
Cats with Genetic Variants of AGXT2 Respond Differently to a Dietary Intervention Known to Reduce the Risk of Calcium Oxalate Stone Formation


Abstract
:1. Introduction
2. Materials and Methods
2.1. Cat Colony and Feeding Study Participants
2.2. Personalized Nutrition Study Design
2.3. Foods
2.4. Serum and Urine Analyses
2.5. Serum Metabolomics for Feeding Study Cats
2.6. Statistical Methods
3. Results
3.1. Genome Wide Association Study of Colony Cats
3.2. The Effect of Dietary Treatment on Food Intake and Body Weights
3.3. The Effect of Dietary Treatment on CBC and Selected Serum Chemistries
3.4. The Effect of Dietary Treatment on Urinalysis Parameters
3.5. The Effect of Dietary Treatment on Serum Metabolites
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ordovas, J.M.; Ferguson, L.R.; Tai, E.S.; Mathers, J.C. Personalised nutrition and health. BMJ 2018, 361, bmj.k2173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peneş, N.O.; Weber, B.; Păun, S.D. Role of genetic polymorphism in nutritional supplementation therapy in personalized medicine. Rom. J. Morphol. Embryol. 2017, 58, 53–58. [Google Scholar] [PubMed]
- Bush, C.L.; Blumberg, J.B.; El-Sohemy, A.; Minich, D.M.; Ordovás, J.M.; Reed, D.G.; Behm, V.A.Y. Toward the Definition of Personalized Nutrition: A Proposal by The American Nutrition Association. J. Am. Coll. Nutr. 2019, 39, 5–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barreiro, L.B.; Laval, G.; Quach, H.; Patin, E.; Quintana-Murci, L. Natural selection has driven population differentiation in modern humans. Nat. Genet. 2008, 40, 340–345. [Google Scholar] [CrossRef]
- Hall, J.A.; Brockman, J.A.; Davidson, S.J.; MacLeay, J.M.; Jewell, D.E. Increased dietary long-chain polyunsaturated fatty acids alter serum fatty acid concentrations and lower risk of urine stone formation in cats. PLoS ONE 2017, 12, e0187133. [Google Scholar] [CrossRef] [Green Version]
- Lulich, J.P.; Kruger, J.M.; MacLeay, J.M.; Merrills, J.M.; Paetau-Robinson, I.; Albasan, H.; Osborne, C.A. Efficacy of two commercially available, low-magnesium, urine-acidifying dry foods for the dissolution of struvite uroliths in cats. J. Am. Veter Med. Assoc. 2013, 243, 1147–1153. [Google Scholar] [CrossRef]
- Salido, E.; Pey, A.L.; Rodriguez, R.; Lorenzo, V. Primary hyperoxalurias: Disorders of glyoxylate detoxification. Biochim. Biophys. Acta Mol. Basis Dis. 2012, 1822, 1453–1464. [Google Scholar] [CrossRef] [Green Version]
- Cochat, P.; Rumsby, G. Primary Hyperoxaluria. N. Engl. J. Med. 2013, 369, 649–658. [Google Scholar] [CrossRef]
- Hu, X.-L.; Li, M.-P.; Song, P.-Y.; Tang, J.; Chen, X.-P. AGXT2: An unnegligible aminotransferase in cardiovascular and urinary systems. J. Mol. Cell. Cardiol. 2017, 113, 33–38. [Google Scholar] [CrossRef]
- Kittel, A.; Müller, F.; Koenig, J.; Mieth, M.; Sticht, H.; Zolk, O.; Kralj, A.; Heinrich, M.; Fromm, M.F.; Maas, R. Alanine-glyoxylate aminotransferase 2 (AGXT2) Polymorphisms Have Considerable Impact on Methylarginine and beta-aminoisobutyrate Metabolism in Healthy Volunteers. PLoS ONE 2014, 9, e88544. [Google Scholar] [CrossRef]
