
Combining Abilities and Heterotic Patterns among Early Maturing Maize Inbred Lines under Optimal and Striga-Infested Environments

 ,
,  , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Genetic Materials
2.2. Field Experiment
2.3. Data Collection
2.4. DNA Extraction and Genotyping Using SNPs
2.5. Statistical Analysis
2.6. Heterotic Grouping
3. Results
3.1. Analysis of Variance of Phenotypic Traits under Striga Infestation and Optimal Environments
3.2. Proportionate Contributions of Combining Ability Effects of the Inbred Lines
3.3. Estimates of General Combining Ability Effects of Grain Yield and Striga Adaptive Traits of the Maize Inbred Lines
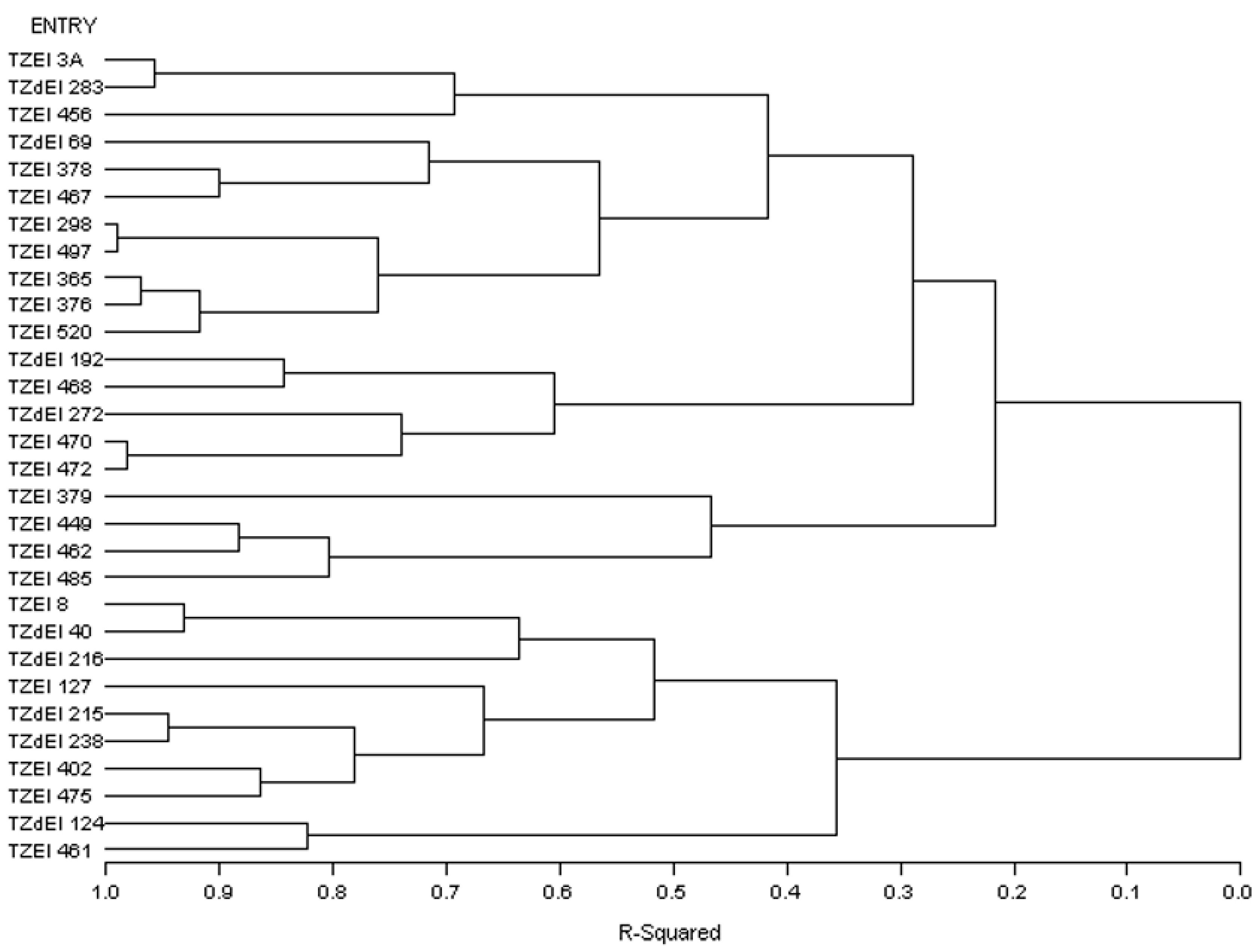
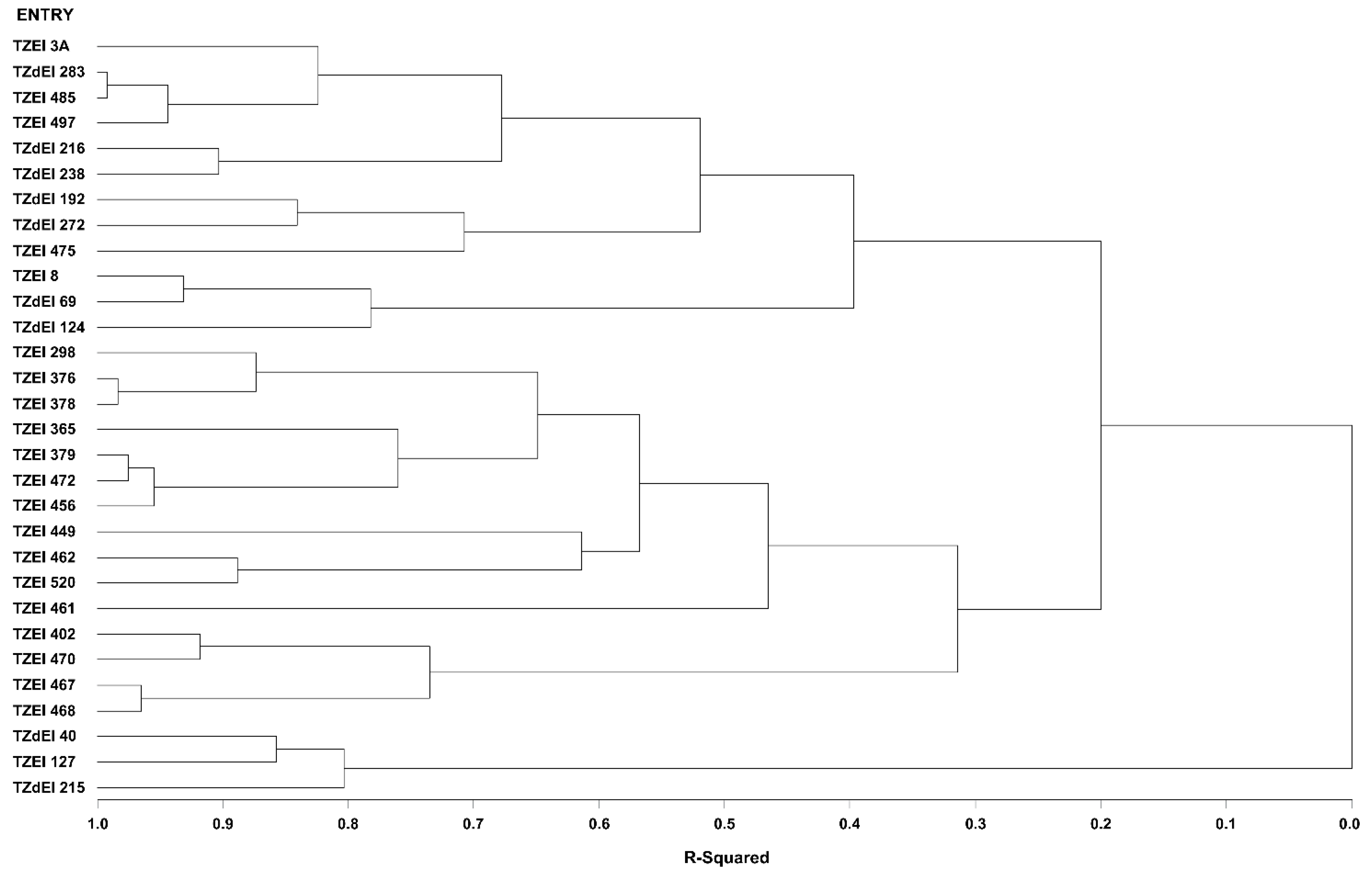
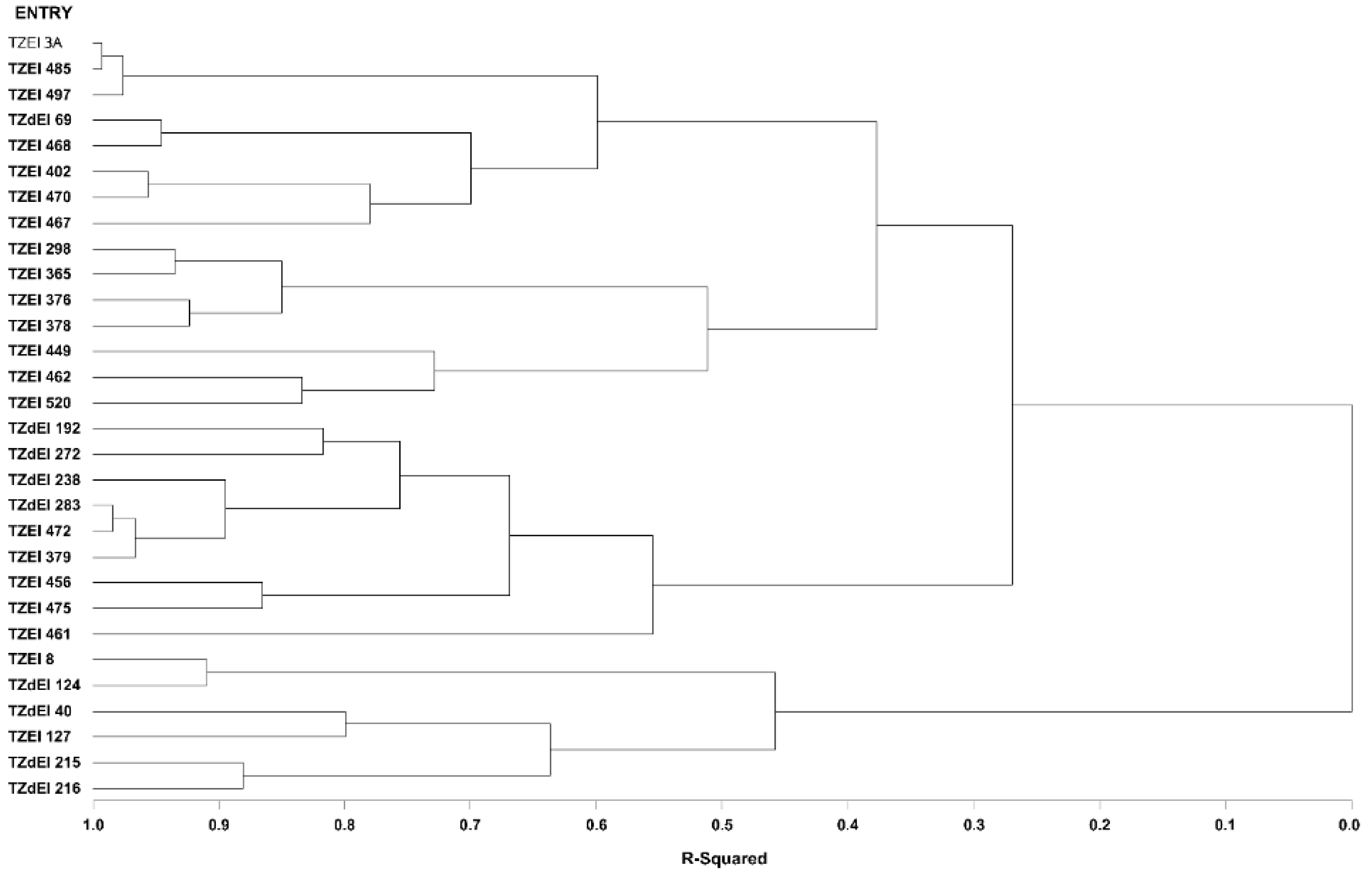
3.4. Heterotic Groupings and Relationships among the Different Heterotic Grouping Methods
3.5. Comparison of Different Methods of Heterotic Grouping under Contrasting Environments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- OECD/FAO. OECD-FAO Agricultural Outlook 2018–2027; OECD Publisher: Paris, France; FAO: Rome, Italy, 2018; 181p. [Google Scholar] [CrossRef]
- Ejeta, G.; Gressel, J. Integrating New Technologies for Striga Control: Towards Ending the Witch-Hunt; World Scientific: Singapore, 2007; 345p. [Google Scholar] [CrossRef]
- Shayanowako, A.T.; Laing, M.; Shimelis, H.; Mwadzingeni, L. Resistance Breeding and Biocontrol of Striga asiatica (L.) Kuntze in Maize: A Review. Acta Agric. Scand. Sect. B Soil Plant Sci. 2018, 68, 110–120. [Google Scholar] [CrossRef]
- Makumbi, D.; Diallo, A.; Kanampiu, F.; Mugo, S.; Karaya, H. Agronomic Performance and Genotype × Environment Interaction of Herbicide-Resistant Maize Varieties in Eastern Africa. Crop Sci. 2015, 55, 540–555. [Google Scholar] [CrossRef]
- Kroschel, J. Parasitic Weeds of the World: Biology and Control. By, C. Parker and C. R. Riches. Wallingford, Oxfordshire: CAB International (1993), pp. 332, £45.00, US£ 85.00 (Americas Only). ISBN 0-85198-873-3. Exp. Agric. 1994, 30, 490. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B. Advances in Genetic Enhancement of Early and Extra-Early Maize for Sub-Saharan Africa; Springer: Berlin/Heidelberg, Germany, 2017; ISBN 9783319648521. [Google Scholar]
- Runo, S.; Macharia, S.; Alakonya, A.; Machuka, J.; Sinha, N.; Scholes, J. Striga Parasitizes Transgenic Hairy Roots of Zea Mays and Provides a Tool for Studying Plant-Plant Interactions. Plant Methods 2012, 8, 20. [Google Scholar] [CrossRef] [PubMed]
