

Genome-Wide Identification and In Silico Analysis of ZF-HD Transcription Factor Genes in Zea mays L.

 , ,
, ,  ,
,  , , , ,
, , , , 

Abstract
1. Introduction
2. Results
2.1. Identification of ZHD Family Genes and Sequence Analysis of Their Proteins
2.2. Sequence Alignment and Phylogenetic Tree Construction
2.3. Chromosomal Location, Gene Structure, and Motif Composition Analysis
2.4. Analysis of Cis-Elements in ZmZHD Promoters
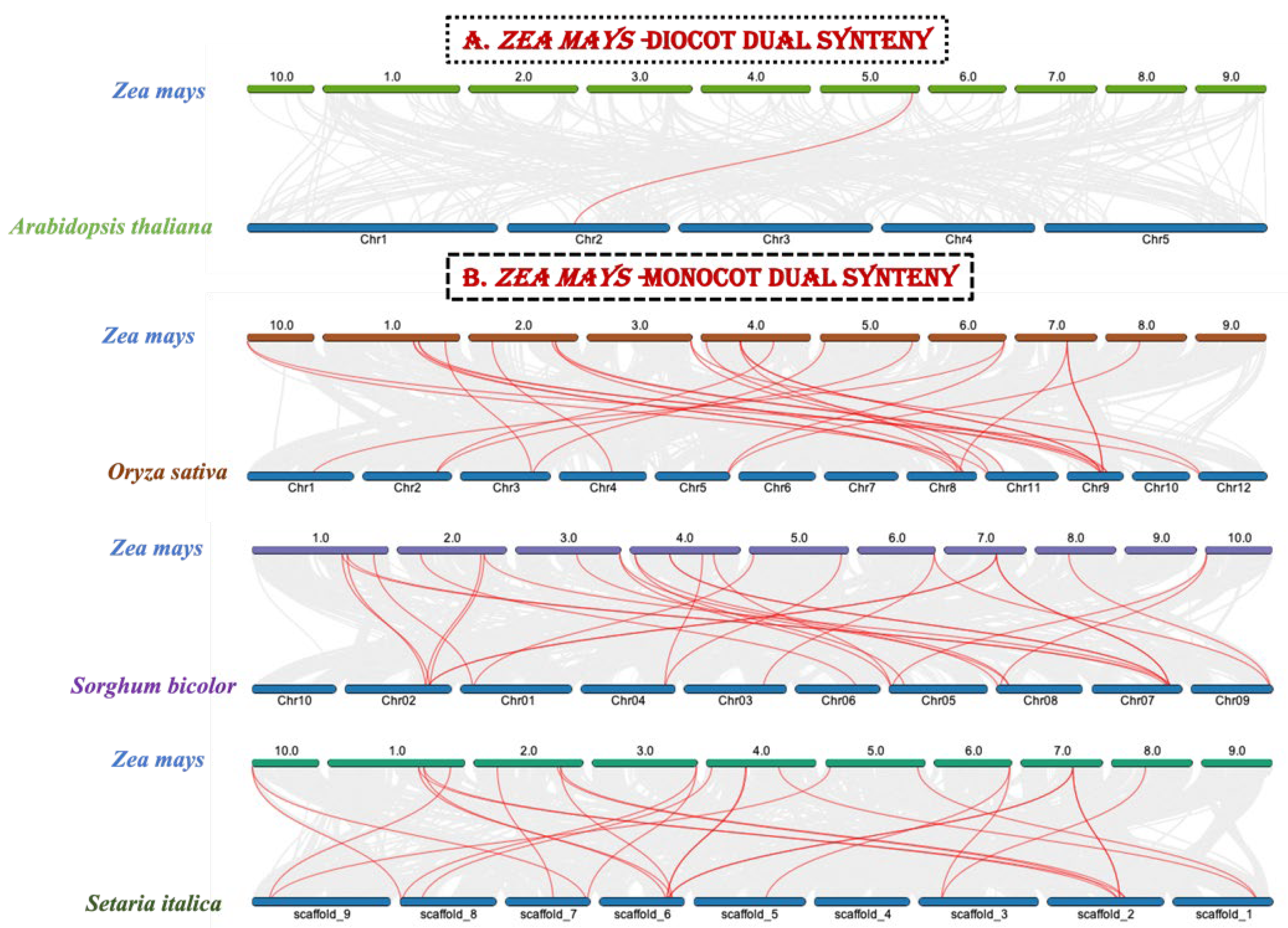
2.5. Synteny and Evolutionary Analysis of ZmZHDGenes
2.6. Construction of a PPI Network and Expression Profiling of ZmZHD Genes in Various Tissues
2.7. MiRNA Target Site Prediction and Validation
3. Discussion
4. Materials and Methods
4.1. Gene Retrieval and Sequence Analysis of the ZHD Gene Family in Maize
4.2. Sequence Alignment and Phylogenetic Tree Construction
4.3. Chromosomal Locations, Motif Compositions, and Exon and Intron Distributions
4.4. Cis-Acting Elements and Functional Prediction
4.5. Synteny Analysis of ZHD Proteins and ks/ka Ratios
4.6. Protein–Protein Interaction and Z. mays L. RNA-Sequencing Data Analysis
4.7. MiRNA Target Site Prediction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, W.; Wu, P.; Li, Y.; Hou, X. Genome-wide analysis and expression patterns of ZF-HD transcription factors under different developmental tissues and abiotic stresses in Chinese cabbage. Mol. Genet. Genom. 2016, 291, 1451–1464. [Google Scholar] [CrossRef] [PubMed]
- Khoso, M.A.; Hussain, A.; Ritonga, F.N.; Ali, Q. WRKY transcription factors (TFs): Molecular switches to regulate drought, temperature, and salinity stresses in plants. Front. Plant Sci. 2022, 13, 1039329. [Google Scholar] [CrossRef]
- Ali, Q.; Ahsan, M.; Ali, F.; Aslam, M.; Khan, N.H.; Munzoor, M.; Mustafa, H.S.B.; Muhammad, S. Heritability, heterosis and heterobeltiosis studies for morphological traits of maize (Zea mays L.) seedlings. Adv. Life Sci. 2013, 1, 52–63. [Google Scholar]
- Ali, Q.; Ahsan, M.; Kanwal, N.; Ali, F.; Ali, A.; Ahmed, W.; Ishfaq, M.; Saleem, M. Screening for drought tolerance: Comparison of maize hybrids under water deficit condition. Adv. Life Sci. 2016, 3, 51–58. [Google Scholar]
- Ali, Q.; Ali, A.; Ahsan, M.; Nasir, I.A.; Abbas, H.G.; Ashraf, M.A. Line× Tester analysis for morpho-physiological traits of Zea mays L seedlings. Adv. Life Sci. 2014, 1, 242–253. [Google Scholar]
- Jalal, A.; Ali, Q.; Manghwar, H.; Zhu, D. Identification, Phylogeny, Divergence, Structure, and Expression Analysis of A20/AN1 Zinc Finger Domain Containing Stress-Associated Proteins (SAPs) Genes in Jatropha curcas L. Genes 2022, 13, 1766. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Mu, G.; Hussain, A.; Yuanyuan, Q.; Yu, C.; Xu, Y.; Manghwar, H.; Gu, Q.; Wu, H.; et al. Revealing Plant Growth-Promoting Mechanisms of Bacillus strains in Elevating Rice Growth and Its Interaction with Salt Stress. Front. Plant Sci. 2022, 13, 17. [Google Scholar] [CrossRef]
