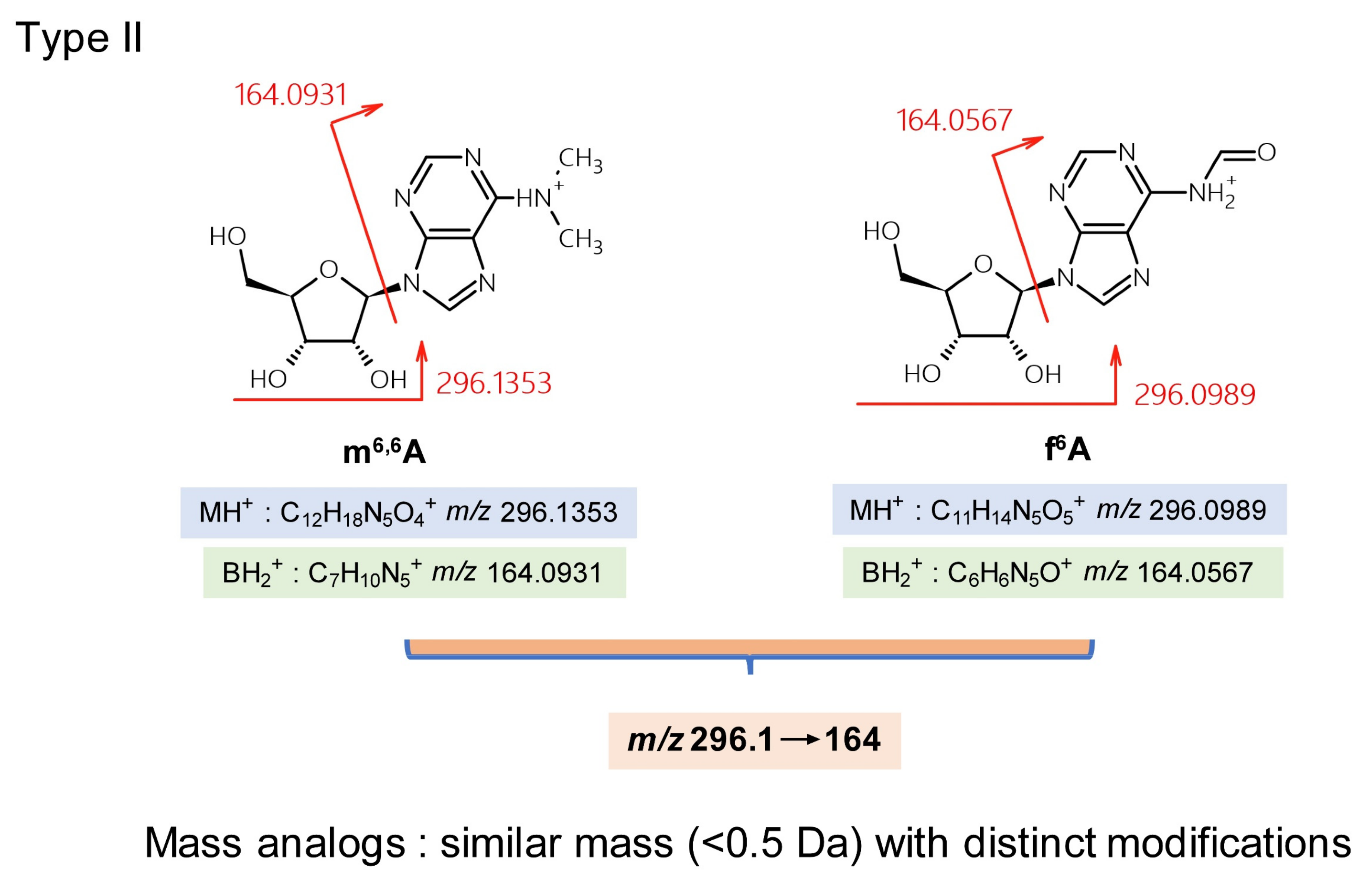
| 2,8-Dimethyladenosine | m2,8A | C12H17N5O4 | 296.1 | 164 |
| N6,N6-Dimethyladenosine | m6,6A | C12H17N5O4 | 296.1 | 164 |
| Cytidine and Uridine derivatives | ||||
| 3-Methylcytidine | m3C | C10H15N3O5 | 258.1 | 126 |
| N4-Methylcytidine | m4C | C10H15N3O5 | 258.1 | 126 |
| 5-Methylcytidine | m5C | C10H15N3O5 | 258.1 | 126 |
| 3-Methyluridine | m3U | C10H14N2O6 | 259.1 | 127 |
| 5-Methyluridine | m5U | C10H14N2O6 | 259.1 | 127 |
| 1-Methylpseudouridine | m1Y | C10H14N2O6 | 259.1 | 179 |
| 3-Methylpseudouridine | m3Y | C10H14N2O6 | 259.1 | 179 |
| 2-Thiouridine | s2U | C9H12N2O5S | 261.1 | 129 |
| 4-Thiouridine | s4U | C9H12N2O5S | 261.1 | 129 |
| 5,2′-O-Dimethylcytidine | m5Cm | C11H17N3O5 | 272.1 | 126 |
| N4,2′-O-Dimethylcytidine | m4Cm | C11H17N3O5 | 272.1 | 126 |
| 3,2′-O-Dimethyluridine | m3Um | C11H16N2O6 | 273.1 | 127 |
| 5,2′-O-Dimethyluridine | m5Um | C11H16N2O6 | 273.1 | 127 |
| 5-Carboxyhydroxymethyluridine | chm5U | C11H14N2O9 | 319.1 | 187 |
| Uridine 5-oxyacetic acid | cmo5U | C11H14N2O9 | 319.1 | 187 |
| 5-(Carboxyhydroxymethyl)uridine methyl ester | mchm5U | C12H16N2O9 | 333.1 | 297 |
| Uridine 5-oxyacetic acid methyl ester | mcmo5U | C12H16N2O9 | 333.1 | 297 |
| 3-(3-amino-3-carboxypropyl)pseudouridine | acp3Y | C13H19N3O8 | 346.1 | 214 |
| 3-(3-amino-3-carboxypropyl)uridine | acp3U | C13H19N3O8 | 346.1 | 214 |
| Guanosine derivatives | ||||
| 1-Methylguanosine | m1G | C11H15N5O5 | 298.1 | 166 |
| N2-Methylguanosine | m2G | C11H15N5O5 | 298.1 | 166 |
