Abstract
The purpose of this study was to compare the frequency of COL1A1 rs1107946 polymorphism between sport climbers and controls from three ethnic groups (Japanese, Polish, and Russian) and investigate the effect of the COL1A1 rs1107946 polymorphism on the age-related decrease in flexibility in the general population. Study I consisted of 1929 healthy people (controls) and 218 climbers, including Japanese, Polish, and Russian participants. The results of the meta-analysis showed that the frequency of the AC genotype was higher in climbers than in the controls (p = 0.03). Study II involved 1093 healthy Japanese individuals (435 men and 658 women). Flexibility was assessed using a sit-and-reach test. There was a tendency towards association between sit-and-reach and the COL1A1 rs1107946 polymorphism (genotype: p = 0.034; dominant: p = 0.435; recessive: p = 0.035; over-dominant: p = 0.026). In addition, there was a higher negative correlation between sit-and-reach and age in the AA + CC genotype than in the AC genotype (AA + CC: r = −0.216, p < 0.001; AC: r = −0.089, p = 0.04; interaction p = 0.037). However, none of these results survived correction for multiple testing. Further studies are warranted to investigate the association between the COL1A1 gene variation and exercise-related phenotypes.
Keywords:
genotype; heterozygosity; polymorphism; COL1A1; aging; flexibility; sit-and-reach; sports climbing 1. Introduction
Sport climbing has become increasingly popular and was selected for inclusion in the 2020 Olympic Games. In particular, bouldering and lead climbing are performed professionally by athletes and as recreational activities by the general population. Climbing performance is associated with various physical and morphological characteristics. Previous studies have suggested that climbing performance is correlated with endurance performance, flexibility, and grip strength relative to body mass [1]. In addition, elite climbers have significantly lower fat mass percentage [2,3,4] and a higher grip strength relative to their body mass and flexibility when compared with controls and non-elite climbers [5,6]. These physical and morphological characteristics are affected by environmental and genetic factors. Furthermore, the heritability of athletic performance was estimated to be 66% in a twin study [7]. Saito, et al. [8] reported that the frequency of the X allele in ACTN3 R577X polymorphism, which is associated with high flexibility [9], was higher in climbers than in controls.
Flexibility is influenced by both genetic and environmental factors. Previous studies on monozygotic and dizygotic twins suggest that 18–55% of the heritability in flexibility (based on assessments of “sit-and-reach”) is explained by genetic factors [10,11,12]. Gene polymorphisms associated with flexibility include the range of motion, muscle stiffness, and general joint laxity [9,13,14,15,16]. A recent study showed that COL1A1 gene rs1107946 polymorphism is associated with muscle stiffness and injury [13]. Flexibility is influenced by the properties of intramuscular tissues [17]. Among the 28 identified types, type I collagen is an important component that affects intramuscular connective tissues properties [18], such as hysteresis, elasticity, and viscosity [19]. Therefore, elongation of the muscle tissue across the joint may affect the range of motion measured by the sit-and-reach test. A previous study suggested that the C allele in the COL1A1 rs1107946 polymorphism tends to have a larger type I collagen α1/α2 chain ratio than the AA genotype, and lower muscle stiffness was shown in the C allele compared with the AA genotype [13]. Hence, COL1A1 gene rs1107946 polymorphism may also be associated with the range of motion, but this remains to be verified.
Flexibility gradually decreases with age [20] and is affected by factors such as physical training and stretching [21,22,23,24]. The mechanical alteration of collagen molecules, which is important in the extracellular matrix, may affect the age-related decrease in flexibility. Pro-collagen I genes in the muscle are expressed in lower levels in older men than in younger men [25]. In addition, collagen turnover decreases in mature mice and induces fibrosis [26]. Therefore, age-related decrease in flexibility may be affected by genetic factors, such as COL1A1 rs1107946 polymorphism. In addition, continuous training and stretching to improve athletic performance are adapted to athlete-specific physical fitness. For example, elite climbers have higher flexibility than recreational climbers [6,27]. Therefore, the climbers’ status and flexibility in general populations may be influenced by COL1A1 rs1107946 polymorphism.