- Jewell, D.E.; Panickar, K.S. Botanicals Reduce Circulating Concentrations of Cholesterol and Triglycerides and Work Synergistically with Arachidonic Acid to Reduce Inflammatory Cytokines in Cats. Front. Veter Sci. 2021, 8, 620447. [Google Scholar] [CrossRef]
- NRC. Guide for the Care and Use of Laboratory Animals; National Academy Press: Washington, DC, USA, 2011. [Google Scholar]
- Chang, C.C.; Chow, C.C.; Tellier, L.C.; Vattikuti, S.; Purcell, S.M.; Lee, J.J. Second-generation PLINK: Rising to the challenge of larger and richer datasets. Gigascience 2015, 4, s13742-015. [Google Scholar] [CrossRef]
- Purcell, S.; Neale, B.; Todd-Brown, K.; Thomas, L.; Ferreira, M.A.R.; Bender, D.; Maller, J.; Sklar, P.; de Bakker, P.I.W.; Daly, M.J.; et al. PLINK: A Tool Set for Whole-Genome Association and Population-Based Linkage Analyses. Am. J. Hum. Genet. 2007, 81, 559–575. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.A.; Melendez, L.D.; Jewell, D.E. Using Gross Energy Improves Metabolizable Energy Predictive Equations for Pet Foods Whereas Undigested Protein and Fiber Content Predict Stool Quality. PLoS ONE 2013, 8, e54405. [Google Scholar] [CrossRef] [Green Version]
- Hall, J.; Yerramilli, M.; Obare, E.; Jewell, D. Comparison of Serum Concentrations of Symmetric Dimethylarginine and Creatinine as Kidney Function Biomarkers in Cats with Chronic Kidney Disease. J. Veter Intern. Med. 2014, 28, 1676–1683. [Google Scholar] [CrossRef] [Green Version]
- Hesse, A.; Bongartz, D.; Heynck, H.; Berg, W. Measurement of urinary oxalic acid: A comparison of five methods. Clin. Biochem. 1996, 29, 467–472. [Google Scholar] [CrossRef]
- Werness, P.G.; Brown, C.M.; Smith, L.H.; Finlayson, B. Equil2: A Basic Computer Program for the Calculation of Urinary Saturation. J. Urol. 1985, 134, 1242–1244. [Google Scholar] [CrossRef]
- Robert, M.; Boularan, A.M.; Colette, C.; Averous, M.; Monnier, M. Urinary calcium oxalate saturation in ‘stone formers’ and normal subjects: An application of the EQUIL2program. Br. J. Urol. 1994, 73, 358–361. [Google Scholar] [CrossRef]
- Milošević, D.; Batinić, D.; Blau, N.; Konjevoda, P.; Štambuk, N.; Votava-Raić, A.; Barbarić, V.; Fumić, K.; Rumenjak, V.; Stavljenić-Rukavina, A.; et al. Determination of urine saturation with computer program EQUIL 2 as a method for estimation of the risk of urolithiasis. J. Chem. Inf. Comput. Sci. 1998, 38, 646–650. [Google Scholar] [CrossRef]
- Laube, N.; Labedzke, V.; Hergarten, S.; Hesse, A. Determination of Urinary Calcium-Oxalate Formation Risk with BONN-Risk-Index and EQUIL Applied to a Family. J. Chem. Inf. Comput. Sci. 2002, 42, 633–639. [Google Scholar] [CrossRef]
- Laube, N.; Schneider, A.; Hesse, A. A new approach to calculate the risk of calcium oxalate crystallization from unprepared native urine. Urol. Res. 2000, 28, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, J.P.; Laube, N. Why Does the Bonn Risk Index Discriminate Between Calcium Oxalate Stone Formers and Healthy Controls? J. Urol. 2006, 175, 766–770. [Google Scholar] [CrossRef]