- Ali, Q.; Ayaz, M.; Yu, C.; Wang, Y.; Gu, Q.; Wu, H.; Gao, X. Cadmium Tolerant Microbial Strains Possess Different Mechanisms for Cadmium Biosorption and Immobilization in Rice Seedlings. Chemosphere 2022, 303, 135206. [Google Scholar] [CrossRef]
- Rajer, F.U.; Samma, M.K.; Ali, Q.; Rajar, W.A.; Gao, X. Bacillus spp.-Mediated Growth Promotion of Rice Seedlings and Suppression of Bacterial Blight Disease under Greenhouse Conditions. Pathogens 2022, 11, 1251. [Google Scholar] [CrossRef]
- Ali, Q.; Yu, C.; Hussain, A.; Ali, M.; Ahmar, S.; Sohail, M.A. Genome engineering technology for durable disease resistance: Recent progress and future outlooks for sustainable agriculture. Front. Plant Sci. 2022, 13, 860281. [Google Scholar] [CrossRef]
- Mitsuda, N.; Ohme-Takagi, M. Functional analysis of transcription factors in Arabidopsis. Plant Cell Physiol. 2009, 50, 1232–1248. [Google Scholar] [CrossRef]
- Amorim, L.L.; da Fonseca Dos Santos, R.; Neto, J.P.B.; Guida-Santos, M.; Crovella, S.; Benko-Iseppon, A.M. Transcription factors involved in plant resistance to pathogens. Curr. Protein Pept. Sci. 2017, 18, 335–351. [Google Scholar] [CrossRef]
- Ariel, F.D.; Manavella, P.A.; Dezar, C.A.; Chan, R.L. The true story of the HD-Zip family. Trends Plant Sci. 2007, 12, 419–426. [Google Scholar] [CrossRef]
- Englbrecht, C.C.; Schoof, H.; Böhm, S. Conservation, diversification and expansion of C2H2 zinc finger proteins in the Arabidopsis thaliana genome. BMC Genom. 2004, 5, 39. [Google Scholar] [CrossRef]
- Halbach, T.; Scheer, N.; Werr, W. Transcriptional activation by the PHD finger is inhibited through an adjacent leucine zipper that binds 14–3-3 proteins. Nucleic Acids Res. 2000, 28, 3542–3550. [Google Scholar] [CrossRef]
- Windhövel, A.; Hein, I.; Dabrowa, R.; Stockhaus, J. Characterization of a novel class of plant homeodomain proteins that bind to the C4 phosphoenolpyruvate carboxylase gene of Flaveria trinervia. Plant Mol. Biol. 2001, 45, 201–214. [Google Scholar] [CrossRef]
- Krishna, S.S.; Majumdar, I.; Grishin, N.V. Structural classification of zinc fingers: Survey and summary. Nucleic Acids Res. 2003, 31, 532–550. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Jiang, Q.; Wang, R. Salt Tolerant Bacillus Strains Improve Plant Growth Traits and Regulation of Phytohormones in Wheat under Salinity Stress. Plants 2022, 11, 2769. [Google Scholar] [CrossRef]
- Sun, J.; Xie, M.; Li, X.; Li, Z.; Wang, Q.; Ding, A.; Wang, W.; Sun, Y. Systematic investigations of the ZF-HD gene family in tobacco reveal their multiple roles in abiotic stresses. Agronomy 2021, 11, 406. [Google Scholar] [CrossRef]
- Yong, Y.; Zhang, Y.; Lyu, Y. Functional characterization of Lilium lancifolium cold-responsive Zinc Finger Homeodomain (ZFHD) gene in abscisic acid and osmotic stress tolerance. PeerJ 2021, 9, e11508. [Google Scholar] [CrossRef]