The purpose of this study was (1) to compare the frequency of COL1A1 rs1107946 polymorphism between climbers and controls in three ethnic groups (Japanese, Polish, and Russian), and (2) to investigate the effect of COL1A1 rs1107946 polymorphism on the age-related decrease in flexibility in the general population.
2. Materials and Methods
2.1. Study I
2.1.1. Subjects
Study I consisted of 1929 healthy people (controls) and 218 climbers. The Japanese cohort included 1093 controls (n = 435 males, n = 658 females; age 54.3 ± 15.2 years) and 62 climbers (38 males, 24 females; age 23.2 ± 8.4 years). The Polish cohort included 635 controls (n = 505 males, n = 130 females; age 25.1 ± 5.0 years) and 126 climbers (97 males, 29 females; age 28.4 ± 6.1 years). The Russian cohort included 201 controls (n = 158 males, n = 43 females; age 45 ± 4.3 years) and 30 climbers (17 men, 13 women; age 22.8 ± 4.1 years). All participants were informed of the purpose and methods of the study, and each provided written informed consent for participation. This study was approved by the ethics committees of the Nippon Sport Science University, Gdańsk University of Physical Education and Sport, and the Federal Research and Clinical Center of Physical-Chemical Medicine of Federal Medical Biological Agency. This study was conducted in accordance with the principles of the Declaration of Helsinki for Human Research.
2.1.2. Genotyping
Japanese Climbers and Controls and Polish Climbers
Total DNA was extracted and isolated from the saliva of the participants using an Oragene-DNA kit (DNA Genotek, ON, Canada). The COL1A1 rs1107946 polymorphism was genotyped using the TaqMan SNP Genotyping Assay (Assay ID: COL1A1 gene rs1107946 polymorphisms: C___7477171_10) using a Bio-Rad PCR System (CFD-3120J1, Bio-Rad, Hercules, CA, USA). The genotyping mixture (total volume: 5 μL) contained 2.5 μL of GTXpress Master Mix, 0.125 μL of assay mix (40×), and 1.375 μL of distilled water, with 1 μL of genomic DNA (10 ng/μL) per reaction. The thermal cycling conditions included an initial denaturation at 95 °C for 20 s, followed by 40 cycles of denaturation at 95 °C for 3 s, and annealing/extension at 60 °C for 20 s.
Polish Controls
To obtain the DNA of COL1A1 rs1107946 polymorphisms from the Polish controls, samples were extracted from the buccal cells using a High Pure PCR Template Preparation Kit (Roche, Switzerland) according to the manufacturer’s instructions. The genotyping mixture (total volume: 5 μL) contained 2.5 μL of TaqPath ProAmp Master Mix (Thermo Fisher Scientific, Waltham, MA, USA), 0.25 μL of assay mix (10×), and 1 μL of distilled water with 1.25 μL of genomic DNA (~10 ng/μL) per reaction. The thermal cycling conditions included a pre-read at 60 °C for 30 s, an initial denaturation at 95 °C for 5 min, followed by 40 cycles of denaturation at 95 °C for 5 s, annealing/extension at 60 °C for 30 s, and a post-read at 60 °C for 30 s. Genotyping reactions were performed on a CFX Connect Real-Time PCR Detection System (BioRad, Hercules, CA, USA).
Russian Climbers and Controls
In Russian climbers and controls, molecular genetic analysis was performed using DNA samples obtained from leukocytes (venous blood). Venous blood samples (4 mL) were collected in tubes containing ethylenediaminetetraacetic acid (EDTA) (Vacuette EDTA tubes, Greiner Bio-One, Kremsmünster, Austria). Blood samples were transported to the laboratory at 4 °C, and the DNA was extracted on the same day. DNA extraction and purification were performed using a commercial kit (Techno-Sorb, Moscow, Russia) according to the manufacturer’s instructions (Technoclon, Moscow, Russia). The protocol included chemical lysis, selective DNA binding on silica spin columns, and ethanol washing. The quality of the extracted DNA was assessed using agarose gel electrophoresis. HumanOmni1-Quad BeadChips (Illumina Inc., San Diego, CA, USA) were used for genotyping 1,140,419 single-nucleotide polymorphisms (SNPs; including COL1A1 polymorphism) in athletes and controls. The assay required 200 ng of DNA sample input at a concentration of at least 50 ng/µL. The exact concentrations of DNA in each sample were measured using a Qubit Fluorometer (Invitrogen, Waltham, Massachusetts, USA). All further procedures were performed according to the instructions provided for the Infinium HD Assay (Illumina, San Diego, CA, USA).