- Hall, J.A.; Jackson, M.; Vondran, J.C.; Vanchina, M.A.; Jewell, D.E. Comparison of circulating metabolite concentrations in dogs and cats when allowed to freely choose macronutrient intake. Biol. Open 2018, 7, bio036228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.-L.; Zeng, W.-J.; Li, M.-P.; Yang, Y.-L.; Kuang, D.-B.; Li, H.; Zhang, Y.-J.; Jiang, C.; Peng, L.-M.; Qi, H.; et al. AGXT2 rs37369 polymorphism predicts the renal function in patients with chronic heart failure. Gene 2017, 637, 145–151. [Google Scholar] [CrossRef]
- Jarzebska, N.; Georgi, S.; Jabs, N.; Brilloff, S.; Maas, R.; Rodionov, R.N.; Zietz, C.; Montresor, S.; Hohenstein, B.; Weiss, N. Kidney and liver are the main organs of expression of a key metabolic enzyme alanine:glyoxylate aminotransferase 2 in humans. Atheroscler. Suppl. 2019, 40, 106–112. [Google Scholar] [CrossRef]
- Rodionov, R.N.; Jarzebska, N.; Weiss, N.; Lentz, S.R. AGXT2: A promiscuous aminotransferase. Trends Pharmacol. Sci. 2014, 35, 575–582. [Google Scholar] [CrossRef] [Green Version]
- Suhre, K.; Wallaschofski, H.; Raffler, J.; Friedrich, N.; Haring, R.; Michael, K.; Wasner, C.; Krebs, A.; Kronenberg, F.; Chang, D.; et al. A genome-wide association study of metabolic traits in human urine. Nat. Genet. 2011, 43, 565–569. [Google Scholar] [CrossRef]
- Rhee, E.P.; Ho, J.; Chen, M.-H.; Shen, D.; Cheng, S.; Larson, M.; Ghorbani, A.; Shi, X.; Helenius, I.T.; O’Donnell, C.J.; et al. A Genome-wide Association Study of the Human Metabolome in a Community-Based Cohort. Cell Metab. 2013, 18, 130–143. [Google Scholar] [CrossRef] [Green Version]
- Caplin, B.; Wang, Z.; Slaviero, A.; Tomlinson, J.; Dowsett, L.; Delahaye, M.; Salama, A.; The International Consortium for Blood Pressure Genome-Wide Association Studies; Wheeler, D.C.; Leiper, J. Alanine-Glyoxylate Aminotransferase-2 Metabolizes Endogenous Methylarginines, Regulates NO, and Controls Blood Pressure. Arter. Thromb. Vasc. Biol. 2012, 32, 2892–2900. [Google Scholar] [CrossRef] [Green Version]
- Kittel, A.; Maas, R.; König, J.; Mieth, M.; Weiss, N.; Jarzebska, N.; Hohenstein, B.; Martens-Lobenhoffer, J.; Bode-Böger, S.M.; Rodionov, R.N. In vivo evidence that Agxt2 can regulate plasma levels of dimethylarginines in mice. Biochem. Biophys. Res. Commun. 2013, 430, 84–89. [Google Scholar] [CrossRef]
- Turner, M.A.; Yang, X.; Yin, D.; Kuczera, K.; Borchardt, R.T.; Howell, P.L. Structure and Function of S-Adenosylhomocysteine Hydrolase. Cell Biophys. 2000, 33, 101–125. [Google Scholar] [CrossRef]
- Papasotiriou, M.; Genovese, F.; Klinkhammer, B.M.; Kunter, U.; Nielsen, S.H.; Karsdal, M.A.; Floege, J.; Boor, P. Serum and urine markers of collagen degradation reflect renal fibrosis in experimental kidney diseases. Nephrol. Dial. Transplant. 2015, 30, 1112–1121. [Google Scholar] [CrossRef] [Green Version]