- Shalmani, A.; Muhammad, I.; Sharif, R.; Zhao, C.; Ullah, U.; Zhang, D.; Jing, X.-Q.; Amin, B.; Jia, P.; Mobeen Tahir, M. Zinc finger-homeodomain genes: Evolution, functional differentiation, and expression profiling under flowering-related treatments and abiotic stresses in plants. Evol. Bioinform. 2019, 15, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Barth, O.; Vogt, S.; Uhlemann, R.; Zschiesche, W.; Humbeck, K. Stress induced and nuclear localized HIPP26 from Arabidopsis thaliana interacts via its heavy metal associated domain with the drought stress related zinc finger transcription factor ATHB29. Plant Mol. Biol. 2009, 69, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.P.; Nakashima, K.; Sakuma, Y.; Osakabe, Y.; Qin, F.; Simpson, S.D.; Maruyama, K.; Fujita, Y.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Co-expression of the stress-inducible zinc finger homeodomain ZFHD1 and NAC transcription factors enhances expression of the ERD1 gene in Arabidopsis. Plant J. 2007, 49, 46–63. [Google Scholar] [CrossRef] [PubMed]
- Perrella, G.; Davidson, M.L.; O’Donnell, L.; Nastase, A.-M.; Herzyk, P.; Breton, G.; Pruneda-Paz, J.L.; Kay, S.A.; Chory, J.; Kaiserli, E. ZINC-FINGER interactions mediate transcriptional regulation of hypocotyl growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E4503–E4511. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Barros, P.M.; Cordeiro, A.M.; Serra, T.S.; Lourenço, T.; Chander, S.; Oliveira, M.M.; Saibo, N.J. Seven zinc-finger transcription factors are novel regulators of the stress responsive gene OsDREB1B. J. Exp. Bot. 2012, 63, 3643–3656. [Google Scholar] [CrossRef]
- Lai, W.; Zhu, C.; Hu, Z.; Liu, S.; Wu, H.; Zhou, Y. Identification and transcriptional analysis of zinc finger-homeodomain (ZF-HD) family genes in cucumber. Biochem. Genet. 2021, 59, 884–901. [Google Scholar] [CrossRef]
- Liu, H.; Yang, Y.; Zhang, L. Zinc finger-homeodomain transcriptional factors (ZF-HDs) in wheat (Triticum aestivum L.): Identification, evolution, expression analysis and response to abiotic stresses. Plants 2021, 10, 593. [Google Scholar] [CrossRef]
- Khatun, K.; Nath, U.K.; Robin, A.H.K.; Park, J.-I.; Lee, D.-J.; Kim, M.-B.; Kim, C.K.; Lim, K.-B.; Nou, I.S.; Chung, M.-Y. Genome-wide analysis and expression profiling of zinc finger homeodomain (ZHD) family genes reveal likely roles in organ development and stress responses in tomato. BMC Genom. 2017, 18, 695. [Google Scholar] [CrossRef]
- Liu, M.; Sun, W.; Ma, Z.; Zheng, T.; Huang, L.; Wu, Q.; Zhao, G.; Tang, Z.; Bu, T.; Li, C. Genome-wide investigation of the AP2/ERF gene family in tartary buckwheat (Fagopyum Tataricum). BMC Plant Biol. 2019, 19, 342. [Google Scholar] [CrossRef]
- Wang, C.; Shi, X.; Liu, L.; Li, H.; Ammiraju, J.S.; Kudrna, D.A.; Xiong, W.; Wang, H.; Dai, Z.; Zheng, Y. Genomic resources for gene discovery, functional genome annotation, and evolutionary studies of maize and its close relatives. Genetics 2013, 195, 723–737. [Google Scholar] [CrossRef]
- Ahuja, I.; de Vos, R.C.; Bones, A.M.; Hall, R.D. Plant molecular stress responses face climate change. Trends Plant Sci. 2010, 15, 664–674. [Google Scholar] [CrossRef]
