
Genetic Dissection of Temperament Personality Traits in Italian Isolates

 , , , ,
, , , , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Description of the Sample
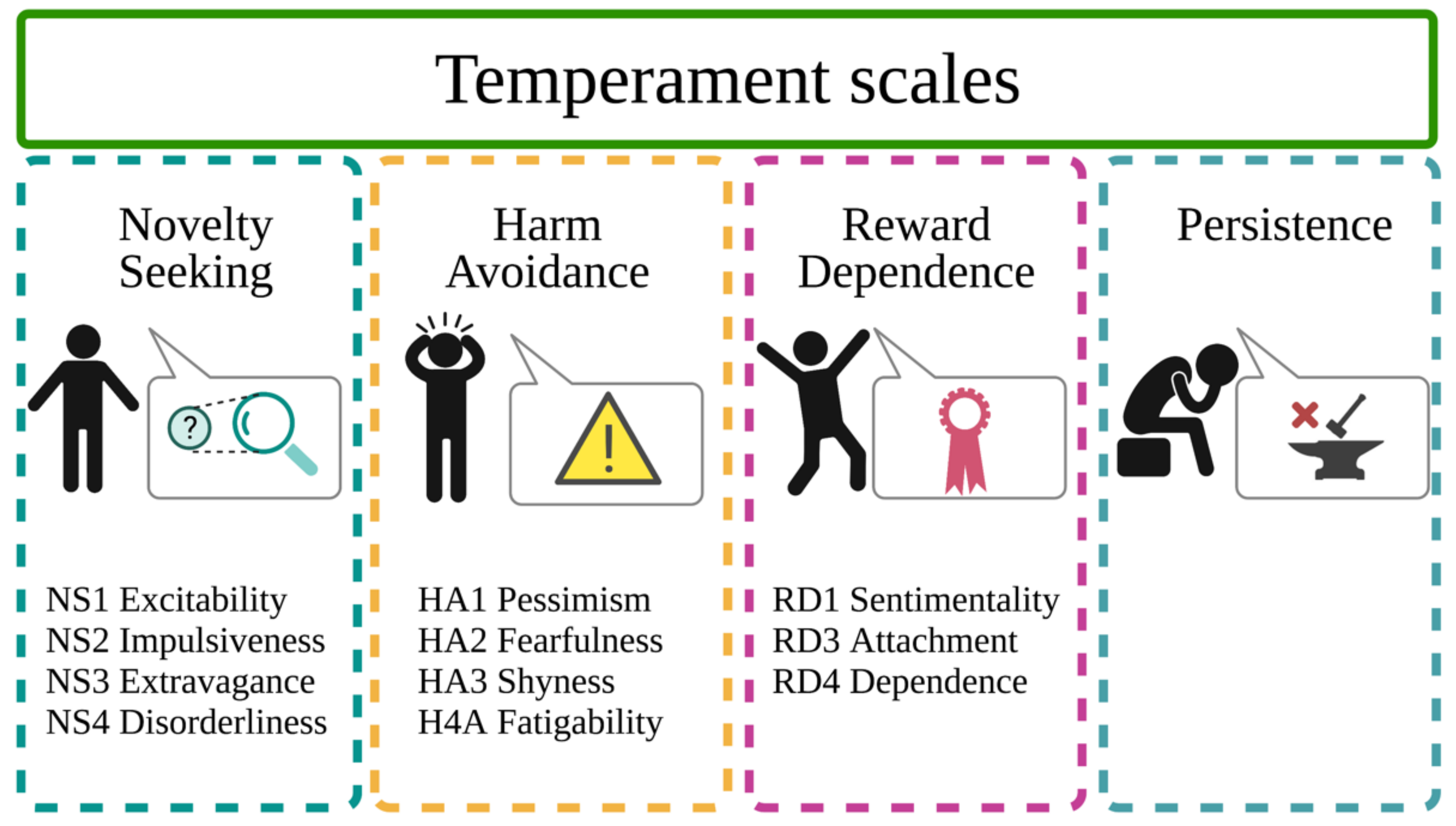
2.2. Personality Assessment
- -
- NS is measured using 40 items; of these, 11 for Excitability, 10 for Impulsiveness, 10 for Extravagance, and nine for Disorderliness;
- -
- HA is measured using 35 items; of these, 11 for Pessimism (anticipatory worry), seven for Fearfulness (fear of uncertainty), eight for Shyness, and nine for Fatigability;
- -
- RD is measured using 24 items; of these, 10 for Sentimentality, 8 for Attachment, 6 for Dependence (on approval of others);
- -
- P is measured using eight items.
