
FDDM1 and FDDM2, Two SGS3-like Proteins, Function as a Complex to Affect DNA Methylation in Arabidopsis

 , ,
, , 
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Condition
2.2. Construction of Plasmids and Plant Transformation
2.3. DNA Methylation Assay
2.4. RT-PCR Analysis
2.5. DNA/RNA Binding Assay
2.6. Yeast Two-Hybrid Assay
2.7. BiFC and Co-IP Assays
3. Results
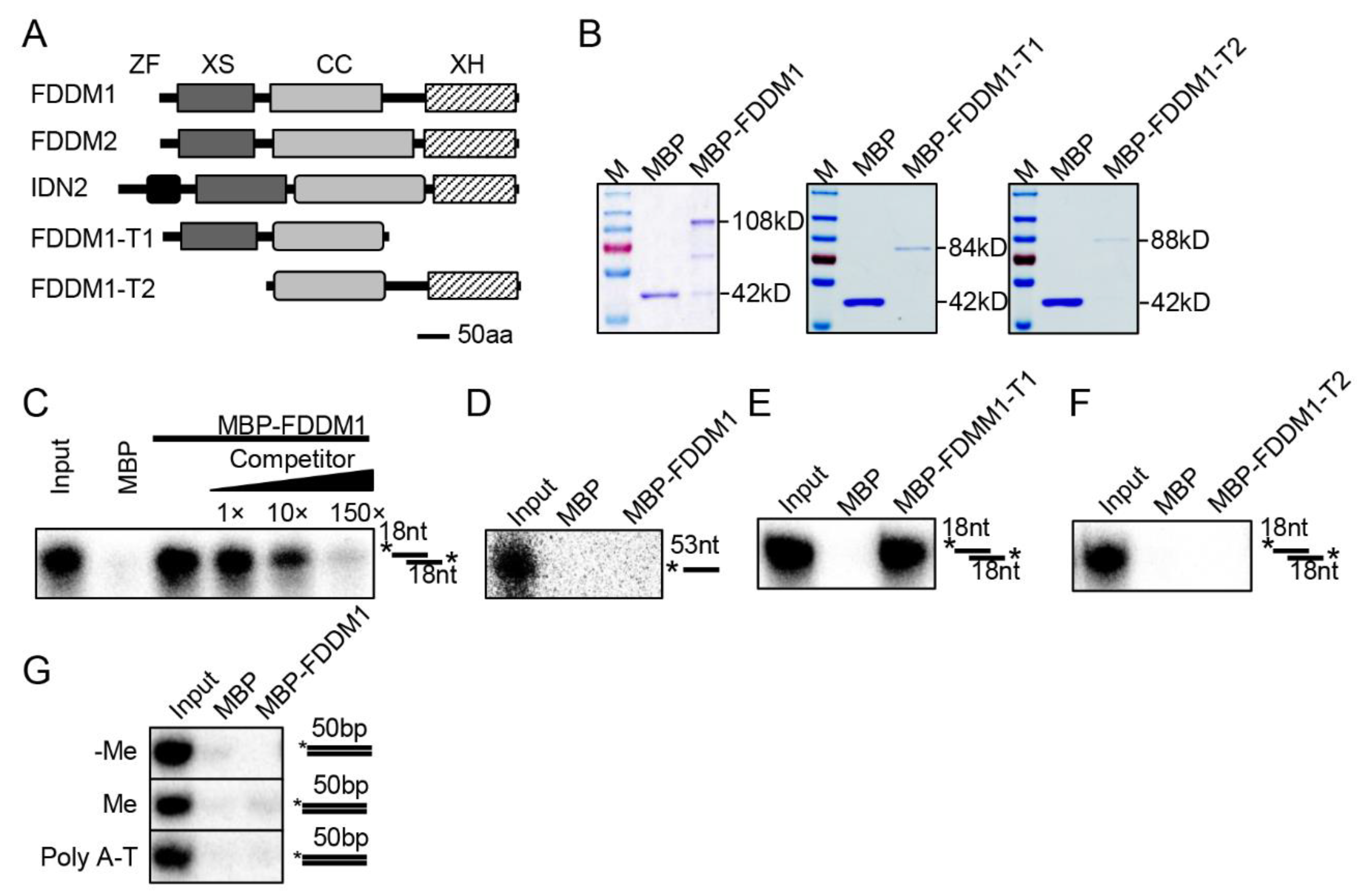
3.1. XS Domain of FDDM1 Is Required for Binding of 5′ Overhang dsRNAs
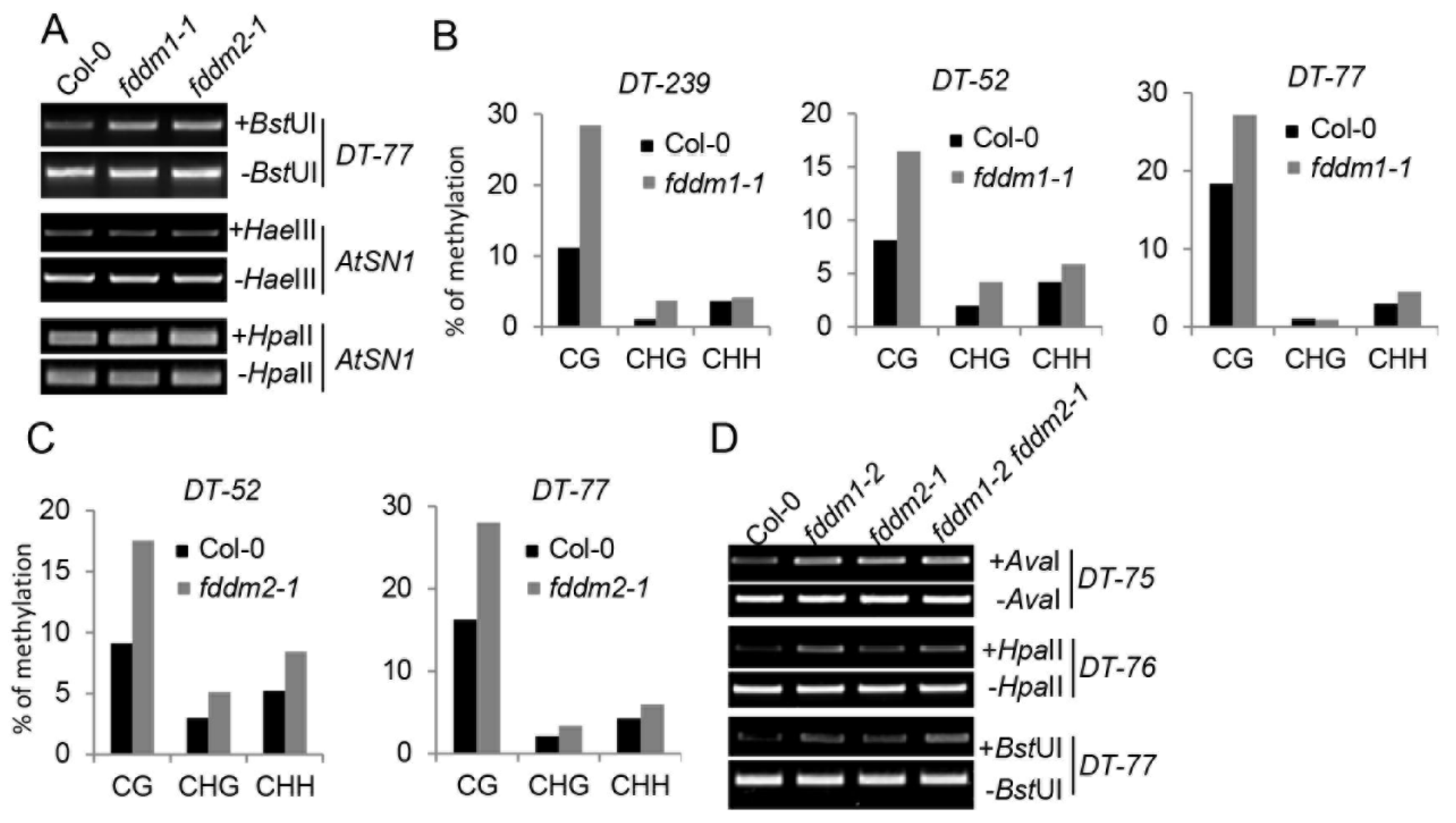
3.2. FDDM1 and FDDM2 Are Involved in DNA Demethylation
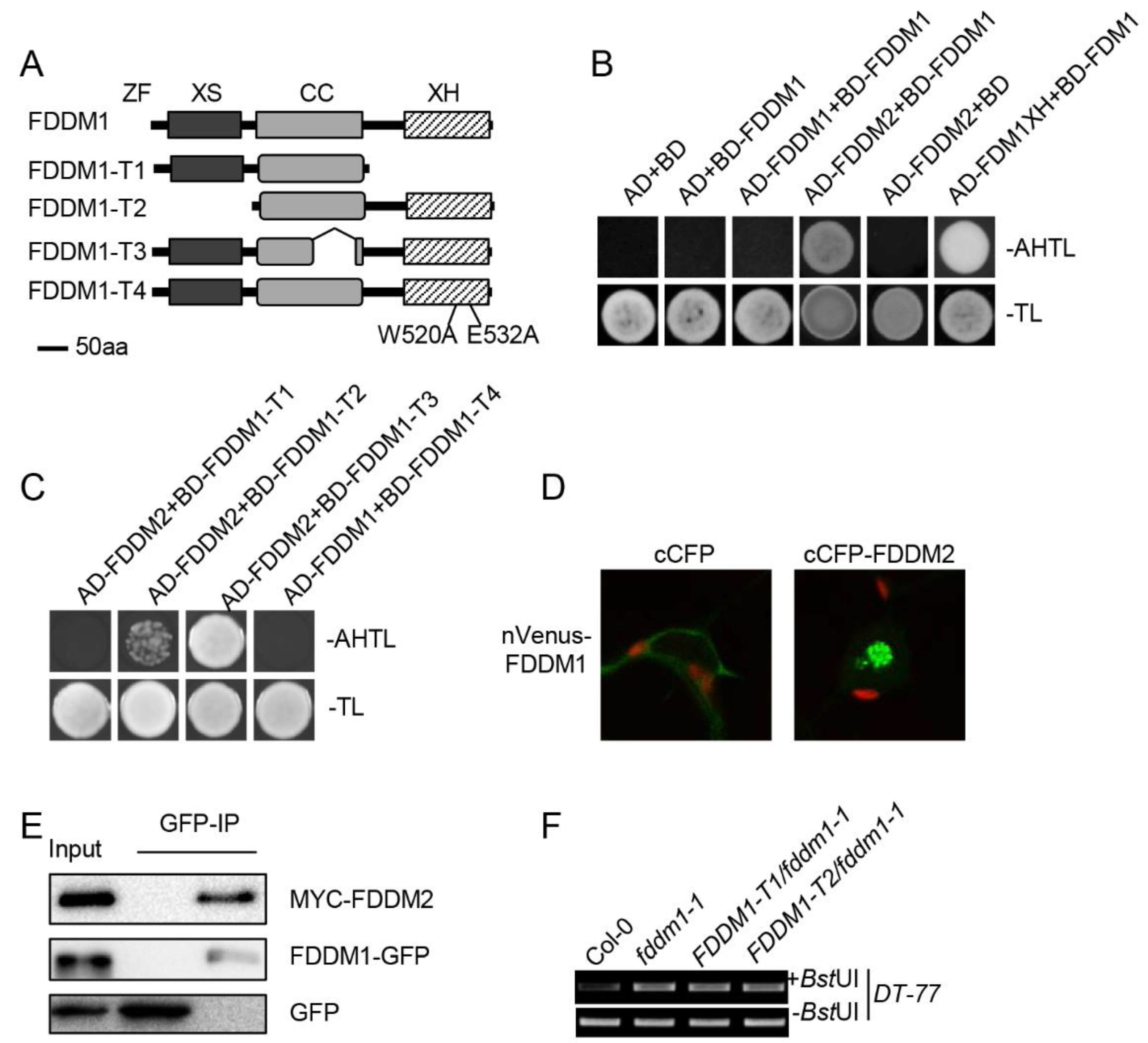
3.3. FDDM1 and FDDM2 Form a Heterodimer
3.4. XS and XH Domains of FDDM1 Are Required for Modulating DNA Methylation Levels
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Zhang, N.; Wang, Z.; Bao, Z.; Yang, L.; Wu, D.; Shu, X.; Hua, J. Mos1 functions closely with tcp transcription factors to modulate immunity and cell cycle in arabidopsis. Plant J. 2018, 93, 66–78. [Google Scholar] [CrossRef] [PubMed]
- Tirnaz, S.; Batley, J. DNA methylation: Toward crop disease resistance improvement. Trends Plant Sci. 2019, 24, 1137–1150. [Google Scholar] [CrossRef] [PubMed]
- Law, J.A.; Jacobsen, S.E. Establishing, maintaining and modifying DNA methylation patterns in plants and animals. Nat. Rev. Genet. 2010, 11, 204–220. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Mohapatra, T. Dynamics of DNA methylation and its functions in plant growth and development. Front. Plant Sci. 2021, 12, 596236. [Google Scholar] [CrossRef]