| 7-methylguanosine | m7G | C11H15N5O5 | 298.1 | 166 |
| 1,2′-O-Dimethylguanosine | m1Gm | C12H17N5O5 | 312.1 | 166 |
| N2,2′-O-Dimethylguanosine | m2Gm | C12H17N5O5 | 312.1 | 166 |
| N2,7-Dimethylguanosine | m2,7G | C12H17N5O5 | 312.1 | 180 |
| N2,N2-Dimethylguanosine | m2,2G | C12H17N5O5 | 312.1 | 180 |
| N2,N2,2′-O-Trimethylguanosine | m2,2Gm | C13H19N5O5 | 326.1 | 180 |
| N2,7,2′-O-Trimethylguanosine | m2,7Gm | C13H19N5O5 | 326.1 | 180 |
| Isowyosine | imG2 | C14H17N5O5 | 336.1 | 204 |
| Wyosine | imG | C14H17N5O5 | 336.1 | 204 |
| Name | Abbreviation | Chemical Formula | Precursor Ion | Product Ion |
|---|---|---|---|---|
| Adenosine derivatives | ||||
| N6-Formyladenosine | f6A | C11H13N5O5 | 269.1 | 164 |
| 2,8-Dimethyladenosine | m2,8A | C12H17N5O4 | 269.1 | 164 |
| N6,N6-Dimethyladenosine | m6,6A | C12H17N5O4 | 269.1 | 164 |
| N6-Methyl-N6-threonylcarbamoyladenosine | m6t6A | C15H20N6O8 | 427.2 | 295 |
| N6-Hydroxynorvalylcarbamoyladenosine | hn6A | C16H22N6O8 | 427.2 | 295 |
| Cytidine and Uridine derivatives | ||||
| 2-Thiocytidine | s2C | C9H13N3O4S | 260.1 | 128 |
| 5-Hydroxycytidine | ho5C | C9H13N3O6 | 260.1 | 128 |
| 2-Thiouridine | s2U | C9H12N2O5S | 261.1 | 129 |
| 4-Thiouridine | s4U | C9H12N2O5S | 261.1 | 129 |
| 5-Hydroxyuridine | ho5U | C9H12N2O7 | 261.1 | 129 |
| 5-Methyldihydrouridine | m5D | C10H16N2O6 | 261.1 | 129 |
| 5-Formylcytidine | f5C | C10H13N3O6 | 272.1 | 140 |
| N4,N4-Dimethylcytidine | m4,4C | C11H17N3O5 | 272.1 | 140 |