- Badu-Apraku, B.; Oyekunle, M.; Fakorede, M.A.B.; Vroh, I.; Akinwale, R.O.; Aderounmu, M. Combining ability, heterotic patterns and genetic diversity of extra-early yellow inbreds under contrasting environments. Euphytica 2013, 192, 413–433. [Google Scholar] [CrossRef]
- Talabi, A.O.; Badu-Apraku, B.; Fakorede, M.A.B. Genetic Variances and Relationship among Traits of an Early Maturing Maize Population under Drought-Stress and Low Nitrogen Environments. Crop Sci. 2017, 57, 681–692. [Google Scholar] [CrossRef]
- Aggarwal, V.D. Research on Cowpea-Striga Resistance at IITA. In Proceedings of the Combating Striga in Africa, Ibadan, Nigeria, 22–24 August 1988; Kim, S.K., Ed.; IITA: Oyo State, Nigeria, 1991; pp. 90–95. [Google Scholar]
- Kwon, K.S. Genetics of Maize Tolerance of Striga hermonthica. Crop Sci. 1994, 34, 900–907. [Google Scholar] [CrossRef]
- Betrán, F.J.; Beck, D.; Bänziger, M.; Edmeades, G.O. Genetic Analysis of Inbred and Hybrid Grain Yield under Stress and Nonstress Environments in Tropical Maize. Crop Sci. 2003, 43, 807–817. [Google Scholar] [CrossRef]
- Abu, P.; Badu-Apraku, B.; Ifie, B.E.; Tongoona, P.; Ribeiro, P.F.; Obeng-Bio, E.; Offei, S.K. Genetics of extra-early-maturing yellow and orange quality protein maize inbreds and derived hybrids under low soil nitrogen and Striga infestation. Crop Sci. 2021, 61, 1052–1072. [Google Scholar] [CrossRef] [PubMed]
- Oyekale, S.A.; Badu-Apraku, B.; Adetimirin, V.O. Combining Ability of Extra-early Biofortified Maize Inbreds under Striga Infestation and Low Soil Nitrogen. Crop Sci. 2020, 60, 1925–1945. [Google Scholar] [CrossRef]
- Annor, B.; Badu-Apraku, B.; Nyadanu, D.; Akromah, R.; Fakorede, M.A.B. Testcross performance and combining ability of early maturing maize inbreds under multiple-stress environments. Sci. Rep. 2019, 9, 13809. [Google Scholar] [CrossRef] [PubMed]
- Barata, C.; Carena, M.J. Classification of North Dakota Maize Inbred Lines into Heterotic Groups Based on Molecular and Testcross Data. Euphytica 2006, 151, 339–349. [Google Scholar] [CrossRef]
- Suwarno, W.B.; Pixley, K.V.; Palacios-Rojas, N.; Kaeppler, S.M.; Babu, R. Formation of Heterotic Groups and Understanding Genetic Effects in a Provitamin a Biofortified Maize Breeding Program. Crop Sci. 2014, 54, 14–24. [Google Scholar] [CrossRef]
- Yan, W.; Kang, M.S. GGE Biplot Analysis: A Graphical Tool for Breeders, Geneticists, and Agronomists; CRC Press: Boca Raton, FL, USA, 2002. [Google Scholar] [CrossRef]
- Carena, M.J.; Hallauer, A.R.; Miranda Filho, J.B. Quantitative Genetics in Maize Breeding, 2nd ed.; Prohens, J., Nuez, F., Carena, M.J., Eds.; Springer: Dordrecht, The Netherlands, 2010; ISBN 978-1-4419-0765-3. [Google Scholar]
