
First Glimpse on Spring Starflower Domestication

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Morphological Assessment
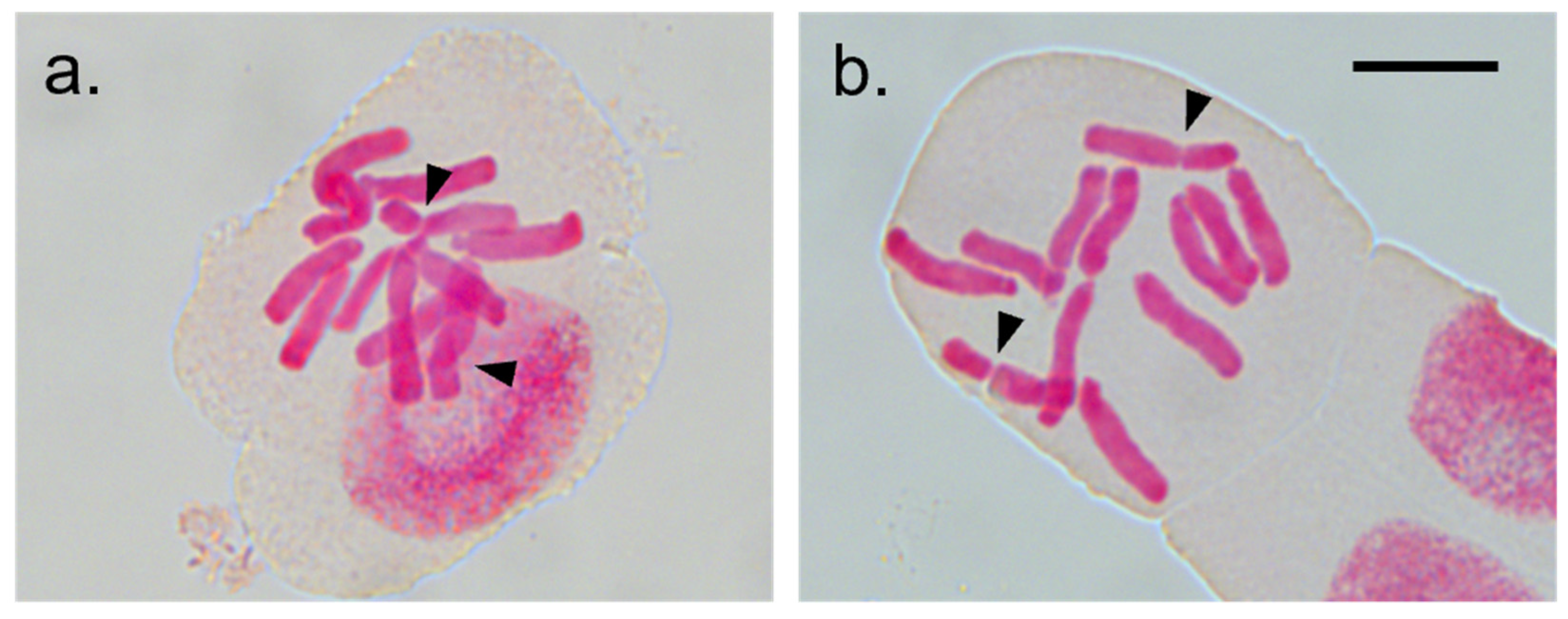
2.3. Karyological Analyses and Ploidy Estimations
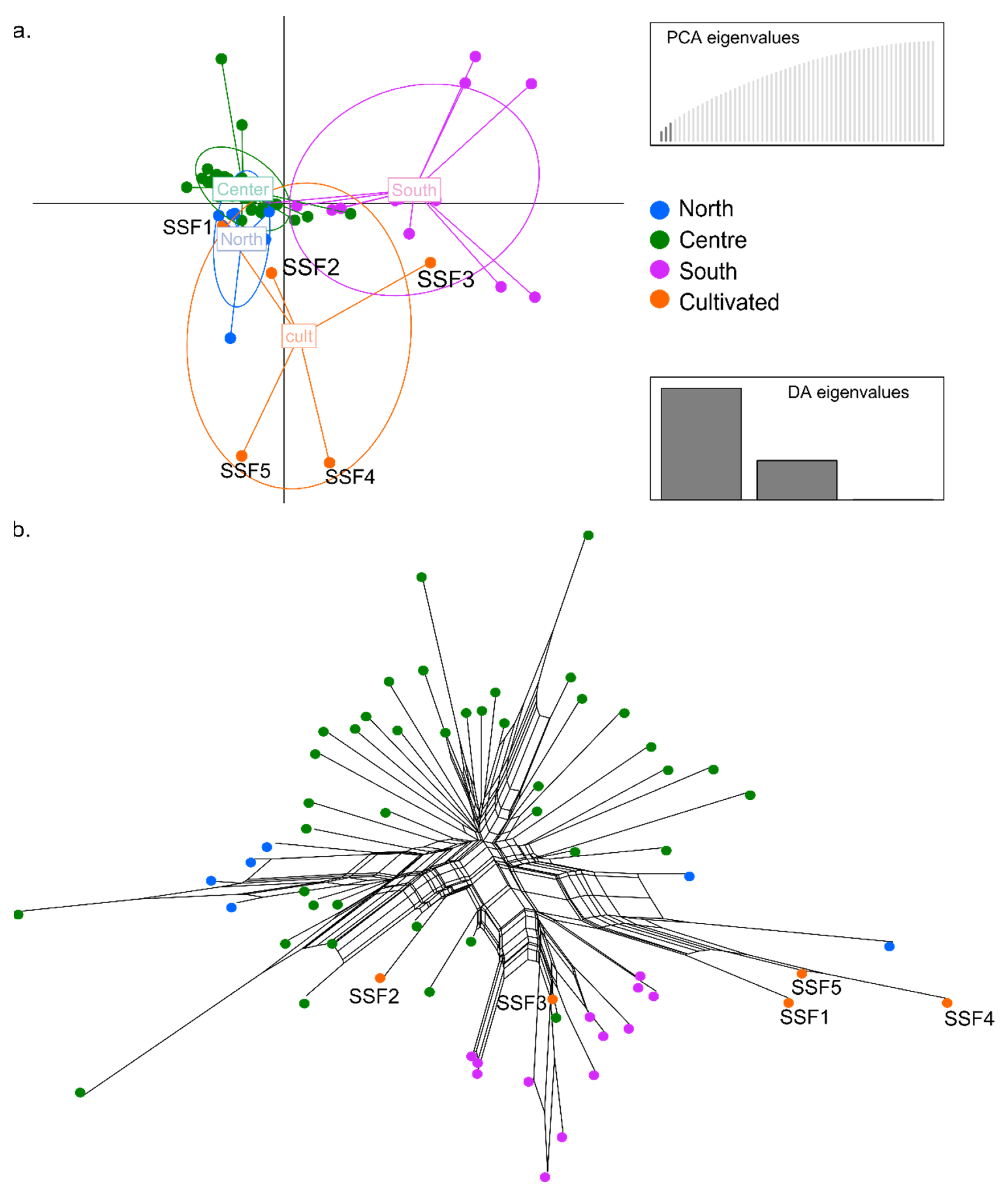
2.4. Genotyping by Sequencing (GBS) Analyses and Population Genomics of Natural and Cultivated Materials
3. Results
3.1. Morphological Characters within Ipheion uniflorum
3.2. Cytological Variability within I. uniflorum
3.3. Genomic Diversity within I. uniflorum
4. Discussion
4.1. Natural Variation within Ipheion uniflorum
4.2. Evidence for Early Domestication Mechanisms in I. uniflorum
4.3. A Glance at the Origin and Future of the Spring Starflower
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purugganan, M.D.; Fuller, D.Q. The nature of selection during plant domestication. Nature 2009, 457, 843–848. [Google Scholar] [CrossRef] [PubMed]
- Kantar, M.B.; Nashoba, A.R.; Anderson, J.E.; Blackman, B.K.; Rieseberg, L.H. The genetics and genomics of plant domestication. Bioscience 2017, 67, 971–982. [Google Scholar] [CrossRef] [Green Version]
- Meyer, R.S.; Duval, A.E.; Jensen, H.R. Patterns and processes in crop domestication: An historical review and quantitative analysis of 203 global food crops. N. Phytol. 2012, 196, 29–48. [Google Scholar] [CrossRef] [PubMed]
- Milla, R.; Bastida, J.M.; Turcotte, M.M.; Jones, G.; Violle, C.; Osborne, C.P.; Chacón-Labella, J.; Sosinski, Ê.E.; Kattge, J.; Laughlin, D.C.; et al. Phylogenetic patterns and phenotypic profiles of the species of plants and mammals farmed for food. Nat. Ecol. Evol. 2018, 2, 1808–1817. [Google Scholar] [CrossRef] [PubMed]
