
Casticin Impacts Key Signaling Pathways in Colorectal Cancer Cells Leading to Cell Death with Therapeutic Implications


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Materials
2.2. Cytotoxicity Assessment of Casticin on Colorectal Cancer Cell Lines DLD-1, HCT116 and Caco-2
2.3. Clonogenic Assay
2.4. Muse® Flow Cytometric Analysis
2.5. Statistical Analysis
3. Results
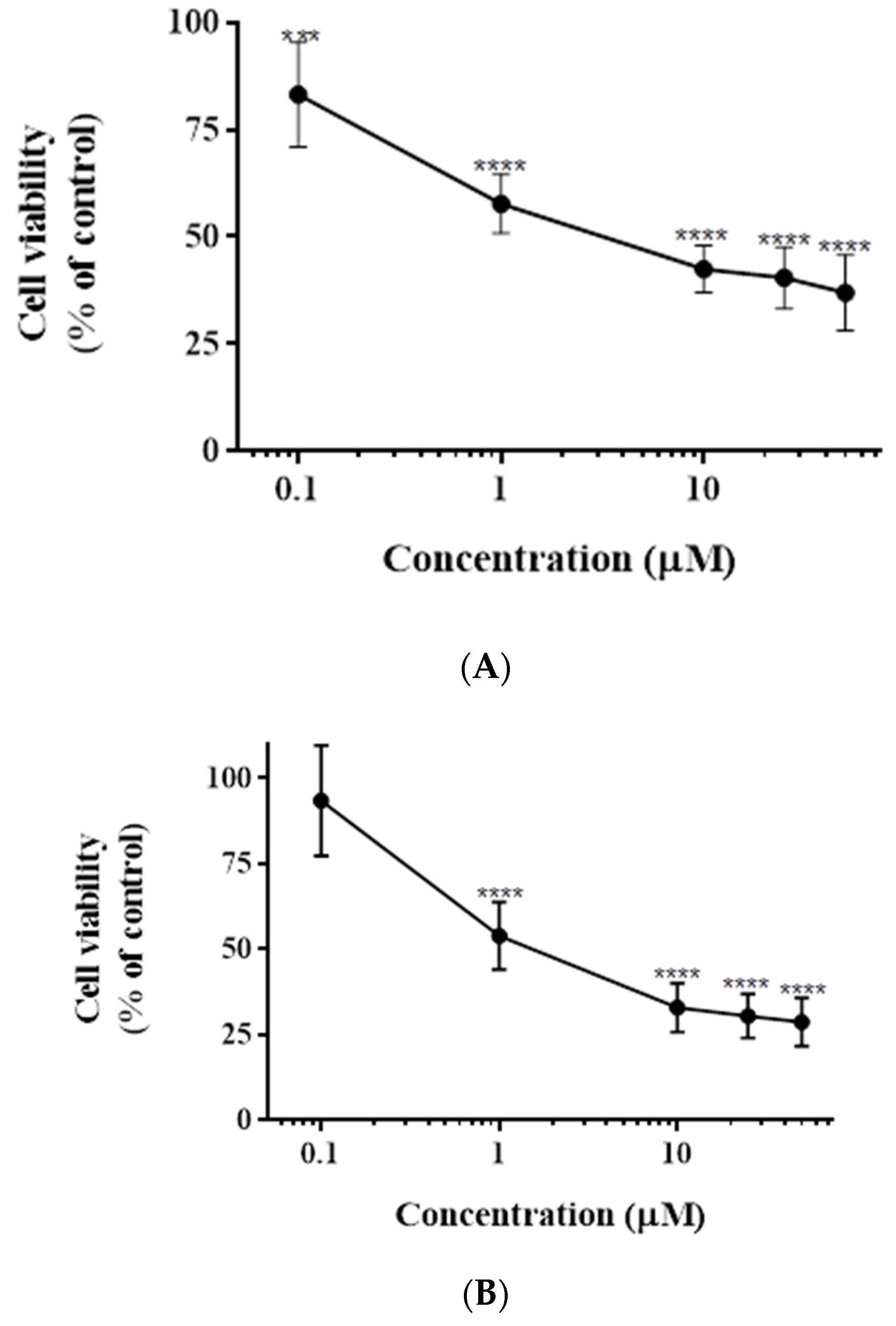