2.2. Study II
2.2.1. Subjects
Study II included 1093 healthy Japanese individuals (435 men, age: 52.7 ± 15.7 years, height: 170.4 ± 6.4 cm, weight: 69.3 ± 10.6 kg, and 658 women: age: 55.4 ± 14.7 years, height: 156.7 ± 5.7 cm, weight: 54.1 ± 8.2 kg). Flexibility was measured using a sit-and-reach test (in cm) with a digital measuring device (Takei Scientific Instruments Co. Ltd., Tokyo, Japan). The subjects were instructed to sit on the floor and stretch their legs out in front of them with their knees straight. The subjects were allowed two to three attempts to attain the highest possible rating. This study was approved by the Ethics Committee of Nippon Sport Science University. This study was conducted in accordance with the principles of the Declaration of Helsinki for Human Research.
2.2.2. Genotyping
We used the same protocol as in study I of Japanese controls.
2.3. Statistical Analysis
The SPSS statistical package version 25.0 for Mac was used to perform all statistical analyses. Genotype and allele frequencies were calculated for all gene polymorphisms, and Hardy–Weinberg equilibrium was assessed using the chi-square (×2) test. In addition, a meta-analysis of the three ethnic cohorts was conducted to investigate the association between the frequency of each gene polymorphism in climbers and controls. The meta-analysis was conducted using the Review Manager software program (version 5.3; Copenhagen: The Nordic Cochrane Center, The Cochrane Collaboration) [28]. Random effect models were applied. Odds ratio with 95% confidence intervals (CI) was estimated using the Mantel–Haenszel method. The heterogeneity degree between the studies was assessed with the I2 statistics.
Age-related decrease in sit-and-reach was examined using correlation analysis. The association between COL1A1 gene rs1107946 polymorphism and age, height, and weight were examined using one-way analysis of variance (ANOVA). In addition, the association between COL1A1 gene rs1107946 polymorphism and sit-and-reach was examined by one-way analysis of covariance (ANCOVA), adjusting for sex and age. Age-related effects on the association between COL1A1 gene rs1107946 polymorphism and sit-and-reach were assessed from the difference in the r-value (slope of the line) of the correlation between age and sit-and-reach, and two-way ANCOVA was adjusted for sex. A Bonferroni adjustment for multiple testing (climbing status and flexibility) was made with statistical significance set at p < 0.025.
3. Results
3.1. Study I
The COL1A1 rs1107946 frequencies among climbers and controls from Poland and Russia were in Hardy–Weinberg equilibrium (Polish: climbers p = 0.383, controls p = 0.868; Russian: climbers p = 0.461, controls p = 0.090), but not in Japanese climbers (p = 0.035). There were no significant differences in the distribution of COL1A1 rs1107946 between the climbers and controls in any ethnic cohort for genotype and allele models (Table 1). In addition, the odds ratio of each genetic model (dominant, recessive, and additive) was not significantly associated with any ethnic cohort (Table 2). There was no significant sex difference of genotype frequency in Japanese (p = 0.657), Polish (p = 0.721), and Russian (0.549).

Table 1.
Frequency of genotype and allele model of COL1A1 rs1107946 polymorphism in Japanese, Polish, and Russian climbers and controls.
Table 2.
Odds ratio of each genetic model (dominant, recessive, and additive) in Japanese, Polish, and Russian climbers.
The results of the meta-analysis across the three cohorts showed a trend towards significance of AC genotype frequency in climbers compared with AA and CC genotype (Figure 1). There were no associations in the dominant and recessive models. The frequency of the AC genotypes in the COL1A1 rs1107946 polymorphism was higher in Japanese, Polish, and Russian climbers than in the controls (Figure 1 odds ratio: 1.41, 95% confidence interval (CI):1.03–1.93, p = 0.03). No heterogeneity between studies (I2 = 0%; p = 0.98) was observed.