- Vangaveti, V.; Baune, B.T.; Kennedy, R.L. Hydroxyoctadecadienoic acids: Novel regulators of macrophage differentiation and atherogenesis. Ther. Adv. Endocrinol. Metab. 2010, 1, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Wolkow, A.; Aisbett, B.; Reynolds, J.; Ferguson, S.A.; Main, L.C. Relationships between inflammatory cytokine and cortisol responses in firefighters exposed to simulated wildfire suppression work and sleep restriction. Physiol. Rep. 2015, 3, e12604. [Google Scholar] [CrossRef] [Green Version]
- Jia, Y.; Liu, L.; Sheng, C.; Cheng, Z.; Cui, L.; Li, M.; Zhao, Y.; Shi, T.; Yau, T.O.; Li, F.; et al. Increased Serum Levels of Cortisol and Inflammatory Cytokines in People with Depression. J. Nerv. Ment. Dis. 2019, 207, 271–276. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Nutrient 3 | Pre-Trial & Control Food | Test Food |
|---|---|---|
| Moisture | 5.47 | 5.81 |
| Protein | 32.9 | 33.0 |
| Fat | 14.6 | 14.5 |
| Atwater Energy, 4 kcal/kg | 3815 | 3816 |
| Ash | 4.89 | 4.69 |
| Crude Fiber | 1.5 | 1.2 |
| Betaine (mg/kg) | 344 | 5210 |
| Calcium | 0.69 | 0.71 |
| Phosphorus | 0.80 | 0.83 |
| Sodium | 0.38 | 0.41 |
| Magnesium | 0.08 | 0.08 |
| ARA [20:4 (n-6)] | 0.09 | 0.10 |
| EPA [20:5 (n-3)] | 0.15 | 0.16 |
| DHA [22:6 (n-3)] | 0.11 | 0.12 |
| SFA 5 | 3.40 | 3.28 |
| MUFA 6 | 4.56 | 4.74 |
| PUFA 7 | 3.78 | 3.94 |
| Total FA | 11.62 | 12.07 |
| (n-6) FA 8 | 3.19 | 3.29 |
| (n-3) FA 9 | 0.51 | 0.56 |
| (n-6):(n-3) ratio | 6.2 | 5.9 |
| AA | AG | GG | |
|---|---|---|---|
| 2-oxoarginine | 0.45 ± 0.10 c | 0.92 ± 0.04 b | 1.27 ± 0.04 a |
| β-aminoisobutyrate | 6.53 ± 0.94 a | 3.05 ± 0.81 b | 1.53 ± 0.81 b |
| Dimethyl arginines(SDMA + ADMA) | 1.26 ± 0.05 a | 1.01 ± 0.05 b | 1.00 ± 0.05 b |
| Initial | Control Food | Test Food | |
|---|---|---|---|
| Body weight (kg) | 5.0 ± 0.7 | 5.0 ± 0.7 | 5.0 ± 0.8 |
| Food intake (grams) | 44.4 ± 17.6 | 47.4 ± 1.7 | 45.9 ± 1.7 |
| Serum Concentrations | AA | AG | GG |
|---|---|---|---|
| Creatinine (mg/dL) | |||
| Initial | 1.15 ± 0.06 | 1.21 ± 0.02 | 1.26 ± 0.06 |
| Control Food | 1.17 ± 0.06 | 1.18 ± 0.09 | 1.21 ± 0.06 |
| Test Food | 1.17 ± 0.06 | 1.15 ± 0.09 | 1.16 ± 0.06 |
| Urea (mg/dL) | |||
| Initial | 23.3 ± 1.3 | 19.9 ± 0.8 | 22.2 ± 1.5 |
| Control Food | 23.3 ± 1.0 | 19.9 ± 1.5 | 21.4 ± 1.0 |
| Test Food | 22.0 ± 1.0 | 20.8 ± 1.5 | 21.8 ± 1.0 |
| Albumin (mg/dL) | |||
| Initial | 3.20 ± 0.07 | 3.50 ± 0.13 | 3.58 ± 0.10 |
| Control Food | 3.38 ± 0.10 | 3.45 ± 0.14 | 3.59 ± 0.10 |
| Test Food | 3.25 ± 0.10 | 3.45 ± 0.14 | 3.46 ± 0.10 |
| Protein (mg/dL) | |||
| Initial | 6.77 ± 0.17 | 7.00 ± 0.25 | 6.84 ± 0.18 |
| Control Food | 7.14 ± 0.21 | 7.09 ± 0.30 | 7.01 ± 0.20 |
| Test Food | 7.03 ± 0.21 | 6.87 ± 0.30 | 6.99 ± 0.20 |
| Cholesterol (mg/dL) | |||
| Initial | 142 ± 16 | 172 ± 17 | 152 ± 18 |
| Control Food | 152 ± 13 | 188 ± 18 | 173 ± 13 |
| Test Food | 155 ± 13 | 200 ± 18 | 181 ± 13 |
| Triglycerides (mg/dL) | |||
| Initial | 32.8 ± 2.4 | 45.5 ± 10.0 | 37.6 ± 4.2 |
| Control Food | 36.7 a ± 3.6 | 43.3 ± 10.0 | 32.8 ± 4.2 |