| 5-Methyl-2-thiouridine | m5s2U | C10H14N2O5S | 275.1 | 143 |
| 5-Methoxyuridine | mo5U | C10H14N2O7 | 275.1 | 143 |
| 5-Formyl-2′-O-methylcytidine | f5Cm | C11H15N3O6 | 286.1 | 140 |
| N4,N4,2′-O-Trimethylcytidine | m4,4Cm | C12H19N3O5 | 286.1 | 140 |
| 5-Carbamoylmethyl-2-thiouridine | ncm5s2U | C11H15N3O6S | 318.1 | 186 |
| 5-Carbamoylhydroxymethyluridine | nchm5U | C11H15N3O8 | 318.1 | 186 |
| 5-Carboxymethyl-2-thiouridine | cm5s2U | C11H14N2O7S | 319.1 | 187 |
| 5-Carboxyhydroxymethyluridine | chm5U | C11H14N2O9 | 319.1 | 187 |
| Uridine 5-oxyacetic acid | cmo5U | C11H14N2O9 | 319.1 | 187 |
| 5-Methoxycarbonylmethyl-2-thiouridine | mcm5s2U | C12H16N2O7S | 333.1 | 201 |
| 5-(carboxyhydroxymethyl)uridine methyl ester | mchm5U | C12H16N2O9 | 333.1 | 201 |
| Uridine 5-oxyacetic acid methyl ester | mcmo5U | C12H16N2O9 | 333.1 | 201 |
| 5-Carboxymethylaminomethyl-2-thiouridine | cmnm5s2U | C12H17N3O7S | 348.1 | 216 |
| 3-(3-amino-3-carboxypropyl)-5,6-dihydrouridine | acp3D | C13H21N3O8 | 348.1 | 216 |
| Name | Abbreviation | Chemical Formula | Precursor Ion | Product Ion |
|---|---|---|---|---|
| Adenosine derivatives | ||||
| 1-Methyladenosine | m1A | C11H15N5O4 | 282.1 | 150 |
| 2-Methyladenosine | m2A | C11H15N5O4 | 282.1 | 150 |
| N6-Methyladenosine | m6A | C11H15N5O4 | 282.1 | 150 |
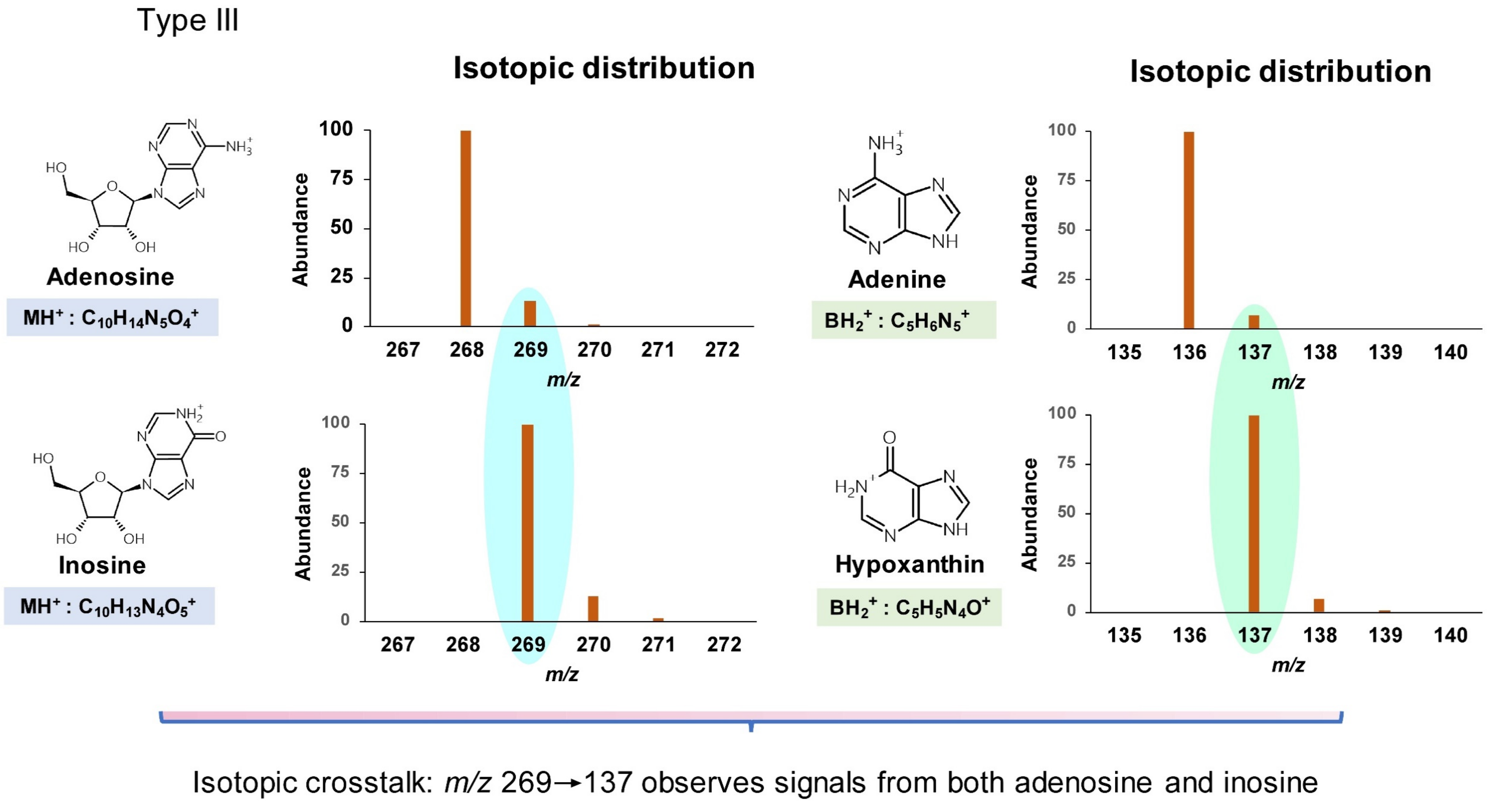
| 1-Methylinosine | m1I | C11H14N4O5 | 283.1 | 151 |
| 2′-O-Methyladenosine | Am | C11H15N5O4 | 282.1 | 136 |
| 2′-O-Methylinosine | Im | C11H14N4O5 | 283.1 | 137 |