- Fan, X.M.; Chen, H.M.; Tan, J.; Xu, C.X.; Zhang, Y.M.; Huang, Y.X.; Kang, M.S. A New Maize Heterotic Pattern between Temperate and Tropical Germplasms. Agron. J. 2008, 100, 917–923. [Google Scholar] [CrossRef]
- Fan, X.M.; Zhang, Y.M.; Yao, W.H.; Chen, H.M.; Tan, J.; Xu, C.X.; Han, X.L.; Luo, L.M.; Kang, M.S. Classifying Maize Inbred Lines into Heterotic Groups Using a Factorial Mating Design. Agron. J. 2009, 101, 106–112. [Google Scholar] [CrossRef]
- Fan, X.M.; Tan, J.; Yang, J.Y.; Chen, H.M. Combining Ability and Heterotic Grouping of Ten Temperate, Subtropical and Tropical Quality Protein Maize Inbreds. Maydica 2004, 49, 267–272. [Google Scholar]
- Akinwale, R.O.; Badu-Apraku, B.; Fakorede, M.A.B.; Vroh-Bi, I. Heterotic Grouping of Tropical Early-Maturing Maize Inbred Lines Based on Combining Ability in Striga-Infested and Striga-Free Environments and the Use of SSR Markers for Genotyping. Field Crops Res. 2014, 156, 48–62. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Yallou, C.G.; Oyekunle, M. Genetic Gains from Selection for High Grain Yield and Striga Resistance in Early Maturing Maize Cultivars of Three Breeding Periods under Striga-Infested and Striga-Free Environments. Field Crops Res. 2013, 147, 54–67. [Google Scholar] [CrossRef]
- Balestre, M.; Von Pinho, R.G.; Souza, J.C.; Lima, J.L. Comparison of Maize Similarity and Dissimilarity Genetic Coefficients Based on Microsatellite Markers. Genet. Mol. Res. 2008, 7, 695–705. [Google Scholar] [CrossRef]
- Lanza, L.L.B.; De Souza, C.L.; Ottoboni, L.M.M.; Vieira, M.L.C.; De Souza, A.P. Genetic Distance of Inbred Lines and Prediction of Maize Single-Cross Performance Using RAPD Markers. Theor. Appl. Genet. 1997, 94, 1023–1030. [Google Scholar] [CrossRef]
- Reif, J.C.; Melchinger, A.E.; Xia, X.C.; Warburton, M.L.; Hoisington, D.A.; Vasal, S.K.; Srinivasan, G.; Bohn, M.; Frisch, M. Genetic Distance Based on Simple Sequence Repeats and Heterosis in Tropical Maize Populations. Crop Sci. 2003, 43, 1275–1282. [Google Scholar] [CrossRef]
- Hamblin, M.T.; Warburton, M.L.; Buckler, E.S. Empirical Comparison of Simple Sequence Repeats and Single Nucleotide Polymorphisms in Assesment of Maize Diversity and Relatedness. PLoS ONE 2007, 2, e1367. [Google Scholar] [CrossRef] [PubMed]
- Semagn, K.; Magorokosho, C.; Ogugo, V.; Makumbi, D.; Warburton, M.L. Genetic Relationships and Structure among Open-Pollinated Maize Varieties Adapted to Eastern and Southern Africa Using Microsatellite Markers. Mol. Breed. 2014, 34, 1423–1435. [Google Scholar] [CrossRef]