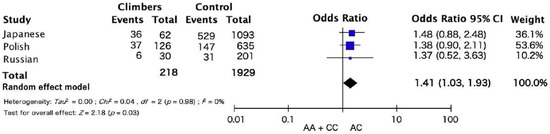
Figure 1.
Meta-analysis results of three ethnic cohorts for the association COL1A1 rs1107946 polymorphism and climbing status.
3.2. Study II
The frequencies of COL1A1 rs1107946 polymorphism among the Japanese controls were in Hardy–Weinberg equilibrium (p = 0.389). We show the associations between COL1A1 rs1107946 polymorphism and sit-and-reach in Table 3. There was a trend of associations between sit-and-reach and COL1A1 rs1107946 polymorphism in genotype, recessive, and over-dominant models. A trend towards significance in sit-and-reach was observed when CC + AC were compared with AA genotype (recessive model: p = 0.035). In addition, the AC genotype showed higher flexibility than the AA + CC genotype (over-dominant model: p = 0.026). There was significant negative correlation between sit-and-reach and age (r = −0.151, p < 0.001), with an interaction effect between them when the AA + CC genotype group was compared with the AC genotype (AA + CC: r = 0.21, p = 2.35 × 10–7, AC: r = −0.09, p = 0.04; interaction p = 0.037; Figure 2) but not in other the models.
Table 3.
Association between COL1A1 rs1107946 and sit and reach.
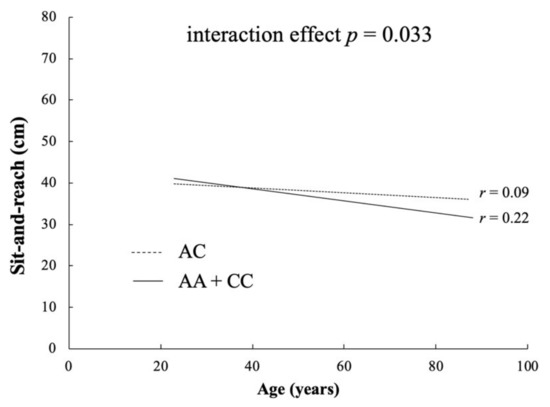
Figure 2.
Correlation between sit-and-reach and age for the AC and AA + CC genotypes in COL1A1 rs1107946 polymorphism.
4. Discussion
One of the purposes of this study was to examine the frequency of COL1A1 rs1107946 polymorphism in climbers, who require higher flexibility and strength, from three ethnicities (Study I). The results of the meta-analysis across the three ethnicities (Japanese, Polish, and Russian) showed that the frequency of the AC genotype was higher in climbers than in the controls (p = 0.03). In addition, our results showed a trend towards significance between the COL1A1 rs1107946 C allele (AC and CC genotype) and flexibility in the general population (Study II). There was a tendency of interaction effect of correlation between sit-and-reach and age in the AC genotype and the AA + CC genotype (AA + CC: r = −0.216, p < 0.001; AC: r = −0.089, p = 0.04; interaction p = 0.037). These results suggest that the AC genotype may have an advantage over the age-related decrease in flexibility, such as sit-and-reach. However, none of these results survived correction for multiple testing (p < 0.025).
In the present study, our meta-analysis across three ethnicities (Japanese, Polish, and Russian) shows that the frequency of the AC genotype is higher in climbers than in controls (p = 0.03). Flexibility is an important component of climbing performance. Sit-and-reach tests were positively correlated with climbing performance [1]. Studies have suggested that sit-and-reach is higher in elite climbers than in recreational climbers [6,27]. In addition, climbing-specific flexibility, such as foot raises in front of the wall, is higher in elite climbers than recreational- and non-climbers [5]. These flexibilities may be influenced by muscle strength. Collectively, our results suggest that climbers require both the range of motion and the muscle strength to control it. Therefore, the AC genotype in COL1A1 might be related to not only flexibility, but also sport climbing performance.