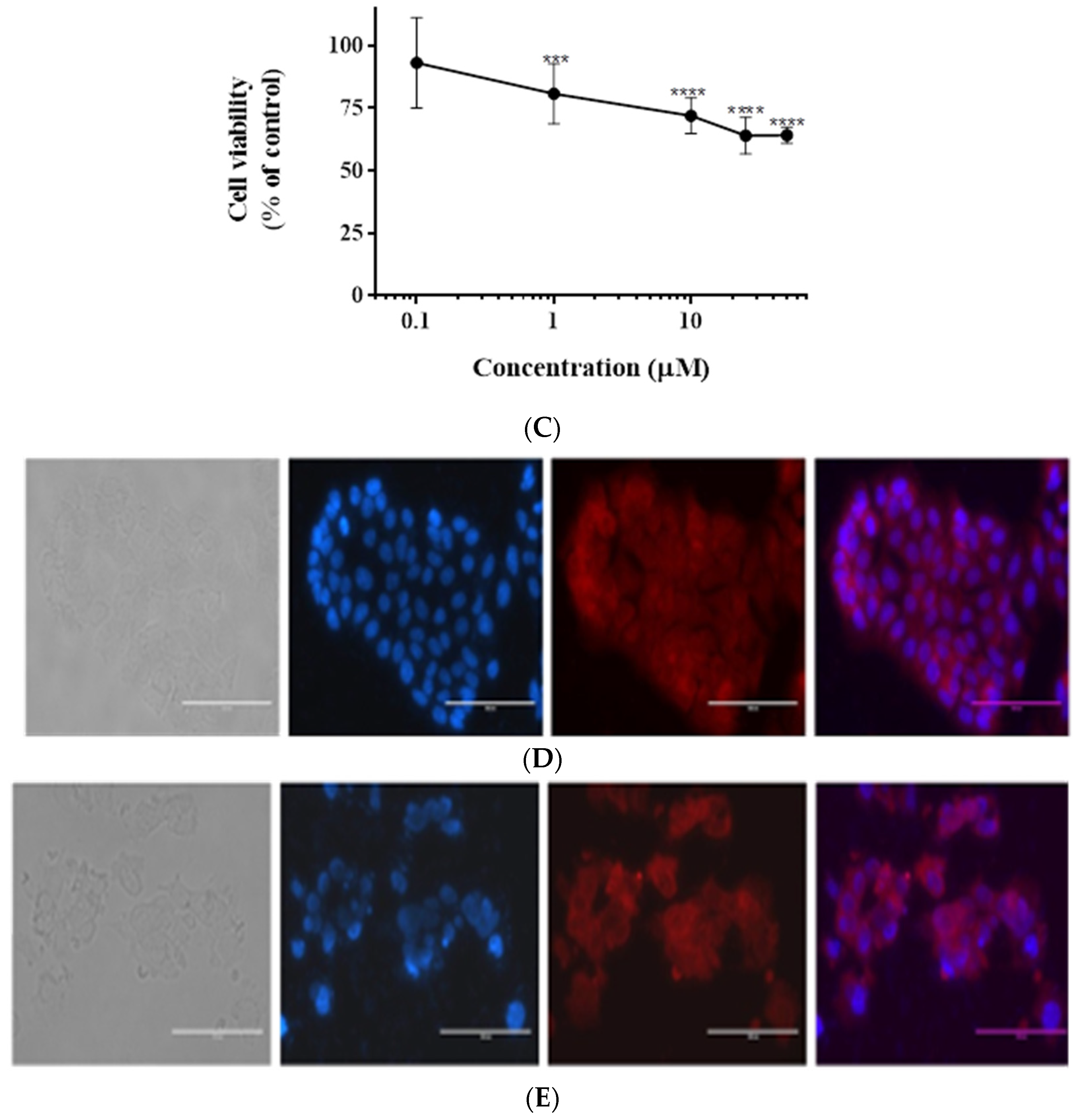
3.1. Impact of Casticin on the Viability of DLD-1, HCT116, and Caco-2 Cells
3.2. Casticin Treatment Impacts Colony-Forming Efficiency of DLD-1 Cells
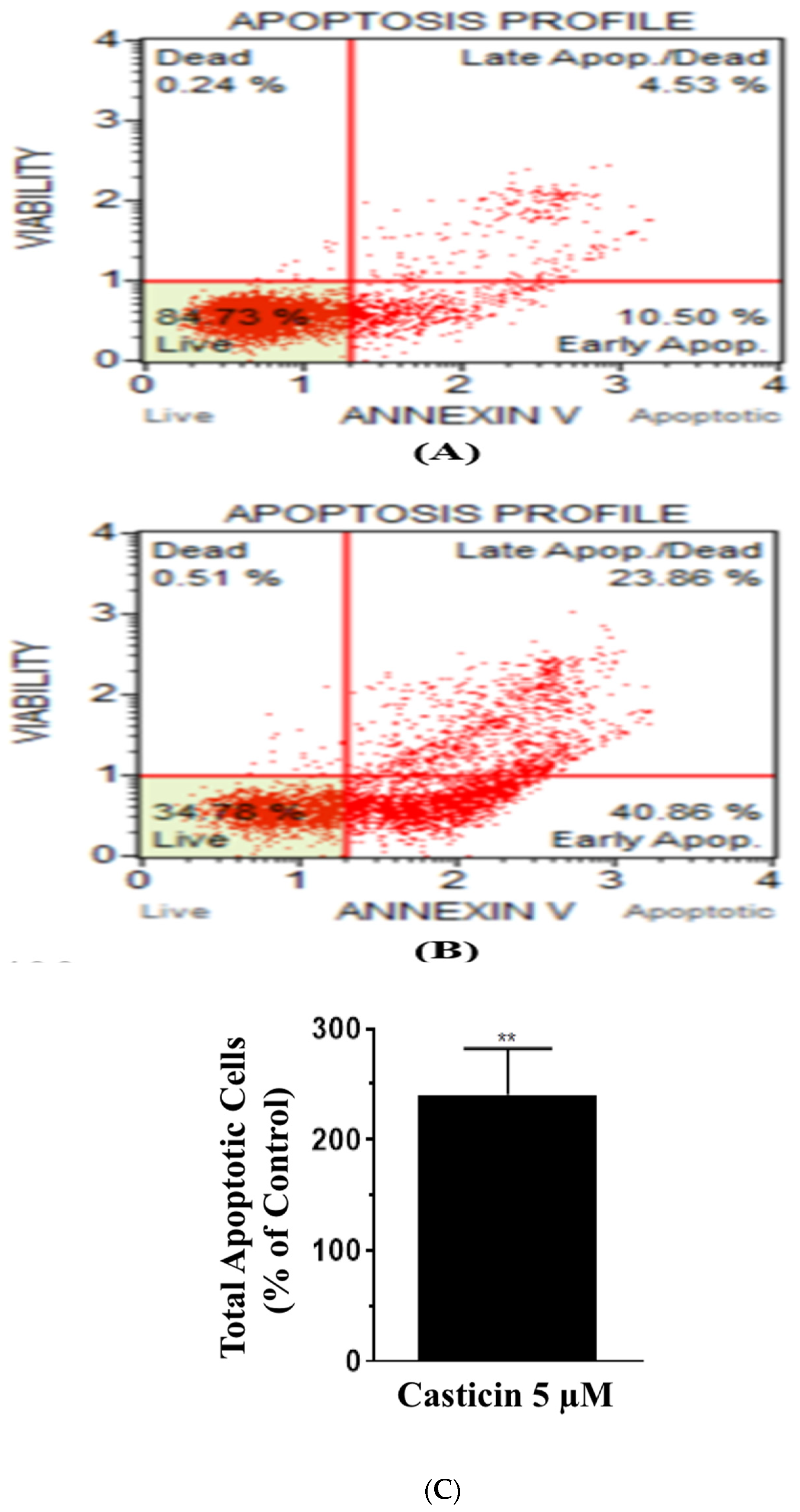