- Gessert, G. The rainforests of domestication: Ornamental gardens as sites of maximum genetic diversity among domesticated Plants. Leonardo 1997, 30, 129–132. [Google Scholar] [CrossRef]
- Heiser, C.B. Aspects of unconscious selection and the evolution of domesticated plants. Euphytica 1988, 37, 77–81. [Google Scholar] [CrossRef]
- Zohary, D. Unconscious selection and the evolution of domesticated plants. Econ. Bot. 2004, 58, 5–10. [Google Scholar] [CrossRef]
- Altman, A.; Shennan, S.; Odling-Smee, J. Ornamental plant domestication by aesthetics-driven human cultural niche construction. Trends Plant Sci. 2022, 27, 124–138. [Google Scholar] [CrossRef]
- Guaglianone, E.R. Sinopsis de las especies de Ipheion Raf. y Nothoscordum Kunth (Liliáceas) de Entre Rios y regiones vecinas. Darwiniana 1972, 17, 159–240. [Google Scholar]
- Sassone, A.B.; Giussani, L.M.; Arroyo-Leuenberger, S. Lectotypification and correct author citation of Ipheion uniflorum (Amaryllidaceae), with a new synonym. Ann. Bot. Fenn. 2017, 3847, 99–103. [Google Scholar] [CrossRef] [Green Version]
- Castillo, J.A. Notes on the cultivation of South American bulbous plants. Herbertia 1986, 42, 58–70. [Google Scholar]
- Howard, T. Bulbs for Warm Climates; University of Texas Press: Austin, TX, USA, 2001. [Google Scholar]
- Stearn, W.T. The Welsh Onion and the Ever-Ready Onion; The Gardeners’ Chronicle: London, UK, 1943. [Google Scholar]
- Turrill, W.B. Ipheion uniflorum . Bot. Mag. 1952, 162, 185. [Google Scholar]
- Hurrell, J.A. (Ed.) Alliaceae. In Flora Rioplatense; LOLA: Buenos Aires, Argentina, 2009; Volume 3, pp. 35–80. [Google Scholar]