A previous study reported an association between COL1A1 rs1107946 polymorphism and flexibility (e.g., muscle stiffness) [13]. The study suggested that the C allele (CC + AC genotype) of COL1A1 rs1107946 polymorphism shows lower muscle stiffness than the AA genotype. Our results are consistent with this finding. In addition, we observed an age-related decrease in flexibility (r = −0.151, p < 0.001), which was the same in men (r = 0.186, p < 0.001) and women (r = −0.156, p < 0.001). Physical ability, including flexibility and muscle strength, is impaired with age [20]. The collagen turnover decreases with aging [26], which increases the non-enzymatic cross-linking of collagen fibers through the accumulation of advanced glycation end-products [29]. Previous studies have suggested that the fibrotic phenotype increases muscle stiffness [30]. The fibrotic collagen phenotype may be prevented by the higher COL1A1 mRNA expression, promoting the turnover of collagens. A previous study suggested that age-related decrease in muscle strength and muscle cross-sectional area may be suppressed by genetic factors [31,32]. Our study showed a higher age-related decrease in the AA + CC genotype compared to the AC genotype in COL1A1 rs1107946 polymorphism. Thus, the AC genotype in COL1A1 rs1107946 polymorphism may weaken the age-related decrease in flexibility.
The sit-and-reach test in our study may reflect muscle and musculotendinous stiffness and muscle strength. Furthermore, muscle stiffness and strength also exhibit a trade-off relationship. Alfuraih, et al. [33] suggested that muscle stiffness measured in the lower limb muscles is positively correlated with isokinetic knee strength. A previous study on the osteogenesis imperfecta murine (oim) model suggested that oim/oim mice exclusively produce homotrimeric (three α1 chains) type I collagen and had decreased tetanic force and body weight compared to the +/oim genotypes [34]. Higher levels of type I collagen may reduce muscle strength. In contrast, the C allele of COL1A1 rs1107946 polymorphism results in a higher collagen type I α1/α2 ratio and lower muscle stiffness [13]. Therefore, muscle strength and stiffness, which influence sit-and-reach, may be differently affected by the COL1A1 rs1107946 polymorphism.
The athletic ability that requires both flexibility and muscle strength, such as sit-and-reach and climbing-specific flexibility, may require both genetic characteristics of COL1A1 rs1107946 gene polymorphism. Heterozygosity is a measure of genetic diversity and may have an effect on the phenotypes [35,36]. For example, muscle injury and decreased bone mineral density (BMD) have opposing relationships with the COL1A1 rs1107946 polymorphism. Miyamoto-Mikami, et al. [13] reported that athletes with muscle injury have a higher frequency of the AA + AC genotype in COL1A1 rs1107946 polymorphism compared to the CC genotype. In contrast, a previous study suggested that the CC genotype carriers had significantly lower lumbar spine BMD than AA genotype carriers [13]. The CC genotype in COL1A1 rs1107946 polymorphism may be a risk factor for low BMD and fatigue fractures [13,37,38,39]. Therefore, there is a trade-off between muscle and bone injury: the A allele in COL1A1 is associated with higher BMD, while the C allele decreases the risk of muscle injury.
The AC genotype has a heterozygous effect on complex phenotypes, such as sit-and-reach and climbing status, which require flexibility and muscle strength. However, the function of the AC genotype in COL1A1 rs1107946 remains unclear and would require more functional and longitudinal studies in the future.
5. Conclusions
In our meta-analysis of three ethnicities, sports climbers, who require higher flexibility and muscle strength, have a higher AC genotype frequency than control participants. In addition, the C allele, especially in AC genotype, might be related to flexibility in general populations. However, none of these results survived correction for multiple testing. Further studies are warranted to investigate the association between the COL1A1 gene variation and exercise-related phenotypes.
Author Contributions
Conceptualization, M.S., I.I.A., and N.K.; data collection and analysis, M.S., M.G., E.A.S., M.M., K.H.-L., M.M.-S., H.H., P.C., T.O., A.K.L., E.V.G., P.M., K.N., I.I.A. and N.K. All authors have read and agreed to the published version of the manuscript.
Funding
The Japanese part of the study was supported by the grant from the program Grants-in-Aid for Scientific Research (C) (19K11531 to N.K.).