3.3. Casticin Invokes Extrinsic Programmed Cell Death Pathway
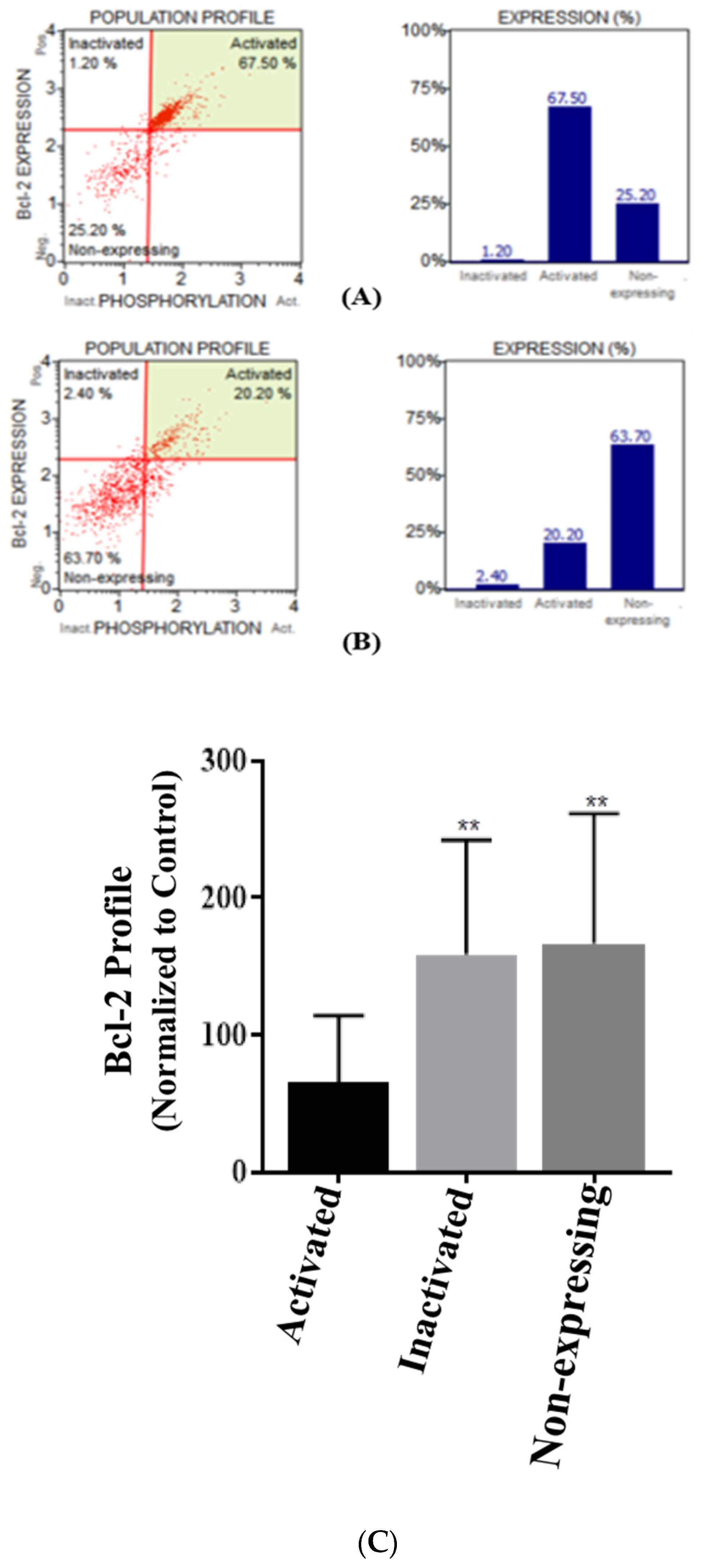
3.4. Casticin Impacts Bcl-2 Levels in DLD-1 Cells
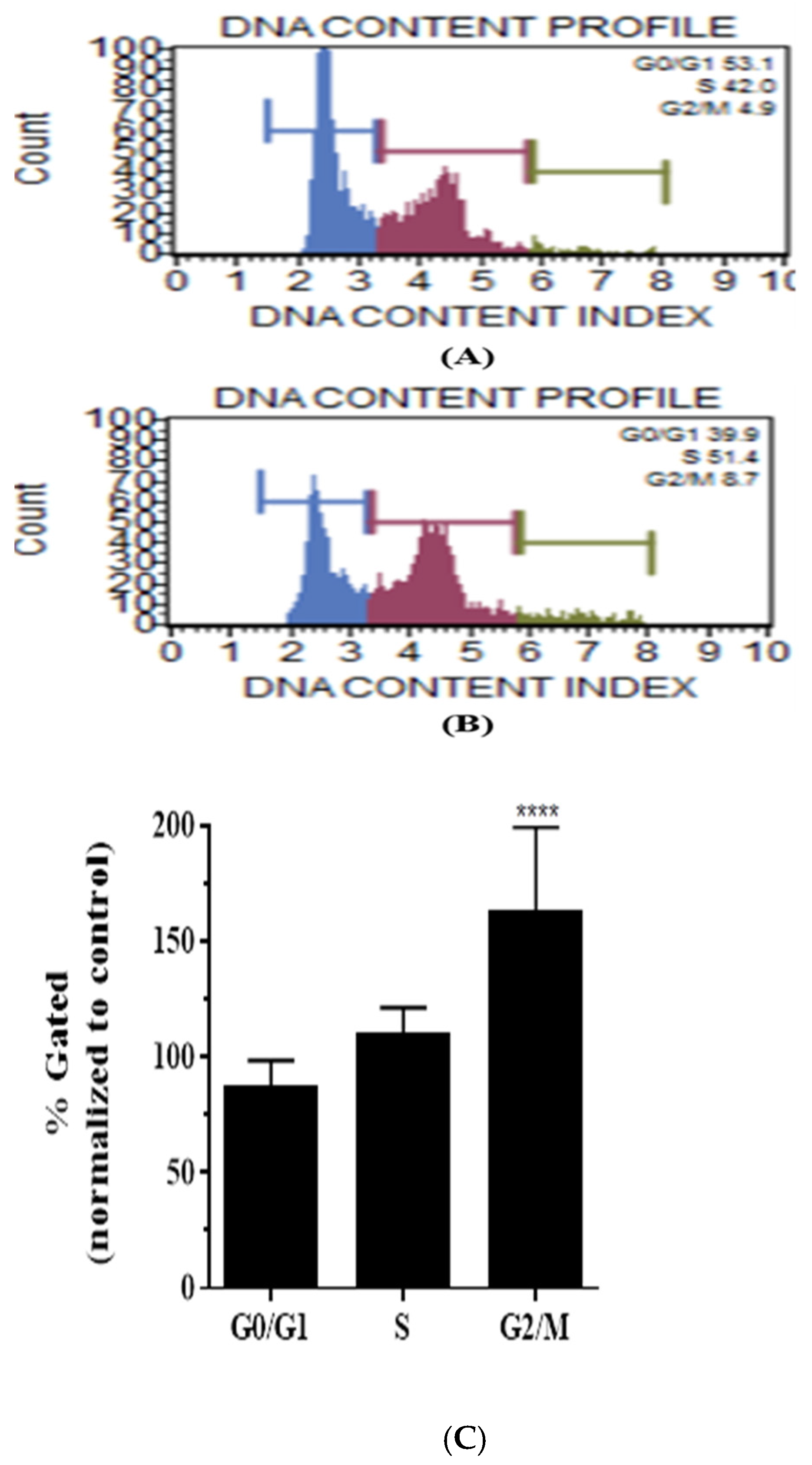
3.5. Casticin Induces Cell Cycle Arrest in DLD-1 Cells
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cancer Statistics Data Visualizations Tool. Available online: https://gis.cdc.gov/Cancer/USCS/DataViz.html (accessed on 8 March 2021).