- Gujjar, R.S.; Akhtar, M.; Singh, M. Transcription factors in abiotic stress tolerance. Indian J. Plant Physiol. 2014, 19, 306–316. [Google Scholar] [CrossRef]
- Udvardi, M.K.; Kakar, K.; Wandrey, M.; Montanari, O.; Murray, J.; Andriankaja, A.; Zhang, J.-Y.; Benedito, V.; Hofer, J.M.; Chueng, F. Legume transcription factors: Global regulators of plant development and response to the environment. Plant Physiol. 2007, 144, 538–549. [Google Scholar] [CrossRef]
- Zhang, P.; Wei, L.; Cao, L.; Qiu, X.; Fu, J.; Wang, G.; Ku, L.; Wang, T. Function analysis of ZmZHD9, a positive regulator in drought stress response in transgenic maize. Res. Sq. 2020. [Google Scholar] [CrossRef]
- Schnable, P.S.; Ware, D.; Fulton, R.S.; Stein, J.C.; Wei, F.; Pasternak, S.; Liang, C.; Zhang, J.; Fulton, L.; Graves, T.A. The B73 maize genome: Complexity, diversity, and dynamics. Science 2009, 326, 1112–1115. [Google Scholar] [CrossRef]
- Yilmaz, A.; Nishiyama, M.Y.; Fuentes, B.G.; Souza, G.M.; Janies, D.; Gray, J.; Grotewold, E. GRASSIUS: A platform for comparative regulatory genomics across the grasses. Plant Physiol. 2009, 149, 171–180. [Google Scholar] [CrossRef]
- Nai, G.; Mao, J.; Lu, S.; Li, Y.; Ma, Z.; Chen, B. Genome-wide analysis and expression characterization of zinc finger homeodomain (ZHD) family genes responsed to different abiotic stresses and hormonal treatments in grape (Vitis vinifera L.). Res. Sq. 2020. [Google Scholar] [CrossRef]
- Ren, R.; Wang, H.; Guo, C.; Zhang, N.; Zeng, L.; Chen, Y.; Ma, H.; Qi, J. Widespread whole genome duplications contribute to genome complexity and species diversity in angiosperms. Mol. Plant 2018, 11, 414–428. [Google Scholar] [CrossRef]
- Khalid, M.H.B.; Raza, M.A.; Yu, H.Q.; Khan, I.; Sun, F.A.; Feng, L.Y.; Qu, J.T.; Fu, F.L.; Li, W.C. Expression, subcellular localization, and interactions of CPK family genes in maize. Int. J. Mol. Sci. 2019, 20, 6173. [Google Scholar] [CrossRef]
- Zhou, C.; Zhu, C.; Xie, S.; Weng, J.; Lin, Y.; Lai, Z.; Guo, Y. Genome-wide analysis of zinc finger motif-associated homeodomain (ZF-HD) family genes and their expression profiles under abiotic stresses and phytohormones stimuli in tea plants (Camellia sinensis). Sci. Hortic. 2021, 281, 109976. [Google Scholar] [CrossRef]
- Li, Y.; Bai, B.; Wen, F.; Zhao, M.; Xia, Q.; Yang, D.-H.; Wang, G. Genome-wide identification and expression analysis of HD-ZIP I gene subfamily in Nicotiana tabacum. Genes 2019, 10, 575. [Google Scholar] [CrossRef] [PubMed]
- Hu, J.; Gao, Y.; Zhao, T.; Li, J.; Yao, M.; Xu, X. Genome-wide identification and expression pattern analysis of zinc-finger homeodomain transcription factors in tomato under abiotic stress. J. Am. Soc. Hortic. Sci. 2018, 143, 14–22. [Google Scholar] [CrossRef]
- Hu, B.; Jin, J.; Guo, A.-Y.; Zhang, H.; Luo, J.; Gao, G. GSDS 2.0: An upgraded gene feature visualization server. Bioinformatics 2015, 31, 1296–1297. [Google Scholar] [CrossRef] [PubMed]