Institutional Review Board Statement
The study was conducted according to the guidelines of the Declaration of Helsinki and approved by the Ethics Committee of the Federal Research and Clinical Center of Physical-Chemical Medicine and by the ethics committees of the Nippon Sport Science University.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Acknowledgments
The authors would like to thank Oleg Borisov for assistance with statistical analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- MacKenzie, R.; Monaghan, L.; Masson, R.A.; Werner, A.K.; Caprez, T.S.; Johnston, L.; Kemi, O.J. Physical and Physiological Determinants of Rock Climbing. Int. J. Sports Physiol. Perform. 2020, 15, 168–179. [Google Scholar] [CrossRef] [PubMed]
- Philippe, M.; Wegst, D.; Muller, T.; Raschner, C.; Burtscher, M. Climbing-specific finger flexor performance and forearm muscle oxygenation in elite male and female sport climbers. Eur. J. Appl. Physiol. 2012, 112, 2839–2847. [Google Scholar] [CrossRef] [PubMed]
- Ozimek, M.; Krawczyk, M.; Zadarko, E.; Barabasz, Z.; Ambrozy, T.; Stanula, A.; Mucha, D.K.; Jurczak, A.; Mucha, D. Somatic Profile of the Elite Boulderers in Poland. J. Strength Cond. Res. 2017, 31, 963–970. [Google Scholar] [CrossRef] [PubMed]
- Giles, D.; Barnes, K.; Taylor, N.; Chidley, C.; Chidley, J.; Mitchell, J.; Torr, O.; Gibson-Smith, E.; Espana-Romero, V. Anthropometry and performance characteristics of recreational advanced to elite female rock climbers. J. Sports Sci. 2021, 39, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Draper, N.; Brent, S.; Hodgson, C.; Blackwell, G. Flexibility assessment and the role of flexibility as a determinant of performance in rock climbing. Int. J. Perform. Anal. Sport 2009, 9, 67–89. [Google Scholar] [CrossRef]
- Grant, S.; Hynes, V.; Whittaker, A.; Aitchison, T. Anthropometric, strength, endurance and flexibility characteristics of elite and recreational climbers. J. Sports Sci. 1996, 14, 301–309. [Google Scholar] [CrossRef]
- De Moor, M.H.; Spector, T.D.; Cherkas, L.F.; Falchi, M.; Hottenga, J.J.; Boomsma, D.I.; De Geus, E.J. Genome-wide linkage scan for athlete status in 700 British female DZ twin pairs. Twin Res. Hum. Genet. 2007, 10, 812–820. [Google Scholar] [CrossRef]
- Saito, M.; Ginszt, M.; Semenova, E.A.; Massidda, M.; Huminska-Lisowska, K.; Michałowska-Sawczyn, M.; Homma, H.; Cięszczyk, P.; Okamoto, T.; Larin, A.K.; et al. Genetic profile of sports climbing athletes from three different ethnicities. Biol. Sport 2022, 39, 913–919. [Google Scholar] [CrossRef]
- Kikuchi, N.; Zempo, H.; Fuku, N.; Murakami, H.; Sakamaki-Sunaga, M.; Okamoto, T.; Nakazato, K.; Miyachi, M. Association between ACTN3 R577X Polymorphism and Trunk Flexibility in 2 Different Cohorts. Int. J. Sports Med. 2017, 38, 402–406. [Google Scholar] [CrossRef]
- Okuda, E.; Horii, D.; Kano, T. Genetic and Environmental Effects on Physical Fitness and Motor Performance. Int. J. Sport Health Sci. 2005, 3, 1–9. [Google Scholar] [CrossRef]
- Maes, H.H.; Beunen, G.P.; Vlietinck, R.F.; Neale, M.C.; Thomis, M.; Vanden Eynde, B.; Lysens, R.; Simons, J.; Derom, C.; Derom, R. Inheritance of physical fitness in 10-yr-old twins and their parents. Med. Sci. Sports Exerc. 1996, 28, 1479–1491. [Google Scholar] [CrossRef]