- André, T.; Boni, C.; Mounedji-Boudiaf, L.; Navarro, M.; Tabernero, J.; Hickish, T.; Topham, C.; Zaninelli, M.; Clingan, P.; Bridgewater, J.; et al. Oxaliplatin, Fluorouracil, and Leucovorin as Adjuvant Treatment for Colon Cancer. N. Engl. J. Med. 2004, 350, 2343–2351. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morawska, K.; Goirand, F.; Marceau, L.; Devaux, M.; Cueff, A.; Bertaut, A.; Vincent, J.; Bengrine-Lefevre, L.; Ghiringhelli, F.; Schmitt, A. 5-FU therapeutic drug monitoring as a valuable option to reduce toxicity in patients with gastrointestinal cancer. Oncotarget 2018, 9, 11559–11571. [Google Scholar] [CrossRef] [Green Version]
- Blondy, S.; David, V.; Verdier, M.; Mathonnet, M.; Perraud, A.; Christou, N. 5-Fluorouracil resistance mechanisms in colorectal cancer: From classical pathways to promising processes. Cancer Sci. 2020, 111, 3142–3154. [Google Scholar] [CrossRef] [PubMed]
- Millimouno, F.M.; Dong, J.; Yang, L.; Li, J.; Li, X. Targeting apoptosis pathways in cancer and perspectives with natural compounds from mother nature. Cancer Prev. Res. 2014, 7, 1081–1107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bugel, S.M.; Tanguay, R.L. Multidimensional chemobehavior analysis of flavonoids and neuroactive compounds in zebrafish. Toxicol. Appl. Pharmacol. 2018, 344, 23–34. [Google Scholar] [CrossRef]
- Duan, C.; Matsumura, S.; Kariya, N.; Nishimura, M.; Shimono, T. In vitro antibacterial activities of Scutellaria baicalensis Georgi against cariogenic bacterial. Pediatric Dent. J. 2007, 17, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Afaq, F.; Mukhtar, H. Selective growth-inhibitory, cell-cycle deregulatory and apoptotic response of apigenin in normal versus human prostate carcinoma cells. Biochem. Biophys. Res. Commun. 2001, 287, 914–920. [Google Scholar] [CrossRef]
- Tang, S.Y.; Zhong, M.Z.; Yuan, G.J.; Hou, S.P.; Yin, L.L.; Jiang, H.; Yu, Z. Casticin, a flavonoid, potentiates TRAIL-induced apoptosis through modulation of anti-apoptotic proteins and death receptor 5 in colon cancer cells. Oncol. Rep. 2013, 29, 474–480. [Google Scholar] [CrossRef] [Green Version]
- He, L.; Yang, X.; Cao, X.; Liu, F.; Quan, M.; Cao, J. Casticin induces growth suppression and cell cycle arrest through activation of FOXO3a in hepatocellular carcinoma. Oncol. Rep. 2013, 29, 103–108. [Google Scholar] [CrossRef]
- Hussain, I.; Waheed, S.; Ahmad, K.A.; Pirog, J.E.; Syed, V. Scutellaria baicalensis targets the hypoxia-inducible factor-1α and enhances cisplatin efficacy in ovarian cancer. J. Cell. Biochem. 2018, 119, 7515–7524. [Google Scholar] [CrossRef]
- Liu, L.P.; Cao, X.C.; Liu, F.; Quan, M.F.; Sheng, X.F.; Ren, K.Q. Casticin induces breast cancer cell apoptosis by inhibiting the expression of forkhead box protein M1. Oncol. Lett. 2014, 7, 1711–1717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Razak, S.; Afsar, T.; Ullah, A.; Almajwal, A.; Alkholief, M.; Alshamsan, A.; Jahan, S. Taxifolin, a natural flavonoid interacts with cell cycle regulators causes cell cycle arrest and causes tumor regression by activating Wnt/β-catenin signaling pathway. BMC Cancer 2018, 18, 1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, X.-L.; Zhang, Y.-J.; Wang, X.-F.; Zhang, W.-J.; Wang, Z.; Zhang, F.; Zhang, Y.-J.; Lu, J.-H.; Mei, J.-W.; Hu, Y.