- Niu, H.; Xia, P.; Hu, Y.; Zhan, C.; Li, Y.; Gong, S.; Li, Y.; Ma, D. Genome-wide identification of ZF-HD gene family in Triticum aestivum: Molecular evolution mechanism and function analysis. PLoS ONE 2021, 16, e0256579. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yin, X.; Li, X.; Wang, L.; Zheng, Y.; Xu, X.; Zhang, Y.; Wang, X. Genome-wide identification, evolution and expression analysis of the grape (Vitis vinifera L.) zinc finger-homeodomain gene family. Int. J. Mol. Sci. 2014, 15, 5730–5748. [Google Scholar] [CrossRef]
- Wang, D.-P.; Wan, H.-L.; Zhang, S.; Yu, J. γ-MYN: A new algorithm for estimating Ka and Ks with consideration of variable substitution rates. Biol. Direct 2009, 4, 20. [Google Scholar] [CrossRef]
- Huang, H.; Ayaz, A.; Zheng, M.; Yang, X.; Zaman, W.; Zhao, H.; Lü, S. Arabidopsis KCS5 and KCS6 Play Redundant Roles in Wax Synthesis. Int. J. Mol. Sci. 2022, 23, 4450. [Google Scholar] [CrossRef]
- Abdullah, M.; Cheng, X.; Cao, Y.; Su, X.; Manzoor, M.A.; Gao, J.; Cai, Y.; Lin, Y. Zinc finger-homeodomain transcriptional factors (ZHDs) in upland cotton (Gossypium hirsutum): Genome-wide identification and expression analysis in fiber development. Front. Genet. 2018, 9, 357. [Google Scholar] [CrossRef]
- Ayaz, A.; Huang, H.; Zheng, M.; Zaman, W.; Li, D.; Saqib, S.; Zhao, H.; Lü, S. Molecular cloning and functional analysis of GmLACS2–3 reveals its involvement in cutin and suberin biosynthesis along with abiotic stress tolerance. Int. J. Mol. Sci. 2021, 22, 9175. [Google Scholar] [CrossRef]
- Li, D.; Pan, C.; Lu, J.; Zaman, W.; Zhao, H.; Zhang, J.; Lü, S. Lupeol Accumulation Correlates with Auxin in the Epidermis of Castor. Molecules 2021, 26, 2978. [Google Scholar] [CrossRef]
- Khan, M.; Ali, S.; Manghwar, H.; Saqib, S.; Ullah, F.; Ayaz, A.; Zaman, W. Melatonin function and crosstalk with other phytohormones under normal and stressful conditions. Genes 2022, 13, 1699. [Google Scholar] [CrossRef]
- Liaquat, F.; Munis, M.F.H.; Arif, S.; Haroon, U.; Shi, J.; Saqib, S.; Zaman, W.; Che, S.; Liu, Q. PacBio single-molecule long-read sequencing reveals genes tolerating manganese stress in Schima superba saplings. Front. Genet. 2021, 12, 635043. [Google Scholar] [CrossRef]
- Kong, H.; Landherr, L.L.; Frohlich, M.W.; Leebens-Mack, J.; Ma, H.; DePamphilis, C.W. Patterns of gene duplication in the plant SKP1 gene family in angiosperms: Evidence for multiple mechanisms of rapid gene birth. Plant J. 2007, 50, 873–885. [Google Scholar] [CrossRef]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Tan, Q.K.-G.; Irish, V.F. The Arabidopsis zinc finger-homeodomain genes encode proteins with unique biochemical properties that are coordinately expressed during floral development. Plant Physiol. 2006, 140, 1095–1108. [Google Scholar] [CrossRef]
- Simon, M.; Bruex, A.; Kainkaryam, R.M.; Zheng, X.; Huang, L.; Woolf, P.J.; Schiefelbein, J. Tissue-specific profiling reveals transcriptome alterations in Arabidopsis mutants lacking morphological phenotypes. Plant Cell 2013, 25, 3175–3185. [Google Scholar] [CrossRef]