- Satipati Chatterjee, N.D. Physical and Motor Fitness in Twins. Jpn. J. Physiol. 1995, 45, 519–534. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto-Mikami, E.; Kumagai, H.; Tanisawa, K.; Taga, Y.; Hirata, K.; Kikuchi, N.; Kamiya, N.; Kawakami, R.; Midorikawa, T.; Kawamura, T.; et al. Female Athletes Genetically Susceptible to Fatigue Fracture Are Resistant to Muscle Injury: Potential Role of COL1A1 Variant. Med. Sci. Sports Exerc. 2021, 53, 1855–1864. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, H.; Miyamoto-Mikami, E.; Hirata, K.; Kikuchi, N.; Kamiya, N.; Hoshikawa, S.; Zempo, H.; Naito, H.; Miyamoto, N.; Fuku, N. ESR1 rs2234693 Polymorphism Is Associated with Muscle Injury and Muscle Stiffness. Med. Sci. Sports Exerc. 2019, 51, 19–26. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, N.; Miyamoto-Mikami, E.; Hirata, K.; Kimura, N.; Fuku, N. Association analysis of the ACTN3 R577X polymorphism with passive muscle stiffness and muscle strain injury. Scand. J. Med. Sci. Sports 2018, 28, 1209–1214. [Google Scholar] [CrossRef]
- Abrahams, S.; Posthumus, M.; Collins, M. A polymorphism in a functional region of the COL5A1 gene: Association with ultraendurance-running performance and joint range of motion. Int. J. Sports Physiol. Perform. 2014, 9, 583–590. [Google Scholar] [CrossRef]
- Gajdosik, R.L. Passive extensibility of skeletal muscle: Review of the literature with clinical implications. Clin. Biomech. 2001, 16, 87–101. [Google Scholar] [CrossRef]
- Light, N.; Champion, A.E. Characterization of muscle epimysium, perimysium and endomysium collagens. Biochem. J. 1984, 219, 1017–1026. [Google Scholar] [CrossRef]
- Alter, M.J. Science of Flexibility; Human Kinetics: Champaign, IL, USA, 1996. [Google Scholar]
- McKay, M.J.; Baldwin, J.N.; Ferreira, P.; Simic, M.; Vanicek, N.; Burns, J. Normative reference values for strength and flexibility of 1,000 children and adults. Neurology 2017, 88, 36–43. [Google Scholar] [CrossRef]
- Kim, E.; Dear, A.; Ferguson, S.L.; Seo, D.; Bemben, M.G. Effects of 4 weeks of traditional resistance training vs. superslow strength training on early phase adaptations in strength, flexibility, and aerobic capacity in college-aged women. J. Strength Cond. Res. 2011, 25, 3006–3013. [Google Scholar] [CrossRef]
- Monteiro, W.D.; Simao, R.; Polito, M.D.; Santana, C.A.; Chaves, R.B.; Bezerra, E.; Fleck, S.J. Influence of strength training on adult women’s flexibility. J. Strength Cond. Res. 2008, 22, 672–677. [Google Scholar] [CrossRef] [PubMed]
- Santos, E.; Rhea, M.R.; Simao, R.; Dias, I.; Salles, B.F.; Novaes, J.; Leite, T.; Blair, J.C.; Bunker, D.J. Influence of moderately intense strength training on flexibility in sedentary young women. J. Strength Cond. Res. 2010, 24, 3144–3149. [Google Scholar] [CrossRef] [PubMed]
- Medeiros, D.M.; Cini, A.; Sbruzzi, G.; Lima, C.S. Influence of static stretching on hamstring flexibility in healthy young adults: Systematic review and meta-analysis. Physiother. Theory Pract. 2016, 32, 438–445. [Google Scholar] [CrossRef]
- Welle, S.; Brooks, A.; Thornton, C.A. Senescence-related changes in gene expression in muscle: Similarities and differences between mice and men. Physiol. Genom. 2001, 5, 67–73. [Google Scholar] [CrossRef]
- Podolsky, M.J.; Yang, C.D.; Valenzuela, C.L.; Datta, R.; Huang, S.K.; Nishimura, S.L.; Dallas, S.L.; Wolters, P.J.; Le Saux, C.J.; Atabai, K. Age-dependent regulation of cell-mediated collagen turnover. JCI Insight 2020, 5, e137519. [Google Scholar] [CrossRef]
- Grant, S.; Hasler, T.; Davies, C.; Aitchison, T.C.; Wilson, J.; Whittaker, A. A comparison of the anthropometric, strength, endurance and flexibility characteristics of female elite and recreational climbers and non-climbers. J. Sports Sci. 2001, 19, 499–505. [Google Scholar] [CrossRef] [PubMed]
- Cochrane Training. Available online: http://tech.cochrane.org/revman (accessed on 3 December 2021).