- Semagn, K.; Magorokosho, C.; Vivek, B.S.; Makumbi, D.; Beyene, Y.; Mugo, S.; Prasanna, B.M.; Warburton, M.L. Molecular Characterization of Diverse CIMMYT Maize Inbred Lines from Eastern and Southern Africa Using Single Nucleotide Polymorphic Markers. BMC Genom. 2012, 13, 113. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fakorede, M.A.B.; Gedil, M.; Talabi, A.O.; Annor, B.; Oyekunle, M.; Akinwale, R.O.; Fasanmade, T.Y.; Akaogu, I.C.; Aderounmu, M. Heterotic Responses among Crosses of IITA and CIMMYT Early White Maize Inbred Lines under Multiple Stress Environments. Euphytica 2015, 206, 245–262. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Annor, B.; Oyekunle, M.; Akinwale, R.O.; Fakorede, M.A.B.; Talabi, A.O.; Akaogu, I.C.; Melaku, G.; Fasanmade, Y. Grouping of Early Maturing Quality Protein Maize Inbreds Based on SNP Markers and Combining Ability under Multiple Environments. Field Crops Res. 2015, 183, 169–183. [Google Scholar] [CrossRef]
- Comstock, R.E.; Robinson, H.F. The Components of Genetic Variance in Populations of Biparental Progenies and Their Use in Estimating the Average Degree of Dominance. Biometrics 1948, 4, 254. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Oyekunle, M. Genetic Analysis of Grain Yield and other Traits of Extra-Early Yellow Maize Inbreds and Hybrid Performance under Contrasting Environments. Field Crops Res. 2012, 129, 99–110. [Google Scholar] [CrossRef]
- Jaccoud, D.; Peng, K.; Feinstein, D.; Kilian, A. Diversity Arrays: A Solid State Technology for Sequence Information Independent Genotyping. Nucleic Acids Res. 2001, 29, 25. [Google Scholar] [CrossRef]
- SAS. Base SAS 9.1.3 Procedures Guide; SAS Documentation; SAS Publisher: Cary, NC, USA, 2006; ISBN 9781590477540. [Google Scholar]
- Cochran, W.G. Experimental Designs; William, G., Cox, G.M., Eds.; CRC: Boca Raton, FL, USA, 1992; p. 611. [Google Scholar]
- Hallauer, A.R.; Miranda Filho, J.B. Quantitative Genetics in Maize Breeding; Springer: Berlin/Heidelberg, Germany, 1988; p. 468. [Google Scholar]
- Satterthwaite, F.E. An Approximate Distribution of Estimates of Variance Components. Biom. Bull. 1946, 2, 110. [Google Scholar] [CrossRef]
- Singh, J.P. Biometrical Techniques in Plant Breeding; Kalyani Publishers: New Delhi, India, 1993. [Google Scholar]
- Jaccard, P. Nouvelles Recherches Sur La Distribuition Florale. Bull. Sociéetée Vandoise Des Sci. Nat. 1908, 44, 223–270. [Google Scholar] [CrossRef]
- Perrier and Jacquemoud-Collet DARwin Software. Available online: http://http//darwin.cirad.fr (accessed on 18 February 2018).