- Hu, W.; DePamphilis, C.W.; Ma, H. Phylogenetic analysis of the plant-specific zinc finger-homeobox and mini zinc finger gene families. J. Integr. Plant Biol. 2008, 50, 1031–1045. [Google Scholar] [CrossRef]
- Zhao, B.; Liang, R.; Ge, L.; Li, W.; Xiao, H.; Lin, H.; Ruan, K.; Jin, Y. Identification of drought-induced microRNAs in rice. Biochem. Biophys. Res. Commun. 2007, 354, 585–590. [Google Scholar] [CrossRef]
- Zhou, X.; Wang, G.; Sutoh, K.; Zhu, J.-K.; Zhang, W. Identification of cold-inducible microRNAs in plants by transcriptome analysis. Biochim. Biophys. Acta BBA Gene Regul. Mech. 2008, 1779, 780–788. [Google Scholar] [CrossRef]
- Sunkar, R.; Zhou, X.; Zheng, Y.; Zhang, W.; Zhu, J.-K. Identification of novel and candidate miRNAs in rice by high throughput sequencing. BMC Plant Biol. 2008, 8, 25. [Google Scholar] [CrossRef]
- Cheng, Y.; Long, M. A cytosolic NADP-malic enzyme gene from rice (Oryza sativa L.) confers salt tolerance in transgenic Arabidopsis. Biotechnol. Lett. 2007, 29, 1129–1134. [Google Scholar] [PubMed]
- Ding, D.; Zhang, L.; Wang, H.; Liu, Z.; Zhang, Z.; Zheng, Y. Differential expression of miRNAs in response to salt stress in maize roots. Ann. Bot. 2009, 103, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Finn, R.D.; Clements, J.; Eddy, S.R. HMMER web server: Interactive sequence similarity searching. Nucleic Acids Res. 2011, 39, W29–W37. [Google Scholar] [CrossRef]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 2007, 23, 127–128. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v5: An online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 2021, 49, W293–W296. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Y.; Fei, Z. iTAK-Identification and Classification of Plant Transcription Factors and Protein Kinases. In Proceedings of the Plant and Animal Genome XXII Conference, San Diego, CA, USA, 11–15 January 2014. [Google Scholar]
- Jin, J.; Tian, F.; Yang, D.-C.; Meng, Y.-Q.; Kong, L.; Luo, J.; Gao, G. PlantTFDB 4.0: Toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2017, 45, D1040–D1045. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Nicholas, K.B. GeneDoc: Analysis and visualization of genetic variation. Embnew. News 1997, 4, 14. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular evolutionary genetics analysis version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Jiangtao, C.; Yingzhen, K.; Qian, W.; Yuhe, S.; Daping, G.; Jing, L.; Guanshan, L. MapGene2Chrom, a tool to draw gene physical map based on Perl and SVG languages. Yi Chuan Hered. 2015, 37, 91–97. [Google Scholar]
- Bailey, T.L.; Boden, M.; Buske, F.A.; Frith, M.; Grant, C.E.; Clementi, L.; Ren, J.; Li, W.W.; Noble, W.S. MEME SUITE: Tools for motif discovery and searching. Nucleic Acids Res. 2009, 37, W202–W208. [Google Scholar] [CrossRef]