- Sassone, A.B.; Giussani, L.M.; Guaglianone, E.R. Multivariate studies of Ipheion (Amaryllidaceae, Allioideae) and related genera. Plant Syst. Evol. 2013, 299, 1561–1575. [Google Scholar] [CrossRef]
- Howard-Borjas, P. Women in the Plant World: The Significance of Women and Gender Bias for Biodiversity Conservation; Wageningen Universiteit: Wageningen, The Netherlands, 2001. [Google Scholar]
- Pellicer, J.; Hidalgo, O.; Walker, J.; Chase, M.W.; Christenhusz, M.J.M.; Shackelford, G.; Leitch, I.J.; Fay, M.F. Genome size dynamics in tribe Gilliesieae (Amaryllidaceae, subfamily Allioideae) in the context of polyploidy and unusual incidence of Robertsonian translocations. Bot. J. Linn. Soc. 2017, 184, 16–31. [Google Scholar] [CrossRef]
- Sassone, A.B.; Hojsgaard, D.H.; Giussani, L.M.; Brassac, J.; Blattner, F.R. Genomic, karyological and morphological changes of South American garlics (Ipheion) provide insights into mechanisms of speciation in the Pampean region. Mol. Ecol. 2021, 30, 3716–3729. [Google Scholar] [CrossRef]
- Denham, T.P.; Iriarte, J.; Vrydaghs, L. Selection, cultivation and reproductive isolation: A reconsideration of the morphological and molecular signals of domestication. In Rethinking Agriculture. Archaeological and Ethnoarchaeological Perspectives; Denham, T.P., Iriarte, J., Vrydaghs, L., Eds.; Routledge: New York, NY, USA, 2007; pp. 36–49. [Google Scholar]
- GBIF.org, GBIF Occurrence. Available online: https://doi.org/10.15468/dl.epcmy9 (accessed on 26 December 2021).
- Bivand, R.S.; Lewin-Koh, N. Maptools: Tools for Reading and Handling Spatial Objects. R Package, Ver. 0.8-36. Available online: https://rdrr.io/rforge/maptools/ (accessed on 27 December 2021).