- Adu, G.B.; Badu-Apraku, B.; Akromah, R.; Haruna, A.; Amegbor, I.K.; Amadu, M.K. Grain Yield and Stability of Early-Maturing Single-Cross Hybrids of Maize across Contrasting Environments. J. Crop Improv. 2019, 33, 776–796. [Google Scholar] [CrossRef]
- Badu-Apraku, B.; Fontem, L.A.; Akinwale, R.O.; Oyekunle, M. Biplot Analysis of Diallel Crosses of Early Maturing Tropical Yellow Maize Inbreds in Stress and Nonstress Environments. Crop Sci. 2011, 51, 173–188. [Google Scholar] [CrossRef]
- Ifie, B.E.; Badu-Apraku, B.; Gracen, V.; Danquah, E.Y. Genetic Analysis of Grain Yield of IITA and CIMMYT Early-Maturing Maize Inbreds under Striga-Infested and Low–Soil-Nitrogen Environments. Crop Sci. 2015, 55, 610–623. [Google Scholar] [CrossRef]
- Musila, R.N.; Diallo, A.O.; Makumbi, D.; Njoroge, K. Combining Ability of Early-Maturing Quality Protein Maize Inbred Lines Adapted to Eastern Africa. Field Crops Res. 2010, 119, 231–237. [Google Scholar] [CrossRef]
- Konate, L.; Baffour, B.A.; Traore, D. Combining Ability and Heterotic Grouping of Early Maturing Provitamin a Maize Inbreds across Striga Infested and Optimal Growing Environments. J. Agric. Environ. Int. Dev. 2017, 111, 157–173. [Google Scholar] [CrossRef]
- Baker, R.J. Issues in Diallel Analysis. Crop Sci. 1978, 18, 533–536. [Google Scholar] [CrossRef]
- Gethi, J.G.; Smith, M.E. Genetic Responses of Single Crosses of Maize to Striga hermonthica (Del.) Benth. and Striga asiatica (L.) Kuntze. Crop Sci. 2004, 44, 2068–2077. [Google Scholar] [CrossRef]
- Yallou, C.G.; Menkir, A.; Adetimirin, V.O.; Kling, J.G. Combining Ability of Maize Inbred Lines Containing Genes from Zea Diploperennis for Resistance to Striga hermonthica (Del.) Benth. Plant Breed. 2009, 128, 143–148. [Google Scholar] [CrossRef]
- Mulualem, T.; Abate, M. Heterotic Response in Major Cereals and Vegetable Crops. Int. J. Plant Breed. Genet. 2016, 10, 69–78. [Google Scholar] [CrossRef][Green Version]
- Beckett, R.C.; Ludwick, T.M.; Rader, E.R.; Hines, H.C.; Pearson, R. Specific and General Combining Abilities for Production and Reproduction among Lines of Holstein Cattle. J. Dairy Sci. 1979, 62, 613–620. [Google Scholar] [CrossRef]
- Hu, L.; Huang, T.; Liu, X.J.; Cai, Y.D. Predicting Protein Phenotypes Based on Protein-Protein Interaction Network. PLoS ONE 2011, 6, e17668. [Google Scholar] [CrossRef] [PubMed]
- Menkir, A.; Akintunde, A.O. Evaluation of the Performance of Maize Hybrids—Improved Open-Pollinated and Farmers’ Local Varieties under Well-Watered and Drought Stress Conditions [Zea mays, L.—Ghana–Nigeria]. Maydica 2001, 46, 227–238. [Google Scholar]
- Derera, J.; Tongoona, P.; Vivek, B.S.; Laing, M.D. Gene Action Controlling Grain Yield and Secondary Traits in Southern African Maize Hybrids under Drought and Non-Drought Environments. Euphytica 2008, 162, 411–422. [Google Scholar] [CrossRef]
- Oyekunle, M.; Badu-Apraku, B. Genetic Analysis of Grain Yield and other Traits of Early-Maturing Maize Inbreds under Drought and Well-Watered Conditions. J. Agron. Crop Sci. 2014, 200, 92–107. [Google Scholar] [CrossRef]
- Adu, G.B.; Badu-Apraku, B.; Akromah, R. Strategies for Selecting Early Maturing Maize Inbred Lines for Hybrid Production under Low Soil Nitrogen and Striga Infestation. Agronomy 2021, 11, 1309. [Google Scholar] [CrossRef]
- Lee, M. DNA Markers and Plant Breeding Programs. Adv. Agron. 1995, 55, 265–344. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Name | Set | Endosperm Color | Reaction to Striga Infestation |
|---|---|---|---|---|
| 1 | TZdEI 283 | A | White | Tolerant |
| 2 | TZdEI 216 | A | White | Tolerant |
| 3 | TZEI 378 | A | White | Tolerant |
| 4 | TZEI 379 | A | White | Tolerant |
| 5 | TZdEI 69 | A | White | Susceptible |
| 6 | TZdEI 272 | B | White | Tolerant |
| 7 | TZdEI 215 | B | White | Tolerant |
| 8 | TZEI 402 | B | White | Tolerant |
| 9 | TZEI 3A | B | White | Tolerant |
| 10 | TZEI 298 | B | White | Tolerant |
| 11 | TZdEI 238 | C | White | Tolerant |
| 12 | TZEI 365 | C | White | Tolerant |
| 13 | TZEI 376 | C | White | Tolerant |
| 14 | TZdEI 192 | C | White | Tolerant |
| 15 | TZdEI 124 | C | White | Tolerant |
| 16 | TZEI 468 | D | Yellow | Tolerant |