- Zhang, X.; Yazaki, J.; Sundaresan, A.; Cokus, S.; Chan, S.W.; Chen, H.; Henderson, I.R.; Shinn, P.; Pellegrini, M.; Jacobsen, S.E.; et al. Genome-wide high-resolution mapping and functional analysis of DNA methylation in arabidopsis. Cell 2006, 126, 1189–1201. [Google Scholar] [CrossRef]
- Lister, R.; O’Malley, R.C.; Tonti-Filippini, J.; Gregory, B.D.; Berry, C.C.; Millar, A.H.; Ecker, J.R. Highly integrated single-base resolution maps of the epigenome in arabidopsis. Cell 2008, 133, 523–536. [Google Scholar] [CrossRef]
- Zhong, X.; Du, J.; Hale, C.J.; Gallego-Bartolome, J.; Feng, S.; Vashisht, A.A.; Chory, J.; Wohlschlegel, J.A.; Patel, D.J.; Jacobsen, S.E. Molecular mechanism of action of plant drm de novo DNA methyltransferases. Cell 2014, 157, 1050–1060. [Google Scholar] [CrossRef]
- Kankel, M.W.; Ramsey, D.E.; Stokes, T.L.; Flowers, S.K.; Haag, J.R.; Jeddeloh, J.A.; Riddle, N.C.; Verbsky, M.L.; Richards, E.J. Arabidopsis met1 cytosine methyltransferase mutants. Genetics 2003, 163, 1109–1122. [Google Scholar] [CrossRef]
- Lindroth, A.M.; Cao, X.; Jackson, J.P.; Zilberman, D.; McCallum, C.M.; Henikoff, S.; Jacobsen, S.E. Requirement of chromomethylase3 for maintenance of cpxpg methylation. Science 2001, 292, 2077–2080. [Google Scholar] [CrossRef]
- Stroud, H.; Do, T.; Du, J.; Zhong, X.; Feng, S.; Johnson, L.; Patel, D.J.; Jacobsen, S.E. Non-cg methylation patterns shape the epigenetic landscape in arabidopsis. Nat. Struct. Mol. Biol. 2014, 21, 64–72. [Google Scholar] [CrossRef]
- Zhu, J.K. Active DNA demethylation mediated by DNA glycosylases. Annu. Rev. Genet. 2009, 43, 143–166. [Google Scholar] [CrossRef] [PubMed]