- Haus, J.M.; Carrithers, J.A.; Trappe, S.W.; Trappe, T.A. Collagen, cross-linking, and advanced glycation end products in aging human skeletal muscle. J. Appl. Physiol. 2007, 103, 2068–2076. [Google Scholar] [CrossRef]
- Wood, L.K.; Kayupov, E.; Gumucio, J.P.; Mendias, C.L.; Claflin, D.R.; Brooks, S.V. Intrinsic stiffness of extracellular matrix increases with age in skeletal muscles of mice. J. Appl. Physiol. 2014, 117, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, N.; Yoshida, S.; Min, S.K.; Lee, K.; Sakamaki-Sunaga, M.; Okamoto, T.; Nakazato, K. The ACTN3 R577X genotype is associated with muscle function in a Japanese population. Appl. Physiol. Nutr. Metab. 2015, 40, 316–322. [Google Scholar] [CrossRef]
- Zempo, H.; Tanabe, K.; Murakami, H.; Iemitsu, M.; Maeda, S.; Kuno, S. Age differences in the relation between ACTN3 R577X polymorphism and thigh-muscle cross-sectional area in women. Genet. Test. Mol. Biomarkers. 2011, 15, 639–643. [Google Scholar] [CrossRef]
- Alfuraih, A.M.; Tan, A.L.; O’Connor, P.; Emery, P.; Wakefield, R.J. The effect of ageing on shear wave elastography muscle stiffness in adults. Aging Clin. Exp. Res. 2019, 31, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Gentry, B.A.; Ferreira, J.A.; McCambridge, A.J.; Brown, M.; Phillips, C.L. Skeletal muscle weakness in osteogenesis imperfecta mice. Matrix Biol. 2010, 29, 638–644. [Google Scholar] [CrossRef]
- Leznicka, K.; Zyzniewska-Banaszak, E.; Gebska, M.; Machoy-Mokrzynska, A.; Krajewska-Pedzik, A.; Maciejewska-Skrendo, A.; Leonska-Duniec, A. Interactions between Gene Variants within the COL1A1 and COL5A1 Genes and Musculoskeletal Injuries in Physically Active Caucasian. Genes 2021, 12, 1056. [Google Scholar] [CrossRef] [PubMed]
- Samsami, D.A.; Doroudchi, M.; Kalantari, T.; Pezeshki, A.M.; Ghaderi, A. Heterozygosity in CTLA-4 gene and severe preeclampsia. Int. J. Gynaecol. Obstet. 2005, 88, 19–24. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Singh, P.; Singh, S.; Juneja, P.K.; Kaur, T. A haplotype derived from the common variants at the −1997G/T and Sp1 binding site of the COL1A1 gene influences risk of postmenopausal osteoporosis in India. Rheumatol. Int. 2013, 33, 501–506. [Google Scholar] [CrossRef]
- Kostik, M.M.; Smirnov, A.M.; Demin, G.S.; Mnuskina, M.M.; Scheplyagina, L.A.; Larionova, V.I. Genetic polymorphisms of collagen type I alpha1 chain (COL1A1) gene increase the frequency of low bone mineral density in the subgroup of children with juvenile idiopathic arthritis. EPMA J. 2013, 4, 15. [Google Scholar] [CrossRef][Green Version]
- Stewart, T.L.; Jin, H.; McGuigan, F.E.; Albagha, O.M.; Garcia-Giralt, N.; Bassiti, A.; Grinberg, D.; Balcells, S.; Reid, D.M.; Ralston, S.H. Haplotypes defined by promoter and intron 1 polymorphisms of the COLIA1 gene regulate bone mineral density in women. J. Clin. Endocrinol. Metab. 2006, 91, 3575–3583. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).