| 17 | TZEI 485 | D | Yellow | Tolerant |
| 18 | TZEI 461 | D | Yellow | Tolerant |
| 19 | TZdEI 40 | D | Yellow | Tolerant |
| 20 | TZEI 520 | D | Yellow | Tolerant |
| 21 | TZEI 472 | E | Yellow | Tolerant |
| 22 | TZEI 456 | E | Yellow | Tolerant |
| 23 | TZEI 462 | E | Yellow | Susceptible |
| 24 | TZEI 467 | E | Yellow | Tolerant |
| 25 | TZEI 127 | E | Yellow | Tolerant |
| 26 | TZEI 475 | F | Yellow | Tolerant |
| 27 | TZEI 8 | F | Yellow | Tolerant |
| 28 | TZEI 470 | F | Yellow | Tolerant |
| 29 | TZEI 449 | F | Yellow | Susceptible |
| 30 | TZEI 497 | F | Yellow | Tolerant |
| Source of Variation | DF | Grain Yield | Days to Anthesis | Days to Silking | Anthesis–Silking Interval | Plant Height | Ear Height | Root Lodging | Stalk Lodging | Plant Aspect | Ear Aspect | Ear per Plant | Striga Damage (8 WAP) | Striga Damage (10 WAP) | Striga Count (8 WAP) | Striga Count (10 WAP) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Environment (E) | 3 | 45,253,715.30 ** | 2007.65 ** | 1827.14 ** | 20.69 ** | 69452.55 ** | 4624.55 ** | 436.32 ** | 316.83 ** | 771.40 ** | 406.07 ** | 9.21 ** | 1461.83 ** | 1778.24 ** | 0.27 | 64.41 ** |
| Set | 5 | 2,537,531.50 ** | 21.03 * | 27.40 * | 0.33 | 6836.23 ** | 760.45 ** | 0.59 | 1.78 * | 11.68 ** | 12.98 ** | 0.36 ** | 6.79 ** | 7.23 ** | 4.64 * | 3.37 * |
| E*Set | 15 | 569,442.60 * | 10.26 * | 11.90 * | 0.23 | 560.48 * | 108.26 | 1.09 * | 0.65 | 1.95 * | 2.03 * | 0.06 | 2.28 ** | 2.30 * | 1.6 | 1.02 |
| REP (E*Set) | 20 | 549,732.80 * | 5.10 | 4.29 | 0.21 | 120.23 | 46.63 | 0.59 | 0.72 | 0.82 | 0.82 | 0.03 | 0.52 | 0.98 | 0.91 | 0.84 |
| Block (E*REP) | 96 | 772,888.90 ** | 10.23 ** | 12.87 ** | 0.28* | 427.12 ** | 115.41 ** | 0.92 ** | 0.99 * | 1.57 ** | 2.11 ** | 0.05 * | 2.23 ** | 3.41 ** | 2.02 * | 1.15 * |
| Hybrid | 155 | 693,210.70 ** | 14.28 ** | 11.36 ** | 0.21 | 879.49 ** | 165.05 ** | 0.78 ** | 0.85 * | 2.26 ** | 1.86 ** | 0.07 ** | 1.37 ** | 1.62 ** | 2.77 * | 2.35 ** |
| GCA-male (Set) | 24 | 741,937.20* | 23.49 ** | 19.48 ** | 0.13 | 930.75 ** | 89.23 | 0.75 * | 0.89 | 2.40 ** | 1.96 * | 0.08 * | 1.83 ** | 1.40 * | 3.37 * | 2.99 ** |
| GCA-female (Set) | 24 | 1,002,635.70 ** | 18.41 ** | 11.60 * | 0.26 | 962.94 ** | 212.62 ** | 1.10 * | 0.88 | 1.91 * | 1.87 * | 0.06 * | 2.02 ** | 2.69 ** | 2.23 * | 1.84 * |
| SCA (Set) | 96 | 541,397.90 ** | 10.79 ** | 9.23 * | 0.22 | 501.63 ** | 137.13 ** | 0.72 ** | 0.81 | 1.75 ** | 1.46 * | 0.05 * | 0.89* | 1.19 * | 2.32 * | 1.98 ** |
| Hybrid*E | 465 | 461,059.90 * | 6.21 * | 9.28 * | 0.21 | 264.67 * | 64.34 | 0.67 ** | 0.74 | 1.52 ** | 1.33 ** | 0.05 * | 1.04 ** | 1.04 * | 1.29 | 0.82 |
| GCA-male (Set)*E | 72 | 376,159.1 | 5.81 | 9.07 * | 0.21 | 146.91 | 43.73 | 0.55 | 0.54 | 1.30 * | 1.16 | 0.04 | 1.25 ** | 1.12 * | 0.94 | 0.82 |
| GCA-female (Set)*E | 72 | 608,112.80 * | 6.63 * | 9.71 * | 0.18 | 260.05 | 53.88 | 0.73 * | 0.81 | 2.09 ** | 1.53 * | 0.04 | 1.47 ** | 1.23 * | 1.45 | 0.74 |
| SCA (Set)*E | 288 | 442,171.40 * | 5.96 * | 8.41 * | 0.23 | 266.31 * | 67.06 | 0.67 ** | 0.79 * | 1.32 ** | 1.18 * | 0.05 * | 0.83 * | 0.88 | 1.26 | 0.8 |
| Error | 480 | 330,203.40 | 4.85 | 6.92 | 0.22 | 200.50 | 64.60 | 0.44 | 0.64 | 0.76 | 0.91 | 0.04 | 0.62 | 0.78 | 1.40 | 0.84 |
| Source of Variation | DF | Grain Yield | Days to Anthesis | Days to Silking | Anthesis–Silking Interval | Plant Height | Ear Height | Root Lodging | Stalk Lodging | Husk Cover | Plant Aspect | Ear Aspect | Ear per Plant |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Optimal Environment | |||||||||||||
| Environment (E) | 3 | 115,159,309.2 ** | 4466.41 ** | 2588.8 ** | 78.71 ** | 24,827.87 ** | 4028.33 ** | 45.29 ** | 121.96 ** | 1316.51 ** | 830.88 ** | 786.93 ** | 1.43 ** |
| Set | 5 | 9,415,440.8 ** | 35.41 ** | 39.41 ** | 0.06 | 26,721.29 ** | 1615.15 ** | 2.48 ** | 0.95 | 12.19 ** | 11.75 ** | 8.01 ** | 0.17 * |
| E*Set | 15 | 2,016,314.1 * | 8.74 ** | 8.50 ** | 0.03 | 715.28 ** | 130.08 ** | 1.69 ** | 1.23 * | 1.2 ** | 0.87 ** | 0.49 | 0.12 * |
| REP (E*Set) | 20 | 847,607.7 | 3.07 | 3.17 | 0.04 | 297.9 * | 52.55 | 0.27 | 0.77 | 0.48 | 0.41 | 0.18 | 0.13 * |
| Block (E*REP) | 96 | 1,526,045 ** | 6.94 ** | 7.40 ** | 0.04 | 417.74 ** | 141.66 ** | 0.89 ** | 0.92 * | 0.41 * | 0.97 ** | 0.66 ** | 0.07 |