- Gong, Z.; Morales-Ruiz, T.; Ariza, R.R.; Roldan-Arjona, T.; David, L.; Zhu, J.K. Ros1, a repressor of transcriptional gene silencing in arabidopsis, encodes a DNA glycosylase/lyase. Cell 2002, 111, 803–814. [Google Scholar] [CrossRef]
- Gehring, M.; Huh, J.H.; Hsieh, T.F.; Penterman, J.; Choi, Y.; Harada, J.J.; Goldberg, R.B.; Fischer, R.L. Demeter DNA glycosylase establishes medea polycomb gene self-imprinting by allele-specific demethylation. Cell 2006, 124, 495–506. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Galisteo, A.P.; Morales-Ruiz, T.; Ariza, R.R.; Roldan-Arjona, T. Arabidopsis demeter-like proteins dml2 and dml3 are required for appropriate distribution of DNA methylation marks. Plant Mol. Biol. 2008, 67, 671–681. [Google Scholar] [CrossRef]
- Ausin, I.; Mockler, T.C.; Chory, J.; Jacobsen, S.E. Idn1 and idn2 are required for de novo DNA methylation in arabidopsis thaliana. Nat. Struct. Mol. Biol. 2009, 16, 1325–1327. [Google Scholar] [CrossRef] [PubMed]
- Ausin, I.; Greenberg, M.V.; Simanshu, D.K.; Hale, C.J.; Vashisht, A.A.; Simon, S.A.; Lee, T.F.; Feng, S.; Espanola, S.D.; Meyers, B.C.; et al. Involved in de novo 2-containing complex involved in rna-directed DNA methylation in arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 8374–8381. [Google Scholar] [CrossRef]
- Mourrain, P.; Beclin, C.; Elmayan, T.; Feuerbach, F.; Godon, C.; Morel, J.B.; Jouette, D.; Lacombe, A.M.; Nikic, S.; Picault, N.; et al. Arabidopsis sgs2 and sgs3 genes are required for posttranscriptional gene silencing and natural virus resistance. Cell 2000, 101, 533–542. [Google Scholar] [CrossRef]