| Test Food | 30.8 b,d ± 3.6 | 46.1 c ± 10.0 | 32.3 d ± 4.2 |
| Calcium (mg/dL) | |||
| Initial | 9.44 ± 0.14 | 9.82 ± 0.17 | 9.83 ± 0.12 |
| Control Food | 9.57 ± 0.13 | 9.65 ± 0.19 | 9.71 ± 0.12 |
| Test Food | 9.54 ± 0.13 | 9.57 ± 0.19 | 9.60 ± 0.12 |
| Serum Concentrations | AA | AG | GG |
|---|---|---|---|
| Interleuken 1-β (ln pg/mL) | 2.21 ± 0.27 b | 2.95 ± 0.41 a,b | 3.10 ± 0.17 a |
| Interleuken 2 (ln pg/mL) | 1.87 ± 0.35 a,b | 0.88 ± 0.53 b | 2.81 ± 0.34 a |
| Interleuken 4 (ln pg/mL) | 4.73 ± 0.73 b | 6.22 ± 0.51 a | 5.47 ± 0.33 a,b |
| Interleuken 6 (ln pg/mL) | 3.56 ± 0.26 b | 4.63 ± 0.40 a | 4.52 ± 0.26 a |
| Interleuken 8 (ln pg/mL) | 2.73 ± 0.18 | 3.23 ± 0.16 | 3.22 ± 0.17 |
| Interleuken 18 (ln pg/mL) | 4.57 ± 0.28 a,b | 3.62 ± 0.42 b | 4.97 ± 0.28 a |
| MCP-1 (ln pg/mL) | 6.91 ± 0.21 | 7.40 ± 0.32 | 7.23 ± 0.21 |
| Interleuken 13 (ln pg/mL) | 2.02 ± 0.27 b | 1.69 ± 0.41 b | 2.70 ± 0.17 a |
| Flt 3L (ln pg/mL) | 3.90 ± 0.17 | 3.98 ± 0.25 | 3.83 ± 0.16 |
| TNF-α (ln pg/mL) | 0.93 ± 0.47 b | 2.89 ± 0.70 a | 2.62 ± 0.46 a |
| KC 1 (ln pg/mL) | -0.91 ± 0.40 b | -0.62 ± 0.61 a,b | 0.53 ± 0.18 a |
| RANTES 2 (ln pg/mL) | 2.53 ± 0.17 b | 3.04 ± 0.2,6 a,b | 3.16 ± 0.17 a |
| Interferon-γ (ln pg/mL) | 3.09 ± 0.37 b | 3.86 ± 0.56 a,b | 4.13 ± 0.37 a |
| Stem cell factor (ln pg/mL) | 4.67 ± 0.12 b | 5.39 ± 0.18 a | 5.14 ± 0.12 a |
| Interleuken 12 p40 (ln pg/mL) | 5.62 ± 0.21 | 5.77 ± 0.31 | 5.65 ± 0.20 |
| PDGF 3 BB (ln pg/mL) | 4.05 ± 0.37 | 3.10 ± 0.55 | 4.47 ± 0.36 |
| SDF-1 4 (ln pg/mL) | 5.61 ± 0.34 | 6.64 ± 0.52 | 6.04 ± 0.34 |
| FAS 5 (ln pg/mL) | 3.57 ± 0.26 b | 4.63 ± 0.40 a | 4.52 ± 0.26 a |
| Urine Parameters | AA | AG | GG |
|---|---|---|---|
| Calcium (mg/dL) | |||
| Initial | 3.10 ± 1.27 | 3.22 ± 1.58 | 3.36 ± 0.99 |
| Control Food | 3.42 ± 0.39 | 3.16 ± 0.56 | 3.72 ± 0.38 |
| Test Food | 3.78 ± 0.39 | 3.13 ± 0.56 | 3.06 ± 0.39 |
| Creatinine (mg/dL) | |||
| Initial | 329 ± 80 | 414 ± 31 | 447 ± 74 |
| Control Food | 357 ± 27 | 388 ± 38 | 438 ± 28 |
| Test Food | 346 ± 27 | 369 ± 38 | 397 ± 27 |
| Protein to Creatinine ratio | |||
| Initial | 0.16 ± 0.02 | 0.14 ± 0.03 | 0.16 ± 0.02 |
| Control Food | 0.17 ± 0.02 | 0.15 ± 0.03 | 0.16 ± 0.02 |
| Test Food | 0.15 ± 0.02 | 0.13 ± 0.03 | 0.15± 0.02 |
| Oxalate (µM) | |||
| Initial | NA | NA | NA |
| Control Food | 1042 ± 101 | 1337 ± 143 | 1011 ± 95 |
| Test Food | 822 ± 101 | 877 ± 143 | 910 ± 94 |
| pH | |||
| Initial | 6.30 ± 0.12 | 6.30 ± 0.25 | 6.31 ± 0.09 |
| Control Food | 6.11 ± 0.14 | 5.93 ± 0.19 | 6.35 ± 0.13 |
| Test Food | 6.18 ± 0.14 | 6.16 ± 0.19 | 6.41 ± 0.13 |
| Specific gravity | |||
| Initial | 1.044 ± 0.011 | 1.045 ± 0.001 | 1.050 ± 0.003 |
| Control Food | 1.048 ± 0.003 | 1.049 ± 0.004 | 1.049 ± 0.002 |
| Test Food | 1.043 ± 0.003 | 1.046 ± 0.004 | 1.049 ± 0.002 |
| Fractional excretion of calcium (%) | |||
| Initial | 0.115 ± 0.046 | 0.091 ± 0.047 | 0.094 ± 0.038 |
| Control Food | 0.119 ± 0.017 | 0.100 ± 0.024 | 0.103 ± 0.018 |
| Test Food | 0.139 ± 0.017 | 0.102 ± 0.024 | 0.091 ± 0.017 |