| 1,2′-O-Dimethyladenosine | m1Am | C12H17N5O4 | 296.1 | 150 |
| 1,2′-O-Dimethylinosine | m1Im | C12H16N4O5 | 297.1 | 151 |
| Cytidine and Uridine derivatives | ||||
| 3-Methylcytidine | m3C | C10H15N3O5 | 258.1 | 126 |
| 5-Methylcytidine | m5C | C10H15N3O5 | 258.1 | 126 |
| N4-Methylcytidine | m4C | C10H15N3O5 | 258.1 | 126 |
| 3-Methyluridine | m3U | C10H14N2O6 | 259.1 | 127 |
| 5-Methyluridine | m5U | C10H14N2O6 | 259.1 | 127 |
| 2-Thiocytidine | s2C | C9H13N3O4S | 260.1 | 128 |
| 5-Hydroxycytidine | ho5C | C9H13N3O6 | 260.1 | 128 |
| 2-Thiouridine | s2U | C9H12N2O5S | 261.1 | 129 |
| 4-Thiouridine | s4U | C9H12N2O5S | 261.1 | 129 |
| 5-Hydroxyuridine | ho5U | C9H12N2O7 | 261.1 | 129 |
| 5-Methyldihydrouridine | m5D | C10H16N2O6 | 261.1 | 129 |
| 5,2′-O-Dimethylcytidine | m5Cm | C11H17N3O5 | 272.1 | 126 |
| N4,2′-O-Dimethylcytidine | m4Cm | C11H17N3O5 | 272.1 | 126 |
| 3,2′-O-Dimethyluridine | m3Um | C11H16N2O6 | 273.1 | 127 |
| 5,2′-O-Dimethyluridine | m5Um | C11H16N2O6 | 273.1 | 127 |
| 5-Hydroxymethylcytidine | hm5C | C10H15N3O6 | 274.1 | 142 |
| 5-Aminomethyluridine | nm5U | C10H15N3O6 | 274.1 | 142 |
| 5-Methyl-2-thiouridine | m5s2U | C10H14N2O5S | 275.1 | 143 |
| 5-Methoxyuridine | mo5U | C10H14N2O7 | 275.1 | 143 |
| 5-Cyanomethyluridine | cnm5U | C11H13N3O6 | 284.1 | 152 |
| N4-Acetylcytidine | ac4C | C11H15N3O6 | 286.1 | 154 |
| 5-Formyl-2′-O-methylcytidine | f5Cm | C11H15N3O6 | 286.1 | 140 |
| N4,N4,2′-O-Trimethylcytidine | m4,4Cm | C12H19N3O5 | 286.1 | 140 |