- Ginestet, C. ggplot2: Elegant graphics for data analysis. J. R. Stat. Soc. Ser. A 2011, 174, 245–246. [Google Scholar] [CrossRef]
- Romero Zarco, C.M. A new method for estimating karyotype asymmetry. Taxon 1986, 35, 526–530. [Google Scholar] [CrossRef]
- Huziwara, Y. Karyotype analysis in some genera of Compositae. VIII. Further studies on the chromosomes of Aster. Am. J. Bot. 1962, 49, 116–119. [Google Scholar] [CrossRef]
- Arano, H. Cytological studied in subfamily Carduoideae (Compositae) of Japan IX. Bot. Mag. 1963, 76, 32–39. [Google Scholar] [CrossRef]
- Greilhuber, J.; Speta, F. C-banded karyotypes in the Scilla hohenackeri group, S. persica, and Puschkinia (Liliaceae). Plant Syst. Evol. 1976, 126, 149–188. [Google Scholar] [CrossRef]
- Stebbins, G.L. Chromosomal Evolution in Higher Plants; Edward Arnold Ltd.: London, UK, 1971. [Google Scholar]
- Gompert, Z.; Mock, K.E. Detection of Individual ploidy levels with genotyping-by-sequencing (GBS) analysis. Mol. Ecol. Resour. 2017, 17, 1156–1167. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A.R.; Overcast, I. ipyrad: Interactive assembly and analysis of RADseq datasets. Bioinformatics 2020, 36, 2592–2594. [Google Scholar] [CrossRef] [PubMed]
- Eaton, D.A.R.; Spriggs, E.L.; Park, B.; Donoghue, M.J. Misconceptions on missing data in RAD-seq phylogenetics with a deep-scale example from flowering plants. Syst. Biol. 2017, 66, 399–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gargiulo, R.; Kull, T.; Fay, M.F. Effective double-digest RAD sequencing and genotyping despite large genome size. Mol. Ecol. Resour. 2021, 21, 1037–1055. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [Green Version]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [Green Version]
- Bryant, D.; Moulton, V. Neighbor-Net: An agglomerative method for the construction of phylogenetic Networks. Mol. Biol. Evol. 2004, 21, 255–265. [Google Scholar] [CrossRef]
- Pickrell, J.K.; Pritchard, J.K. Inference of population splits and mixtures from genome- wide allele frequency data. PLoS Genet. 2012, 8, e1002967. [Google Scholar] [CrossRef] [Green Version]
- Goudet, J. HIERFSTAT, a package for R to compute and test hierarchical F-statistics. Mol. Ecol. Notes 2005, 5, 184–186. [Google Scholar] [CrossRef] [Green Version]
- Unmack, P.J.; Berry, O.F.; Georges, A.; Gruber, B. DARTR: An R package to facilitate analysis of SNP data generated from reduced representation genome sequencing. Mol. Ecol. Resour. 2018, 18, 691–699. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-statistics for the analysis of population structure. Evolution 1984, 38, 1358. [Google Scholar] [CrossRef] [PubMed]
- Baldi, G.; Guerschman, J.P.; Paruelo, J.M. Characterizing fragmentation in temperate South America grasslands. Agric. Ecosyst. Environ. 2006, 116, 197–208. [Google Scholar] [CrossRef]
- Yu, M.; Liu, Z.H.; Yang, B.; Chen, H.; Zhang, H.; Hou, D.B. The contribution of photosynthesis traits and plant height components to plant height in wheat at the individual quantitative trait locus level. Sci. Rep. 2020, 10, 12261. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, S.; Simmonds, J.; Leverington, M.; Wang, Y.; Fish, L.; Sayers, L.; Alibert, L.; Orford, S.; Wingen, L.; Snape, J. Meta-QTL analysis of the genetic control of crop height in elite European winter wheat germplasm. Mol. Breed. 2012, 29, 159–171. [Google Scholar] [CrossRef]
- FAO and ITPS. A Main Report. Chapter 12, Regional assessment of soil changes in Latin America and the Caribbean. In Status of the World’s Soil Resources; Food and Agriculture Organization of the United Nations and Intergovernmental Technical Panel on Soils: Rome, Italy, 2015; pp. 364–398. ISBN 9789251090046. [Google Scholar]