| Struvite relative super saturation (ln) | |||
| Initial | −1.42 ± 0.40 | −0.46 ± 0.36 | −0.27 ± 0.40 |
| Control Food | −0.97 ± 0.38 | −1.03 ± 0.54 | −0.04 ± 0.38 |
| Test Food | −1.10 ± 0.38 | −0.96 ± 0.54 | 0.06 ± 0.38 |
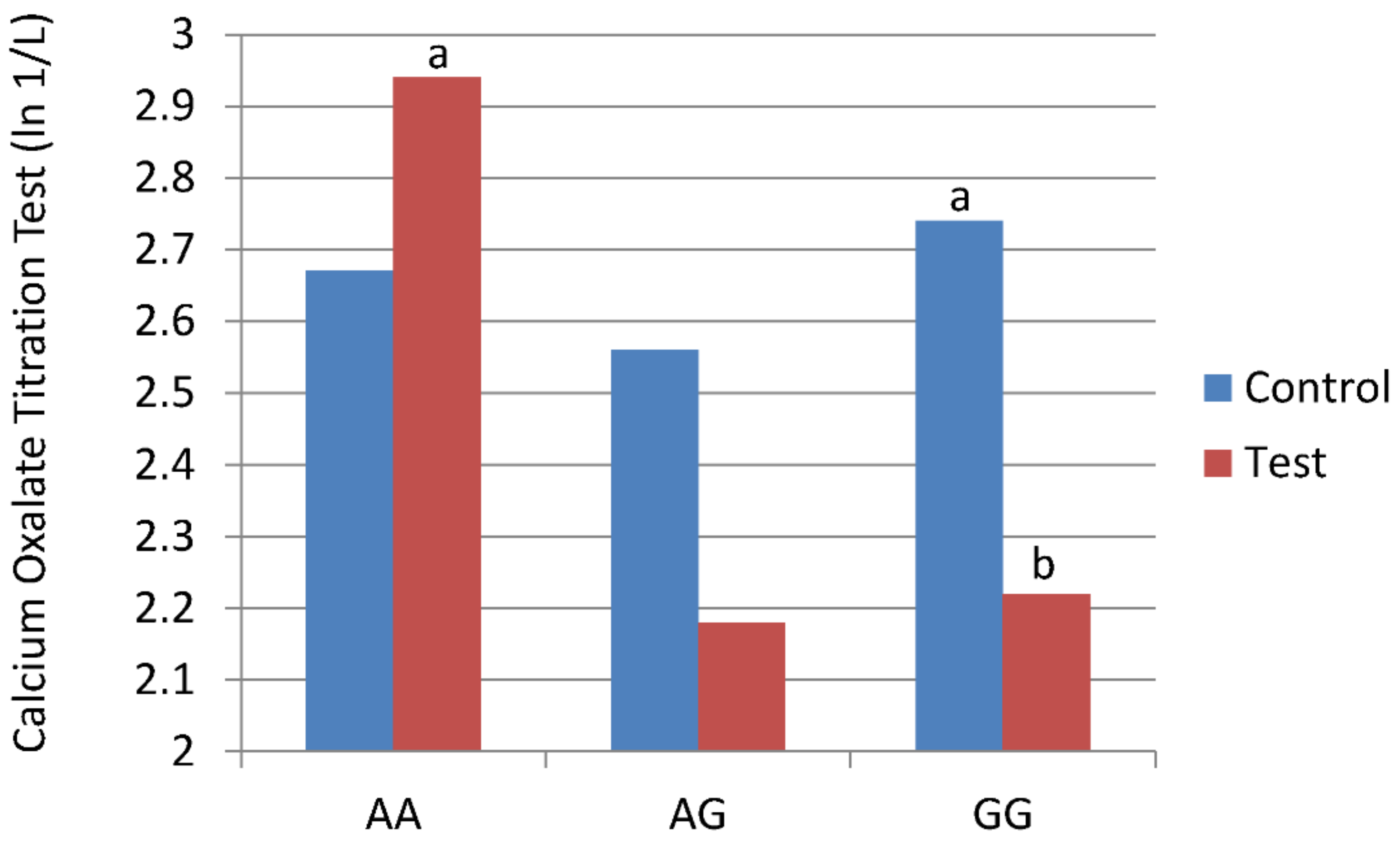
| Calcium oxalate titration test (ln 1/L) | |||
| Initial | 2.55 ± 0.25 | 2.15 ± 0.61 | 2.58 ± 0.21 |
| Control Food | 2.67 ± 0.29 | 2.56 ± 0.40 | 2.74 ± 0.28 a |
| Test Food | 2.94 ± 0.29 c | 2.18 ± 0.40 c,d | 2.12 ± 0.28 b,d |
| Metabolism Pathway | Biochemical Name | AA Test/Control | AG Test/Control | GG Test/Control |
|---|---|---|---|---|
| Glycine, Serine, Threonine Metabolism | sarcosine | 1.39 | 1.76 | 1.64 |
| betaine | 2.06 | 2.6 | 2.28 | |
| N-acetylthreonine | 0.85 | 1.08 | 0.85 | |
| Alanine and Asparagine Metabolism | N-methylalanine | 1.18 | 1.58 | 2.51 |
| N,N-dimethylalanine | 0.51 | 0.48 | 0.45 | |
| N-acetylasparagine | 0.85 | 1.02 | 0.81 | |
| Hydroxyasparagine | 0.85 | 0.96 | 0.81 | |
| Glutamine Metabolism | glutamine | 0.95 | 1.05 | 0.95 |
| α-ketoglutaramate | 1.2 | 1.67 | 1.26 | |
| Pyroglutamine | 1.3 | 1.83 | 1.28 | |
| N-acetyl-aspartyl-glutamate (NAAG) | 0.89 | 1.42 | 0.9 | |
| Histidine Metabolism | imidazole propionate | 1.21 | 1.88 | 1.29 |
| carnosine | 0.94 | 1.2 | 0.91 | |
| N-acetylcarnosine | 0.91 | 1.2 | 0.9 | |
| Lysine Metabolism | N6,N6-dimethyllysine | 0.95 | 1.15 | 0.94 |
| N6,N6,N6-trimethyllysine | 0.95 | 1.13 | 0.91 | |
| hydroxy-N6,N6,N6-trimethyllysine | 0.79 | 1.29 | 0.7 | |
| 5-(galactosylhydroxy)-L-lysine | 0.88 | 1.7 | 0.73 | |
| glutarylcarnitine (C5-DC) | 0.83 | 1.15 | 0.71 | |
| Tryptophan Metabolism | C-glycosyltryptophan | 0.84 | 1.24 | 0.89 |
| Leucine, Isoleucine, Valine Metabolism | α-hydroxyisocaproate | 0.88 | 0.96 | 0.88 |
| 3-methylglutaconate | 0.96 | 1.22 | 0.88 | |
| 2-methylbutyrylcarnitine (C5) | 1.04 | 1.4 | 0.8 | |
| tiglylcarnitine (C5:1-DC) | 0.8 | 0.88 | 0.8 | |
| α-hydroxyisovalerate | 0.91 | 1.09 | 0.85 | |
| isobutyrylcarnitine (C4) | 0.95 | 1.17 | 0.82 | |
| Methionine, Cysteine, Metabolism | methionine | 1.07 | 1.26 | 1.12 |
| N-acetylmethionine | 0.83 | 1.16 | 0.87 | |
| N-formylmethionine | 0.91 | 1.17 | 0.89 | |
| methionine sulfoxide | 1.06 | 1.24 | 1.17 | |
| Proline Metabolism | trans-4-hydroxyproline | 0.68 | 0.63 | 0.65 |
| N-methylproline | 0.93 | 0.97 | 0.72 | |
| Creatine Metabolism | creatinine | 0.98 | 1.05 | 0.95 |
| Polyamine Metabolism | N-acetyl-isoputreanine | 0.92 | 1.11 | 0.91 |
| Glutathione Metabolism | 2-hydroxybutyrate/2-hydroxyisobutyrate | 0.84 | 0.86 | 0.8 |
| γ-glutamyl Amino Acid Metabolism | γ-glutamyl-epsilon-lysine | 0.9 | 1.19 | 0.76 |
| Dipeptide Metabolism | leucylhydroxyproline | 1.27 | 1.52 | 1.68 |
| Energy Metabolism | malate | 0.84 | 0.98 | 0.87 |
| Fatty Acid Metabolism | malonylcarnitine | 0.9 | 0.89 | 0.58 |
| malonate | 1.51 | 1.2 | 0.85 | |
| heptanoate (7:0) | 0.84 | 0.91 | 0.75 | |
| Long Chain Polyunsaturated Fatty Acid (n3 and n6) Metabolism | tetradecadienoate (14:2) | 0.92 | 0.94 | 0.74 |