-P.; et al. Casticin induces apoptosis and G0/G1 cell cycle arrest in gallbladder cancer cells. Cancer Cell Int. 2017, 17, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, C.Z.; Calway, T.D.; Wen, X.D.; Smith, J.; Yu, C.; Wang, Y.; Mehendale, S.R.; Yuan, C.S. Hydrophobic flavonoids from Scutellaria baicalensis induce colorectal cancer cell apoptosis through a mitochondrial-mediated pathway. Int. J. Oncol. 2013, 42, 1018–1026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dhir, H.; Choudhury, M.; Patil, K.; Cheung, C.; Bodlak, A.; Pardo, D.; Adams, A.; Travaglino, S.; Rojas, J.A.; Pai, S.B. Interception of Signaling Circuits of Esophageal Adenocarcinoma Cells by Resveratrol Reveals Molecular and Immunomodulatory Signatures. Cancers 2021, 13, 5811. [Google Scholar] [CrossRef]
- Lodise, O.; Patil, K.; Karshenboym, I.; Prombo, S.; Chukwueke, C.; Pai, S.B. Inhibition of Prostate Cancer Cells by 4,5-Dicaffeoylquinic Acid through Cell Cycle Arrest. Prostate Cancer 2019, 2019, 4520645. [Google Scholar] [CrossRef] [Green Version]
- Mitchel, J.; Bajaj, P.; Patil, K.; Gunnarson, A.; Pourchet, E.; Kim, Y.N.; Skolnick, J.; Pai, S.B. Computational Identification of Stearic Acid as a Potential PDK1 Inhibitor and In Vitro Validation of Stearic Acid as Colon Cancer Therapeutic in Combination with 5-Fluorouracil. Cancer Inf. 2021, 20, 11769351211065979. [Google Scholar] [CrossRef]
- Jiang, H.; Patil, K.; Vashi, A.; Wang, Y.; Strickland, E.; Pai, S.B. Cellular molecular and proteomic profiling deciphers the SIRT1 controlled cell death pathways in esophageal adenocarcinoma cells. Cancer Treat Res. Commun. 2021, 26, 100271. [Google Scholar] [CrossRef]
- Qu, L.; Liu, F.X.; Cao, X.C.; Xiao, Q.; Yang, X.; Ren, K.Q. Activation of the apoptosis signal-regulating kinase 1/c-Jun N-terminal kinase pathway is involved in the casticin-induced apoptosis of colon cancer cells. Exp. Ther. Med. 2014, 8, 1494–1500. [Google Scholar] [CrossRef]
- Li, W.X.; Cui, C.B.; Cai, B.; Wang, H.Y.; Yao, X.S. Flavonoids from Vitex trifolia L. inhibit cell cycle progression at G2/M phase and induce apoptosis in mammalian cancer cells. J. Asian Nat. Prod. Res. 2005, 7, 615–626. [Google Scholar] [CrossRef]
- Tesauro, C.; Keller, J.G.; Gromova, I.; Gromov, P.; Frøhlich, R.; Erlandsen, J.U.; Andersen, A.H.; Stougaard, M.; Knudsen, B.R. Different camptothecin sensitivities in subpopulations of colon cancer cells correlate with expression of different phospho-isoforms of topoisomerase i with different activities. Cancers 2020, 12, 1240. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kowalski, M.; Assa, A.; Patil, K.; Terrell, C.; Holliday, N.; Pai, S.B. Casticin Impacts Key Signaling Pathways in Colorectal Cancer Cells Leading to Cell Death with Therapeutic Implications. Genes 2022, 13, 815. https://doi.org/10.3390/genes13050815
Kowalski M, Assa A, Patil K, Terrell C, Holliday N, Pai SB. Casticin Impacts Key Signaling Pathways in Colorectal Cancer Cells Leading to Cell Death with Therapeutic Implications. Genes. 2022; 13(5):815. https://doi.org/10.3390/genes13050815
Chicago/Turabian StyleKowalski, Michael, Ashley Assa, Ketki Patil, Courtney Terrell, Nathan Holliday, and S. Balakrishna Pai. 2022. "Casticin Impacts Key Signaling Pathways in Colorectal Cancer Cells Leading to Cell Death with Therapeutic Implications" Genes 13, no. 5: 815. https://doi.org/10.3390/genes13050815
APA StyleKowalski, M., Assa, A., Patil, K., Terrell, C., Holliday, N., & Pai, S. B. (2022). Casticin Impacts Key Signaling Pathways in Colorectal Cancer Cells Leading to Cell Death with Therapeutic Implications. Genes, 13(5), 815. https://doi.org/10.3390/genes13050815