| Hybrid | 155 | 3,350,625.4 ** | 11.68 ** | 13.43 ** | 0.04 | 1914.26 ** | 292.32 ** | 1.15 ** | 1.21 ** | 1.18 ** | 1.59 ** | 1.03 ** | 0.16 ** |
| GCA-male (Set) | 24 | 3,449,292.5 ** | 16.70 * | 19.33 ** | 0.04 | 1665.97 ** | 271.18 ** | 1.55 | 1.76 * | 0.99 ** | 0.93 | 0.98 * | 0.22 ** |
| GCA-female (Set) | 24 | 4,688,082.7 ** | 17.71 ** | 21.02 ** | 0.05 | 1797.22 ** | 484.18 ** | 0.9 | 1.53 * | 0.98 | 1.73 ** | 1.25 * | 0.23 * |
| SCA (Set) | 96 | 2,481,527.2 ** | 6.87 ** | 7.87 ** | 0.04 | 549.83 ** | 131.25 * | 0.97 ** | 1.06 | 0.62 | 1.09 ** | 0.59 * | 0.12 |
| Hybrid*E | 465 | 1,584,497.6 ** | 5.18 ** | 6.27 ** | 0.04 | 369.54 ** | 107.02 ** | 0.91 ** | 0.93 ** | 0.65 ** | 0.75 ** | 0.47 ** | 0.12 ** |
| GCA-male (Set)*E | 72 | 1,349,720.6 * | 6.80 ** | 7.86 ** | 0.03 | 335.27 ** | 108.71 ** | 1.35 ** | 0.91 * | 0.7 ** | 0.88 ** | 0.58 ** | 0.10 * |
| GCA-female (Set)*E | 72 | 1,703,657.4 ** | 6.03 ** | 6.68 ** | 0.05 | 390.85 ** | 138.28 ** | 0.76 | 0.84 | 0.87 ** | 0.7 ** | 0.45 ** | 0.14 ** |
| SCA (Set)*E | 288 | 1,521,403.7 ** | 4.04 ** | 5.01 ** | 0.04 | 344.79 ** | 91.04 ** | 0.79 ** | 0.93 ** | 0.53 ** | 0.67 ** | 0.43 ** | 0.12 ** |
| Error | 480 | 975,007 | 2.78 | 3.15 | 0.04 | 180.54 | 59.27 | 0.60 | 0.68 | 0.31 | 0.38 | 0.30 | 0.07 |
| Across Research Environments | |||||||||||||
| Environment (E) | 7 | 205,147,091.00 ** | 6529.19 ** | 3624.53 ** | 46.14 ** | 75,261.48 ** | 32,464.35 ** | 218.13 ** | 209.89 ** | 1080.29 ** | 691.02 ** | 566.25 ** | 7.74 ** |
| SET | 5 | 10,372,430.00 ** | 52.09 ** | 51.58 ** | 0.2 | 29,822.98 ** | 1847.92 ** | 1.95 ** | 2.07 ** | 6.98 ** | 22.58 ** | 20.12 ** | 0.42 ** |
| E*SET | 35 | 1,333,866.00 ** | 8.76 ** | 10.91 ** | 0.14 | 1083.02 ** | 177.6 ** | 1.32 ** | 0.9 | 1.78 ** | 1.33 ** | 1.21 ** | 0.09 ** |
| REP (E*SET) | 40 | 698,670 | 4.08 | 3.73 | 0.12 | 209.07 | 49.59 | 0.43 | 0.75 | 0.38 | 0.62 | 0.5 | 0.08 * |
| Block (E*REP) | 192 | 1,149,467.00 ** | 8.58 ** | 10.14 ** | 0.16 * | 422.43 ** | 128.54 ** | 0.90 ** | 0.96 ** | 0.66 ** | 1.27 ** | 1.39 ** | 0.06 |
| Hybrid | 155 | 2,748,369.00 ** | 18.80 ** | 17.29 ** | 0.13 | 2271.94 ** | 314.22 ** | 1.24 ** | 1.38 ** | 0.85 ** | 2.43 ** | 2.01 ** | 0.13 ** |
| GCA-Male (Set) | 24 | 3,024,316.00 ** | 32.99 ** | 32.81 ** | 0.08 | 2215.06 ** | 279.84 ** | 1.80 * | 1.67 ** | 0.63 | 2.04 ** | 1.69 * | 0.18 ** |
| GCA-Female (Set) | 24 | 4,296,941.00 ** | 28.29 ** | 25.5 ** | 0.15 | 2312.74 ** | 571.42 ** | 1.28 * | 1.81 ** | 0.9 | 1.95 | 1.97 ** | 0.17 * |
| SCA (Set) | 96 | 1,774,458.00 ** | 11.16 ** | 9.64 ** | 0.13 | 656.27 ** | 145.87 ** | 1.00 * | 1.19 ** | 0.52 | 1.47 ** | 1.3 ** | 0.1 |
| Hybrid*E | 1085 | 1,060,594.00 ** | 5.90 ** | 7.73 ** | 0.13 | 346.13 ** | 93.71 ** | 0.78 ** | 0.81 ** | 0.56 ** | 1.18 ** | 0.9 ** | 0.08 ** |
| GCA Male (Set)*E | 168 | 909,546.00 ** | 6.45 ** | 8.11 ** | 0.12 | 261.2 ** | 770 * | 0.88 ** | 0.76 | 0.60 ** | 1.12 ** | 0.92 ** | 0.08 ** |
| GCA-Female (Set)*E | 168 | 1,191,615.00 ** | 6.54 ** | 8.02 ** | 0.12 | 342.87 ** | 100.16 ** | 0.74 ** | 0.79 | 0.77 ** | 1.44 ** | 1.01 ** | 0.09 ** |
| SCA (Set)*E | 672 | 1,018,925.00 ** | 5.21 ** | 6.84 ** | 0.14 | 318.81 ** | 85.37 ** | 0.72 ** | 0.84 ** | 0.44 ** | 1.05 ** | 0.8 ** | 0.08 ** |
| Error | 960 | 652,605.00 | 3.81 | 5.03 | 0.13 | 190.52 | 61.93 | 0.52 | 0.66 | 0.32 | 0.57 | 0.60 | 0.05 |
| Trait | Optimal | Striga-Infested | Across Environments | ||||||
|---|---|---|---|---|---|---|---|---|---|
| GCA | SCA | GCA | SCA | GCA | SCA | ||||
| Male | Female | Male | Female | Male | Female | ||||
| Grain yield | 19.10 | 25.95 | 54.95 | 16.57 | 22.40 | 48.37 | 20.97 | 29.80 | 49.23 |
| Days to anthesis | 26.98 | 28.61 | 44.41 | 25.48 | 19.96 | 46.79 | 31.15 | 26.71 | 42.14 |
| Days to silking | 26.91 | 29.27 | 43.81 | 26.56 | 15.82 | 50.35 | 33.87 | 26.32 | 39.81 |
| Anthesis–silking interval | 14.58 | 18.29 | 67.13 | 9.66 | 18.48 | 64.61 | 10.06 | 20.03 | 69.91 |
| Plant height | 29.42 | 31.74 | 38.84 | 16.39 | 16.95 | 35.33 | 30.97 | 32.33 | 36.70 |
| Ear height | 21.18 | 37.82 | 41.00 | 8.37 | 19.95 | 51.46 | 19.50 | 39.83 | 40.67 |