| hexadecatrienoate (16:3n3) | 0.78 | 1.06 | 0.86 | |
| (11 or 12)-methyltridecanoate (a14:0 or i14:0) | 0.8 | 0.91 | 0.9 | |
| Fatty Acid, Dicarboxylate | glutarate (C5-DC) | 0.92 | 0.78 | 0.81 |
| dodecadienoate (12:2) | 0.96 | 1 | 0.76 | |
| branched chain 14:0 dicarboxylic acid | 0.95 | 1.65 | 0.95 | |
| hexadecanedioate (C16-DC) | 0.75 | 1.22 | 0.85 | |
| hexadecenedioate (C16:1-DC) | 0.83 | 1.38 | 0.96 | |
| heptadecanedioate (C17-DC) | 0.74 | 1.24 | 0.82 | |
| octadecanedioate (C18-DC) | 0.83 | 1.27 | 0.84 | |
| nonadecanedioate (C19-DC) | 0.88 | 1.32 | 0.79 | |
| eicosanedioate (C20-DC) | 0.89 | 1.36 | 0.84 | |
| docosadioate (C22-DC) | 0.75 | 1.35 | 0.83 | |
| butyrylglycine | 0.66 | 0.79 | 0.66 | |
| Fatty Acid Metabolism (Acyl Glycine) | hexanoylglycine | 0.67 | 0.66 | 0.53 |
| 3,4-methylene heptanoylglycine | 0.7 | 0.55 | 0.76 | |
| N-octanoylglycine | 0.65 | 0.92 | 0.54 | |
| lignoceroylcarnitine (C24) | 0.82 | 0.89 | 0.81 | |
| cerotoylcarnitine (C26) | 0.8 | 0.91 | 0.62 | |
| Fatty Acid Acyl Carnitine Metabolism | nervonoylcarnitine (C24:1) | 0.82 | 1.29 | 0.75 |
| ximenoylcarnitine (C26:1) | 0.72 | 0.76 | 0.61 | |
| adrenoylcarnitine (C22:4) | 0.78 | 1.3 | 0.72 | |
| docosahexaenoylcarnitine (C22:6) | 0.93 | 1.44 | 0.87 | |
| Fatty Acyl Carnitine dicarboxylate Metabolism | adipoylcarnitine (C6-DC) | 0.8 | 1.05 | 0.81 |
| suberoylcarnitine (C8-DC) | 0.68 | 0.52 | 0.69 | |
| Fatty Acid Acyl Carnitine hydroxy Metabolism | (R)-3-hydroxybutyrylcarnitine | 1.03 | 1.68 | 0.38 |
| 3-hydroxyoleoylcarnitine | 0.89 | 1.23 | 0.81 | |
| Fatty Acid Mono hydroxyl Metabolism | 2-hydroxyoctanoate | 0.87 | 1.13 | 0.72 |
| 2-hydroxylaurate | 0.85 | 1.07 | 0.83 | |
| 3-hydroxyoctanoate | 1.11 | 0.86 | 0.75 | |
| 3-hydroxydecanoate | 1 | 0.99 | 0.78 | |
| 3-hydroxysebacate | 0.89 | 1 | 0.76 | |
| 16-hydroxypalmitate | 0.85 | 1.12 | 0.85 | |
| Endocannabinoid Metabolism | hexanoyltaurine | 0.83 | 1.14 | 0.75 |
| Lysophospholipid Metabolism | 1-linoleoyl-GPA (18:2) | 0.78 | 0.87 | 0.67 |
| 1-palmitoyl-GPE (16:0) | 0.84 | 1.26 | 0.9 | |
| 1-stearoyl-GPE (18:0) | 0.86 | 1.4 | 0.91 | |
| 1-arachidonoyl-GPE (20:4n6) | 0.83 | 1.41 | 0.96 | |
| 1-(1-enyl-stearoyl)-2-oleoyl-GPE (P-18:0/18:1) | 0.76 | 1.13 | 0.95 | |
| Diacylglycerol Metabolism | oleoyl-linoleoyl-glycerol (18:1/18:2) | 0.63 | 1.52 | 0.93 |
| Mevalonate Metabolism | 3-hydroxy-3-methylglutarate | 0.93 | 1.13 | 0.92 |
| Pyrimidine Metabolism | 3-ureidopropionate | 0.95 | 1.06 | 0.93 |
| 3-(3-amino-3-carboxypropyl) uridine | 0.79 | 1.17 | 0.78 | |
| 5-methyl-2’-deoxycytidine | 1 | 1.2 | 1.15 | |
| nicotinamide | 0.89 | 1.19 | 0.9 | |
| gulonate | 0.77 | 1.1 | 0.92 | |
| 4-allylcatechol sulfate | 2.86 | 2.92 | 3.58 | |
| Xanthine Metabolism | theophylline | 44.08 | 42.57 | 35.37 |
| Food Component | ergothioneine | 1.18 | 1.49 | 1.25 |
| methyl indole-3-acetate | 0.8 | 2.07 | 0.9 | |
| eugenol sulfate | 28.82 | 21.9 | 33.99 | |
| (2,4 or 2,5)-dimethylphenol sulfate | 0.71 | 0.73 | 0.6 | |