- Xie, M.; Ren, G.; Costa-Nunes, P.; Pontes, O.; Yu, B. A subgroup of sgs3-like proteins act redundantly in rna-directed DNA methylation. Nucleic Acids Res. 2012, 40, 4422–4431. [Google Scholar] [CrossRef][Green Version]
- Xie, M.; Ren, G.; Zhang, C.; Yu, B. The DNA- and rna-binding protein factor of DNA methylation 1 requires xh domain-mediated complex formation for its function in rna-directed DNA methylation. Plant J. 2012, 72, 491–500. [Google Scholar] [CrossRef]
- Zhang, C.J.; Ning, Y.Q.; Zhang, S.W.; Chen, Q.; Shao, C.R.; Guo, Y.W.; Zhou, J.X.; Li, L.; Chen, S.; He, X.J. Idn2 and its paralogs form a complex required for rna-directed DNA methylation. PLoS Genet. 2012, 8, e1002693. [Google Scholar] [CrossRef]
- Zheng, Z.; Xing, Y.; He, X.J.; Li, W.; Hu, Y.; Yadav, S.K.; Oh, J.; Zhu, J.K. An sgs3-like protein functions in rna-directed DNA methylation and transcriptional gene silencing in arabidopsis. Plant J. 2010, 62, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Butt, H.; Graner, S.; Luschnig, C. Expression analysis of arabidopsis xh/xs-domain proteins indicates overlapping and distinct functions for members of this gene family. J. Exp. Bot. 2014, 65, 1217–1227. [Google Scholar] [CrossRef] [PubMed]
- Peragine, A.; Yoshikawa, M.; Wu, G.; Albrecht, H.L.; Poethig, R.S. Sgs3 and sgs2/sde1/rdr6 are required for juvenile development and the production of trans-acting sirnas in arabidopsis. Genes Dev. 2004, 18, 2368–2379. [Google Scholar] [CrossRef] [PubMed]
- Qian, W.; Miki, D.; Zhang, H.; Liu, Y.; Zhang, X.; Tang, K.; Kan, Y.; La, H.; Li, X.; Li, S.; et al. A histone acetyltransferase regulates active DNA demethylation in arabidopsis. Science 2012, 336, 1445–1448. [Google Scholar] [CrossRef]