| 2′-O-Methyl-5-hydroxymethylcytidine | hm5Cm | C11H17N3O6 | 288.1 | 142 |
| 5-Methylaminomethyluridine | mnm5U | C11H17N3O6 | 288.1 | 156 |
| 5-Aminomethyl-2-thiouridine | nm5s2U | C10H15N3O5S | 290.1 | 158 |
| 5-Carbamoylmethyluridine | ncm5U | C11H15N3O7 | 302.1 | 170 |
| 5-Carboxymethyluridine | cm5U | C11H14N2O8 | 303.1 | 171 |
| 5-Methylaminomethyl-2-thiouridine | mnm5s2U | C11H17N3O5S | 304.1 | 172 |
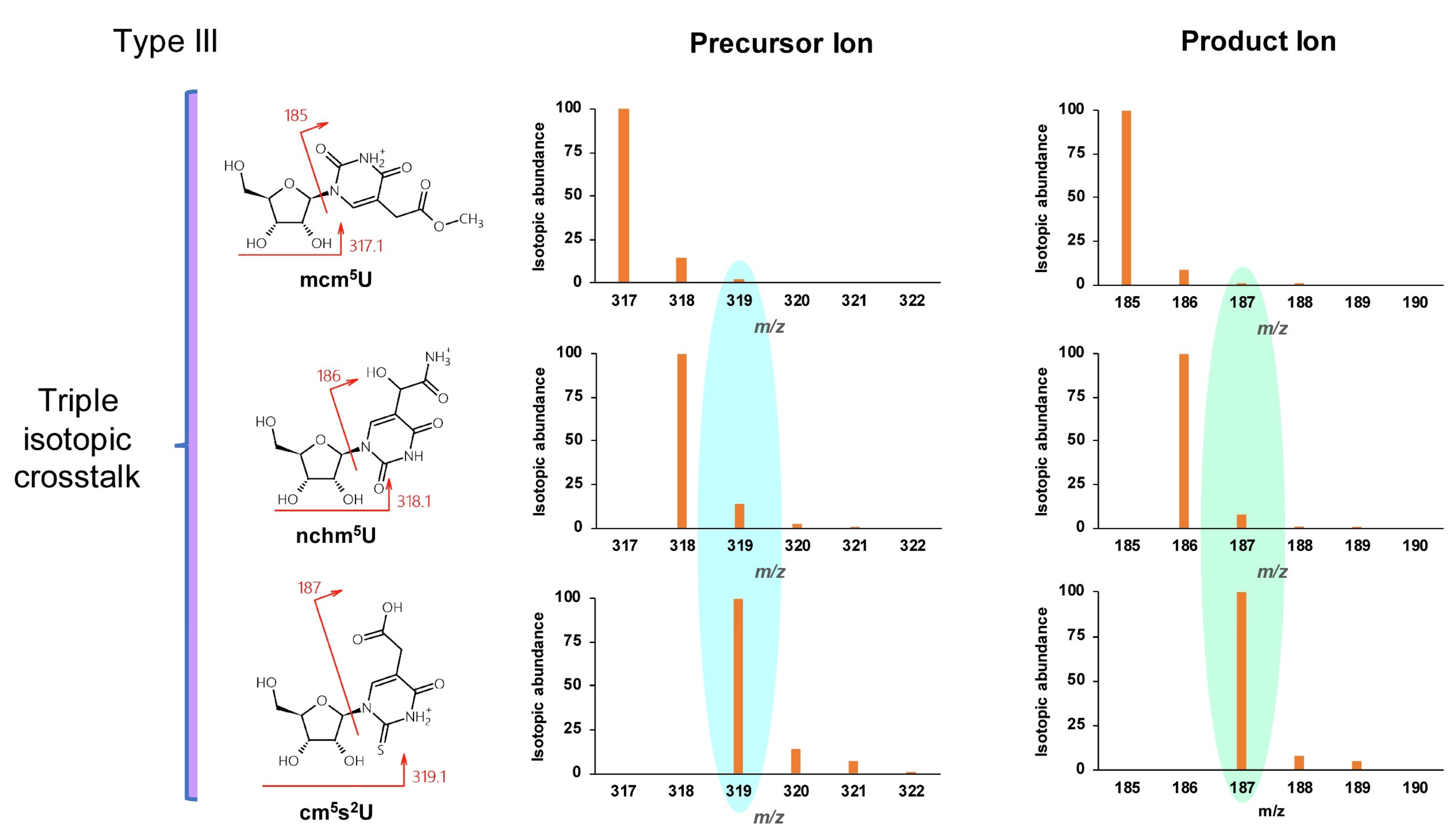
| 5-Methoxycarbonylmethyluridine | mcm5U | C12H16N2O8 | 317.1 | 185 |
| 5-Carbamoylmethyl-2-thiouridine | ncm5s2U | C11H15N3O6S | 318.1 | 186 |
| 5-Carbamoylhydroxymethyluridine | nchm5U | C11H15N3O8 | 318.1 | 186 |
| 5-Carboxymethyl-2-thiouridine | cm5s2U | C11H14N2O7S | 319.1 | 187 |
| 5-Carboxyhydroxymethyluridine | chm5U | C11H14N2O9 | 319.1 | 187 |
| Uridine 5-oxyacetic acid | cmo5U | C11H14N2O9 | 319.1 | 187 |
| 5-Carboxymethylaminomethyluridine | cmnm5U | C12H17N3O8 | 332.1 | 200 |
| 5-Methoxycarbonylmethyl-2-thiouridine | mcm5s2U | C12H16N2O7S | 333.1 | 201 |
| 5-(carboxyhydroxymethyl)uridine methyl ester | mchm5U | C12H16N2O9 | 333.1 | 201 |
| Uridine 5-oxyacetic acid methyl ester | mcmo5U | C12H16N2O9 | 333.1 | 201 |
| 5-Carboxymethylaminomethyl-2′-O-methylu-ridine | cmnm5Um | C13H19N3O8 | 346.1 | 200 |
| 5-(carboxyhydroxymethyl)-2′-O-methyluridi-ne methyl ester | mchm5Um | C13H18N2O9 | 347.1 | 201 |
| 2′-O-Methyluridine 5-oxyacetic acid methyl ester | mcmo5Um | C13H18N2O9 | 347.1 | 201 |
| 5-(isopentenylaminomethyl)-2-thiouridine | inm5s2U | C15H23N3O5S | 358.1 | 226 |