| Root lodging | 24.45 | 14.14 | 61.41 | 14.90 | 21.74 | 57.37 | 25.32 | 18.07 | 56.61 |
| Stalk lodging | 23.36 | 20.29 | 56.34 | 16.24 | 16.08 | 58.74 | 20.21 | 22.01 | 57.78 |
| Husk cover | 22.14 | 22.02 | 55.84 | 18.15 | 21.00 | 55.15 | 17.45 | 24.77 | 57.79 |
| Plant aspect | 13.30 | 24.75 | 61.94 | 16.40 | 13.04 | 47.83 | 20.70 | 19.74 | 59.56 |
| Ear aspect | 21.39 | 27.09 | 51.53 | 16.31 | 15.55 | 48.46 | 19.08 | 22.26 | 58.67 |
| Ears per plant | 23.37 | 24.84 | 51.79 | 19.31 | 14.16 | 49.28 | 23.83 | 21.90 | 54.28 |
| Striga damage syndrome rating at 8 WAP | 20.69 | 22.86 | 40.16 | ||||||
| Striga damage syndrome rating at 10 WAP | 13.37 | 25.66 | 45.40 | ||||||
| Number of emerged Striga plants at 8 WAP | 18.82 | 12.43 | 51.90 | ||||||
| Number of emerged Striga plants at 10 WAP | 19.70 | 12.13 | 51.98 | ||||||
| Method | Group 1 | Group 2 | Group 3 | Group 4 | Group 5 |
|---|---|---|---|---|---|
| HGCAMT-Striga-infested environment | TZEI 3A, TZdEI 283, TZEI 456, TZdEI 69, TZEI 378, TZEI 467, TZEI 298, TZEI 497, TZEI 365, TZEI 376, TZEI 520 | TZdEI 192, TZEI 468, TZdEI 272, TZEI 470, TZEI 472 | TZEI 379, TZEI 449, TZEI 462, TZEI 485 | TZEI 8, TZdEI 40, TZdEI 216, TZEI 127, TZdEI 215, TZdEI 238, TZEI 402, TZEI 475, TZdEI 124, TZEI 461 | |
| HGCAMT-Optimal environment | TZEI 3A, TZEI 497, TZdEI 283, TZEI 462, TZdEI 216, TZdEI 238, TZEI 485, TZEI 402, TZEI 472, TZEI 402, TZEI 456, TZEI 298, TZEI 461 | TZdEI 192, TZdEI 272, TZEI 475, TZEI 449, TZEI 365, TZEI 379, TZEI 520, TZEI 378, TZEI 467, TZEI 468, TZEI 470 | TZEI 8, TZdEI 124, TZdEI 69, TZdEI 40, TZEI 127, TZdEI 215 | ||
| HGCAMT-Across environment | TZEI 3A, TZEI 462, TZdEI 192, TZEI 475, TZdEI 238, TZEI 402, TZdEI 283, TZEI 472, TZEI 379, TZEI 456, TZEI 468 | TZdEI 69, TZdEI 272, TZEI 485, TZEI 497, TZEI 376, TZEI 470, TZEI 467, TZEI 461, TZEI 298, TZEI 378, TZEI 365, TZEI 520, TZEI 449 | TZEI 8, TZdEI 124, TZdEI 40, TZEI 127, TZdEI 215, TZdEI 216 | ||
| SNP-GD | TZEI 8, TZEI 127, TZdEI 40, TZdEI 192, TZdEI 283, TZdEI 216, TZdEI 69, TZdEI 124, TZdEI 272 | TZdEI 215, TZdEI 238 | TZEI 365, TZEI 376, TZEI 298, TZEI 378, TZEI 379, TZEI 3A, TZEI 402 | TZEI 468, TZEI 467, TZEI 472, TZEI 470 | TZEI 461, TZEI 462, TZEI 497, TZEI 475, TZEI 449, TZEI 485, TZEI 520, TZEI 456 |
| Optimal Growing Environments | |||
| Yield Group | Cross Type | HGCAMT | SNP-GD |
| 1 | inter-group | 36 | 36 |
| 1 | intra-group | 14 | 14 |
| 2 | inter-group | 31 | 30 |
| 2 | intra-group | 19 | 20 |
| 3 | inter-group | 20 | 28 |
| 3 | intra-group | 30 | 22 |
| BE | 44.50 | 38.79 | |
| Striga-Infested Environments | |||
| Yield Group | Cross Type | HGCAMT | SNP-GD |
| 1 | inter-group | 36 | 41 |
| 1 | intra-group | 14 | 9 |
| 2 | inter-group | 38 | 28 |
| 2 | intra-group | 12 | 22 |
| 3 | inter-group | 32 | 25 |
| 3 | intra-group | 18 | 25 |
| BE | 37.44 | 44.13 | |
| Across Environments | |||
| Yield Group | Cross Type | HGCAMT | SNP-GD |
| 1 | inter-group | 37 | 43 |
| 1 | intra-group | 13 | 7 |
| 2 | inter-group | 31 | 33 |
| 2 | intra-group | 19 | 17 |
| 3 | inter-group | 21 | 19 |
| 3 | intra-group | 29 | 31 |
| BE | 44.56 | 50.81 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Adu, G.B.; Badu-Apraku, B.; Akromah, R.; Awuku, F.J. Combining Abilities and Heterotic Patterns among Early Maturing Maize Inbred Lines under Optimal and Striga-Infested Environments. Genes 2022, 13, 2289. https://doi.org/10.3390/genes13122289
Adu GB, Badu-Apraku B, Akromah R, Awuku FJ. Combining Abilities and Heterotic Patterns among Early Maturing Maize Inbred Lines under Optimal and Striga-Infested Environments. Genes. 2022; 13(12):2289. https://doi.org/10.3390/genes13122289
Chicago/Turabian StyleAdu, Gloria Boakyewaa, Baffour Badu-Apraku, Richard Akromah, and Frederick Justice Awuku. 2022. "Combining Abilities and Heterotic Patterns among Early Maturing Maize Inbred Lines under Optimal and Striga-Infested Environments" Genes 13, no. 12: 2289. https://doi.org/10.3390/genes13122289
APA StyleAdu, G. B., Badu-Apraku, B., Akromah, R., & Awuku, F. J. (2022). Combining Abilities and Heterotic Patterns among Early Maturing Maize Inbred Lines under Optimal and Striga-Infested Environments. Genes, 13(12), 2289. https://doi.org/10.3390/genes13122289