- Li, S.; Li, M.; Liu, K.; Zhang, H.; Zhang, S.; Zhang, C.; Yu, B. Mac5, an rna-binding protein, protects pri-mirnas from serrate-dependent exoribonuclease activities. Proc. Natl. Acad. Sci. USA 2020, 117, 23982–23990. [Google Scholar] [CrossRef]
- Zhu, Y.; Rowley, M.J.; Bohmdorfer, G.; Wierzbicki, A.T. A swi/snf chromatin-remodeling complex acts in noncoding rna-mediated transcriptional silencing. Mol. Cell 2013, 49, 298–309. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Yang, W.; Liu, Y.; Li, G.; Liu, X.; Liu, Y.; Alfano, J.R.; Zhang, C.; Yu, B. FDDM1 and FDDM2, Two SGS3-like Proteins, Function as a Complex to Affect DNA Methylation in Arabidopsis. Genes 2022, 13, 339. https://doi.org/10.3390/genes13020339
Li S, Yang W, Liu Y, Li G, Liu X, Liu Y, Alfano JR, Zhang C, Yu B. FDDM1 and FDDM2, Two SGS3-like Proteins, Function as a Complex to Affect DNA Methylation in Arabidopsis. Genes. 2022; 13(2):339. https://doi.org/10.3390/genes13020339
Chicago/Turabian StyleLi, Shengjun, Weilong Yang, Yunfeng Liu, Guangyong Li, Xiang Liu, Yaling Liu, James R. Alfano, Chi Zhang, and Bin Yu. 2022. "FDDM1 and FDDM2, Two SGS3-like Proteins, Function as a Complex to Affect DNA Methylation in Arabidopsis" Genes 13, no. 2: 339. https://doi.org/10.3390/genes13020339
APA StyleLi, S., Yang, W., Liu, Y., Li, G., Liu, X., Liu, Y., Alfano, J. R., Zhang, C., & Yu, B. (2022). FDDM1 and FDDM2, Two SGS3-like Proteins, Function as a Complex to Affect DNA Methylation in Arabidopsis. Genes, 13(2), 339. https://doi.org/10.3390/genes13020339