| Chemical | sulfate | 0.9 | 1.05 | 0.93 |
| 1,2,3-benzenetriol sulfate | 2.23 | 15.29 | 11.31 | |
| Partially Characterized Molecules | glycine conjugate of C10H14O2 | 0.72 | 0.89 | 0.8 |
| Serum Concentrations | AA | AG | GG |
|---|---|---|---|
| 2-oxoarginine 1 | |||
| Initial | 0.65 ± 0.30 | 1.82 ± 0.45 | 2.21 ± 0.28 |
| Control Food | 0.78 ± 0.30 | 1.79 ± 0.46 | 2.12 ± 0.29 |
| Test Food | 0.76 ± 0.30 | 2.10 ± 0.46 | 2.11 ± 0.29 |
| β-aminoisobutyrate 1 | |||
| Initial | 1.67 ± 0.28 | 0.57 ± 0.42 | 0.63 ± 0.26 |
| Control Food | 1.78 ± 0.29 | 0.55 ± 0.44 | 0.60 ± 0.28 |
| Test Food | 2.21 ± 0.29 | 0.62 ± 0.44 | 0.78 ± 0.28 |
| Oxalate | |||
| Initial | 1.06 ± 0.12 | 1.13 ± 0.18 | 1.11 ± 0.11 |
| Control Food | 1.01 ± 0.11 | 1.02 ± 0.18 | 1.04 ± 0.11 |
| Test Food | 0.94 ± 0.12 | 1.01 ± 0.18 | 0.95 ± 0.11 |
| Dimethyl arginines (SDMA + ADMA) | |||
| Initial | 1.07 ± 0.06 | 0.96 ± 0.10 | 0.96 ± 0.06 |
| Control Food | 1.11 ± 0.06 | 0.94 ± 0.08 | 0.99 ± 0.05 |
| Test Food | 1.06 ± 0.06 | 1.02 ± 0.08 | 0.94 ± 0.05 |
| S-adenosylhomocysteine 1 | |||
| Initial | 1.01 ± 0.08 | 0.93 ± 0.13 | 1.11 ± 0.08 |
| Control Food | 0.97 ± 0.17 | 0.70 ± 0.25 b | 1.19 ± 0.16 b |
| Test Food | 1.04 ± 0.17 | 0.99 ± 0.25 a | 1.75 ± 0.16 a |
| Oxidized glutathione 2 | |||
| Initial | 0.84 ± 0.12 | 0.99 ± 0.19 | 1.18 ± 0.12 |
| Control Food | 0.97 ± 0.18 | 0.91 ± 0.27 | 1.45 ± 0.17 a |
| Test Food | 0.95 ± 0.18 | 1.25 ± 0.27 | 1.13 ± 0.17 b |
| Cortisol 1 | |||
| Initial | 0.97 ± 0.30 | 1.37 ± 0.44 | 1.69 ± 0.28 |
| Control Food | 0.94 ± 0.26 | 1.13 ± 0.39 | 1.80 ± 0.24 |
| Test Food | 0.74 ± 0.26 | 0.85 ± 0.39 | 1.27 ± 0.24 |
| 9-HODE + 13-HODE 1 | |||
| Initial | 1.60 ± 0.16 | 1.01 ± 0.24 | 1.13 ± 0.15 |
| Control Food | 1.56 ± 0.14 | 1.02 ± 0.21 | 1.02 ± 0.13 |
| Test Food | 1.20 ± 0.14 | 0.73 ± 0.21 | 0.84 ± 0.13 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hall, J.A.; Panickar, K.S.; Brockman, J.A.; Jewell, D.E. Cats with Genetic Variants of AGXT2 Respond Differently to a Dietary Intervention Known to Reduce the Risk of Calcium Oxalate Stone Formation. Genes 2022, 13, 791. https://doi.org/10.3390/genes13050791
Hall JA, Panickar KS, Brockman JA, Jewell DE. Cats with Genetic Variants of AGXT2 Respond Differently to a Dietary Intervention Known to Reduce the Risk of Calcium Oxalate Stone Formation. Genes. 2022; 13(5):791. https://doi.org/10.3390/genes13050791
Chicago/Turabian StyleHall, Jean A., Kiran S. Panickar, Jeffrey A. Brockman, and Dennis E. Jewell. 2022. "Cats with Genetic Variants of AGXT2 Respond Differently to a Dietary Intervention Known to Reduce the Risk of Calcium Oxalate Stone Formation" Genes 13, no. 5: 791. https://doi.org/10.3390/genes13050791
APA StyleHall, J. A., Panickar, K. S., Brockman, J. A., & Jewell, D. E. (2022). Cats with Genetic Variants of AGXT2 Respond Differently to a Dietary Intervention Known to Reduce the Risk of Calcium Oxalate Stone Formation. Genes, 13(5), 791. https://doi.org/10.3390/genes13050791