- Gaut, B.S.; Seymour, D.K.; Liu, Q.; Zhou, Y. Demography and its effects on genomic variation in crop domestication. Nat. Plants 2018, 4, 512–520. [Google Scholar] [CrossRef]
- Wright, S.I.; Bi, I.V.; Schroeder, S.G.; Yamasaki, M.; Doebley, J.F.; McMullen, M.D.; Gaut, B.S. The effects of artificial selection on the maize genome. Science 2005, 308, 1310–1314. [Google Scholar] [CrossRef]
- Wisz, M.S.; Pottier, J.; Kissling, W.D.; Pellissier, L.; Lenoir, J.; Damgaard, C.F.; Dormann, C.F.; Forchhammer, M.C.; Grytnes, J.A.; Guisan, A.; et al. The role of biotic interactions in shaping distributions and realised assemblages of species: Implications for species distribution modelling. Biol. Rev. 2013, 88, 15–30. [Google Scholar] [CrossRef] [Green Version]
- Berry, J.C.; Fahlgren, N.; Pokorny, A.A.; Bart, R.S.; Veley, K.M. An automated, high-throughput method for standardizing image color profiles to improve image-based plant phenotyping. PeerJ 2018, 6, e5727. [Google Scholar] [CrossRef] [Green Version]
- Cuiid, H.; Zhang, Y.; Shi, X.; Gong, F.; Xiong, X.; Kang, X.; Xing, G.; Li, S. The numerical classification and grading standards of daylily (Hemerocallis) flower color. PLoS ONE 2019, 14, e0216460. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Population | Sample | Origin | Ploidy | Karyotype Formula | 2Cx (pg.) | |
|---|---|---|---|---|---|---|
| Cytology | gbs2ploidy | |||||
| 513_uniAZ | AS203 | Natural | 2n = 12 | 2x | 1SM + 5A | 9.6 |
| 540_uniAZ | AS224 | Natural | 2n = 12 | 2x | 1SM + 5A | --- |
| 553_uniTA | AS235 | Natural | 2n = 12 | 2x | 1SM + 5A | 9.65 |
| 555_uniTA | AS98 | Natural | 2n = 12 | 2x | 1SM + 5A | 10.05 |
| S51_uniSCa | AS236 | Natural | 2n = 12 | 2x | 1SM + 5A | --- |
| Vil_uniBB | AS135 | Natural | 2n = 12 | 2x | 1SM + 5A | --- |
| SSF1 | AS182 | Cultivated | 2n = 12 | 2x | 1SM + 5A | 9.6 |
| SSF2 | AS183 | Cultivated | 2n = 24 | 4x | 1SM + 5A | 18.8 |
| SSF3 | AS247 | Cultivated | 2n = 12 | 2x | 1SM + 5A | 9.75 |
| SSF4 | AS266 | Cultivated | --- | 2x | --- | --- |
| SSF5 | AS267 | Cultivated | --- | 2x | --- | --- |
| Region | TCL | c | c max | c min | i | TF% | Ask% | Syi | A1 | A2 | R | r |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Centre | 111.7 ± 9.9 | 9.33 ± 0.8 | 11.1 | 7.1 | 24.1 | 8.9 | 91.1 | 9.8 | 0.99 | 0.093 | 3.4 | 4.0 |
| South | 121 ± 9.6 | 10.1 ± 0.8 | 11.6 | 8 | 24.9 | 9.1 | 90.9 | 10 | 0.99 | 0.076 | 4.0 | 4.2 |
| North | Center | South | |
|---|---|---|---|
| Center | 0.154 (0.147–0.147) | - | - |
| South | 0.322 (0.314–0.314) | 0.308 (0.302–0.314) | - |
| Cultivated | 0.139 (0.128–0.149) | 0.249 (0.242–0.248) | 0.485 (0.477–0.493) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sassone, A.B.; Blattner, F.R.; Giussani, L.M.; Hojsgaard, D.H. First Glimpse on Spring Starflower Domestication. Genes 2022, 13, 243. https://doi.org/10.3390/genes13020243
Sassone AB, Blattner FR, Giussani LM, Hojsgaard DH. First Glimpse on Spring Starflower Domestication. Genes. 2022; 13(2):243. https://doi.org/10.3390/genes13020243
Chicago/Turabian StyleSassone, Agostina B., Frank R. Blattner, Liliana M. Giussani, and Diego H. Hojsgaard. 2022. "First Glimpse on Spring Starflower Domestication" Genes 13, no. 2: 243. https://doi.org/10.3390/genes13020243
APA StyleSassone, A. B., Blattner, F. R., Giussani, L. M., & Hojsgaard, D. H. (2022). First Glimpse on Spring Starflower Domestication. Genes, 13(2), 243. https://doi.org/10.3390/genes13020243