| 1-Methyl-3-(3-amino-3-carboxypropyl)pseud-ouridine | m1acp3Y | C14H21N3O8 | 360.1 | 228 |
| Guanosine derivatives | ||||
| 7-Aminocarboxypropylwyosine methyl ester | yW-58 | C19H26N6O7 | 451.2 | 319 |
| Undermodified hydroxywybutosine | OHyWx | C18H24N6O8 | 453.2 | 321 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, X.; Zhang, Q.; Qin, Y.; Zhong, Q.; Lv, D.; Wu, X.; Fu, P.; Lin, H. Potential Misidentification of Natural Isomers and Mass-Analogs of Modified Nucleosides by Liquid Chromatography–Triple Quadrupole Mass Spectrometry. Genes 2022, 13, 878. https://doi.org/10.3390/genes13050878
Lin X, Zhang Q, Qin Y, Zhong Q, Lv D, Wu X, Fu P, Lin H. Potential Misidentification of Natural Isomers and Mass-Analogs of Modified Nucleosides by Liquid Chromatography–Triple Quadrupole Mass Spectrometry. Genes. 2022; 13(5):878. https://doi.org/10.3390/genes13050878
Chicago/Turabian StyleLin, Xiuying, Qianhui Zhang, Yichao Qin, Qisheng Zhong, Daizhu Lv, Xiaopeng Wu, Pengcheng Fu, and Huan Lin. 2022. "Potential Misidentification of Natural Isomers and Mass-Analogs of Modified Nucleosides by Liquid Chromatography–Triple Quadrupole Mass Spectrometry" Genes 13, no. 5: 878. https://doi.org/10.3390/genes13050878
APA StyleLin, X., Zhang, Q., Qin, Y., Zhong, Q., Lv, D., Wu, X., Fu, P., & Lin, H. (2022). Potential Misidentification of Natural Isomers and Mass-Analogs of Modified Nucleosides by Liquid Chromatography–Triple Quadrupole Mass Spectrometry. Genes, 13(5), 878. https://doi.org/10.3390/genes13050878