- Lescot, M.; Déhais, P.; Thijs, G.; Marchal, K.; Moreau, Y.; Van de Peer, Y.; Rouzé, P.; Rombauts, S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002, 30, 325–327. [Google Scholar] [CrossRef]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Wang, Y.; Tang, H.; DeBarry, J.D.; Tan, X.; Li, J.; Wang, X.; Lee, T.-H.; Jin, H.; Marler, B.; Guo, H. MCScanX: A toolkit for detection and evolutionary analysis of gene synteny and collinearity. Nucleic Acids Res. 2012, 40, e49. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef]
- Song, M.; Zhang, Y.; Wang, L.; Peng, X. Genome-wide identification and phylogenetic analysis of zinc finger Homeodomain family genes in Brassica napus. Chin. Bull. Bot. 2019, 54, 699. [Google Scholar]
- Sievers, F.; Higgins, D.G. Clustal Omega, accurate alignment of very large numbers of sequences. In Multiple Sequence Alignment Methods; Springer: Berlin, Germany, 2014; pp. 105–116. [Google Scholar]
- Suyama, M.; Torrents, D.; Bork, P. PAL2NAL: Robust conversion of protein sequence alignments into the corresponding codon alignments. Nucleic Acids Res. 2006, 34, W609–W612. [Google Scholar] [CrossRef]
- Petryszak, R.; Keays, M.; Tang, Y.A.; Fonseca, N.A.; Barrera, E.; Burdett, T.; Füllgrabe, A.; Fuentes, A.M.-P.; Jupp, S.; Koskinen, S. Expression Atlas update–An integrated database of gene and protein expression in humans, animals and plants. Nucleic Acids Res. 2016, 44, D746–D752. [Google Scholar] [CrossRef]
- Walley, J.W.; Sartor, R.C.; Shen, Z.; Schmitz, R.J.; Wu, K.J.; Urich, M.A.; Nery, J.R.; Smith, L.G.; Schnable, J.C.; Ecker, J.R. Integration of omic networks in a developmental atlas of maize. Science 2016, 353, 814–818. [Google Scholar] [CrossRef]
- Kolde, R.; Vilo, J. GOsummaries: An R package for visual functional annotation of experimental data. F1000Research 2015, 4, 574. [Google Scholar] [CrossRef]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, W155–W159. [Google Scholar] [CrossRef] [PubMed]
- Otasek, D.; Morris, J.H.; Bouças, J.; Pico, A.R.; Demchak, B. Cytoscape automation: Empowering workflow-based network analysis. Genome Biol. 2019, 20, 185. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene ID | Chromosome Location | Protein Length (aa) | Mol. Wt. 1KDA | pI 1 | GRAVY 1 | Exon | Intron | Subcellular Location | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Number | Start | End | Length | |||||||||
| ZmZHD1 | GRMZM2G068330 | 4 | 11278503 | 112881332 | 250330460 | 382 | 39,774.87 | 8.13 | −0.58 | 2 | 1 | Nucleus |
| ZmZHD2 | GRMZM2G161315 | 7 | 112902252 | 1129906963 | 185808916 | 370 | 38,611.45 | 8.21 | −0.524 | 1 | Cell wall, nucleus | |
| ZmZHD3 | GRMZM2G346920 | 1 | 200895999 | 200900836 | 308452471 | 361 | 37,825.61 | 7.03 | −0.515 | 1 | Chloroplast, nucleus | |
| ZmZHD4 | GRMZM2G425236 | 5 | 10221783 | 10225985 | 226353449 | 240 | 24,975.74 | 7.16 | −0.629 | 1 | Nucleus | |
| ZmZHD5 | GRMZM2G438438 | 1 | 212576317 | 212581228 | 308452471 | 373 | 38,662.73 | 7.21 | −0.31 | 1 | Nucleus | |
| ZmZHD6 | GRMZM2G414844 | 6 | 166374440 | 166378910 | 181357234 | 242 | 26,692.75 | 8.8 | −0.985 | 1 | Nucleus | |
| ZmZHD7 | GRMZM2G353734 | 4 | 85319366 | 85325092 | 250330460 | 526 | 55,497.21 | 9.02 | −0.575 | 3 | 2 | Nucleus |
| ZmZHD8 | GRMZM2G423423 | 1 | 269145949 | 269149835 | 308452471 | 231 | 24,075.72 | 8.35 | −0.687 | 1 | Nucleus | |
| ZmZHD9 | GRMZM2G353076 | 3 | 230480803 | 230481862 | 238017767 | 100 | 10,401.41 | 8.93 | −0.608 | 1 | Nucleus | |
| ZmZHD10 | GRMZM2G470974 | 10 | 2632775 | 2633842 | 152435371 | 98 | 10,100.06 | 6.87 | −0.628 | 1 | Nucleus | |
| ZmZHD11 | GRMZM2G328438 | 8 | 73654879 | 73656447 | 182411202 | 254 | 27,689.12 | 8.51 | −0.805 | 1 | Nucleus | |
| ZmZHD12 | GRMZM2G417229 | 5 | 201282254 | 201284278 | 226353449 | 302 | 32,353.12 | 6.95 | −0.746 | 1 | Nucleus | |
| ZmZHD13 | GRMZM2G071112 | 7 | 112658777 | 112661470 | 185808916 | 89 | 9802.04 | 8.16 | −0.361 | 5 | 4 | Nucleus |
| ZmZHD14 | GRMZM2G172586 | 10 | 2639262 | 2640147 | 152435371 | 98 | 10,113.1 | 7.59 | −0.628 | 1 | Nucleus | |
| ZmZHD15 | GRMZM2G089619 | 2 | 50140925 | 50142374 | 243,675,191 | 300 | 31,130.79 | 6.72 | −0.437 | 1 | Nucleus | |
| ZmZHD16 | GRMZM2G389379 | 2 | 188271896 | 188273136 | 243,675,191 | 286 | 29,912.64 | 6.96 | −0.485 | 1 | Nucleus | |
| ZmZHD17 | GRMZM2G069365 | 4 | 160153804 | 160155930 | 250330460 | 446 | 48,227.78 | 6.64 | −0.793 | 1 | Nucleus | |
| ZmZHD18 | GRMZM2G462417 | 4 | 185816491 | 185819532 | 250330460 | 655 | 71,591.15 | 9.01 | −0.046 | 3 | 2 | Nucleus |
| ZmZHD19 | GRMZM2G370863 | 4 | 85758755 | 85759432 | 250330460 | 127 | 13,351.19 | 7.53 | −0.138 | 1 | Nucleus | |
| ZmZHD20 | GRMZM2G051955 | 2 | 181822882 | 181824556 | 243675191 | 361 | 37,714.29 | 7.73 | −0.578 | 1 | Cell wall, nucleus | |
| ZmZHD21 | GRMZM5G821755 | 3 | 136934215 | 136936196 | 238017767 | 331 | 35,115.48 | 8.57 | −0.593 | 2 | 1 | Nucleus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Islam, M.A.U.; Nupur, J.A.; Khalid, M.H.B.; Din, A.M.U.; Shafiq, M.; Alshegaihi, R.M.; Ali, Q.; Ali, Q.; Kamran, Z.; Manzoor, M.; et al. Genome-Wide Identification and In Silico Analysis of ZF-HD Transcription Factor Genes in Zea mays L. Genes 2022, 13, 2112. https://doi.org/10.3390/genes13112112
Islam MAU, Nupur JA, Khalid MHB, Din AMU, Shafiq M, Alshegaihi RM, Ali Q, Ali Q, Kamran Z, Manzoor M, et al. Genome-Wide Identification and In Silico Analysis of ZF-HD Transcription Factor Genes in Zea mays L. Genes. 2022; 13(11):2112. https://doi.org/10.3390/genes13112112
Chicago/Turabian StyleIslam, Md. Abir Ul, Juthy Abedin Nupur, Muhammad Hayder Bin Khalid, Atta Mohi Ud Din, Muhammad Shafiq, Rana M. Alshegaihi, Qurban Ali, Qurban Ali, Zuha Kamran, Mujahid Manzoor, and et al. 2022. "Genome-Wide Identification and In Silico Analysis of ZF-HD Transcription Factor Genes in Zea mays L." Genes 13, no. 11: 2112. https://doi.org/10.3390/genes13112112
APA StyleIslam, M. A. U., Nupur, J. A., Khalid, M. H. B., Din, A. M. U., Shafiq, M., Alshegaihi, R. M., Ali, Q., Ali, Q., Kamran, Z., Manzoor, M., Haider, M. S., Shahid, M. A., & Manghwar, H. (2022). Genome-Wide Identification and In Silico Analysis of ZF-HD Transcription Factor Genes in Zea mays L. Genes, 13(11), 2112. https://doi.org/10.3390/genes13112112