2.3. Anxiety and Depression Evaluation
2.4. Inter-Correlations between the Temperament Scales and Associated Variables
2.5. Genotyping and Imputation
2.6. Genome-Wide Association Study
2.7. Expression Level Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cloninger, C.R.; Cloninger, K.M.; Zwir, I.; Keltikangas-Järvinen, L. The complex genetics and biology of human temperament: A review of traditional concepts in relation to new molecular findings. Transl. Psychiatry 2019, 9, 1–21. [Google Scholar] [CrossRef]
- Zwir, I.; Arnedo, J.; Del-Val, C.; Pulkki-Råback, L.; Konte, B.; Yang, S.S.; Romero-Zaliz, R.; Hintsanen, M.; Cloninger, K.M.; Garcia, D.; et al. Uncovering the complex genetics of human temperament. Mol. Psychiatry 2020, 25, 2275–2294. [Google Scholar] [CrossRef] [PubMed]
- Zwir, I.; Arnedo, J.; Del-Val, C.; Pulkki-Råback, L.; Konte, B.; Yang, S.S.; Romero-Zaliz, R.; Hintsanen, M.; Cloninger, K.M.; Garcia, D.; et al. Uncovering the complex genetics of human character. Mol. Psychiatry 2020, 25, 2295–2312. [Google Scholar] [CrossRef] [PubMed]
- Frazer, K.A.; Murray, S.S.; Schork, N.J.; Topol, E.J. Human genetic variation and its contribution to complex traits. Nat. Rev. Genet. 2009, 10, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Cloninger, C.R.; Svrakic, D.M.; Przybeck, T.R. A Psychobiological Model of Temperament and Character. Arch. Gen. Psychiatry 1993, 50, 975–990. [Google Scholar] [CrossRef]
- Vuijk, R.; de Nijs, P.F.; Deen, M.; Vitale, S.; Simons-Sprong, M.; Hengeveld, M.W. Temperament and character in men with autism spectrum disorder: A reanalysis of scores on the Temperament and Character Inventory by individual case matching. Contemp. Clin. Trials Commun. 2018, 12, 55–59. [Google Scholar] [CrossRef]
- Kantojärvi, L.; Miettunen, J.; Veijola, J.; Läksy, K.; Karvonen, J.T.; Ekelund, J.; Järvelin, M.-R.; Lichtermann, D.; Joukamaa, M. Temperament profiles in personality disorders among a young adult population. Nord. J. Psychiatry 2008, 62, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Luca, A.; Nicoletti, A.; Mostile, G.; Sciacca, G.; Dibilio, V.; Cicero, C.E.; Raciti, L.; Donzuso, G.; Zappia, M. Temperament traits and executive functions in Parkinson’s disease. Neurosci. Lett. 2018, 684, 25–28. [Google Scholar] [CrossRef] [PubMed]
- Hettema, J.M.; Neale, M.C.; Myers, J.M.; Prescott, C.A.; Kendler, K.S. A Population-Based Twin Study of the Relationship Between Neuroticism and Internalizing Disorders. Am. J. Psychiatry 2006, 163, 857–864. [Google Scholar] [CrossRef]
- Esko, T.; Mezzavilla, M.; Nelis, M.; Borel, C.; Debniak, T.; Jakkula, E.; Julia, A.; Karachanak, S.; Khrunin, A.; Kisfali, P.; et al. Genetic characterization of northeastern Italian population isolates in the context of broader European genetic diversity. Eur. J. Hum. Genet. 2012, 21, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Mezzavilla, M.; Haber, M.; McCarthy, S.; Chen, Y.; Narasimhan, V.; Gilly, A.; Ayub, Q.; Colonna, V.; Southam, L.; et al. Enrichment of low-frequency functional variants revealed by whole-genome sequencing of multiple isolated European populations. Nat. Commun. 2017, 8, 15927. [Google Scholar] [CrossRef] [PubMed]
- Cocca, M.; Barbieri, C.; Concas, M.P.; Robino, A.; Brumat, M.; Gandin, I.; Trudu, M.; Sala, C.F.; Vuckovic, D.; Girotto, G.; et al. A bird’s-eye view of Italian genomic variation through whole-genome sequencing. Eur. J. Hum. Genet. 2019, 28, 435–444. [Google Scholar] [CrossRef] [PubMed]
- Howie, B.N.; Donnelly, P.; Marchini, J. A Flexible and Accurate Genotype Imputation Method for the Next Generation of Genome-Wide Association Studies. PLoS Genet. 2009, 5, e1000529. [Google Scholar] [CrossRef]
- Belonogova, N.M.; Svishcheva, G.R.; Van Duijn, C.M.; Aulchenko, Y.S.; Axenovich, T.I. Region-Based Association Analysis of Human Quantitative Traits in Related Individuals. PLoS ONE 2013, 8, e65395. [Google Scholar] [CrossRef] [PubMed]
- Aulchenko, Y.S.; Ripke, S.; Isaacs, A.; Van Duijn, C.M. GenABEL: An R library for genome-wide association analysis. Bioinformatics 2007, 23, 1294–1296. [Google Scholar] [CrossRef]
- Miettunen, J.; Lauronen, E.; Kantojärvi, L.; Veijola, J.; Joukamaa, M. Inter-correlations between Cloninger’s temperament dimensions—A meta-analysis. Psychiatry Res. 2008, 160, 106–114. [Google Scholar] [CrossRef]
- Jiang, R.; Calhoun, V.D.; Zuo, N.; Lin, D.; Li, J.; Fan, L.; Qi, S.; Sun, H.; Fu, Z.; Song, M.; et al. Connectome-based individualized prediction of temperament trait scores. NeuroImage 2018, 183, 366–374. [Google Scholar] [CrossRef] [PubMed]
- McLaren, W.; Gil, L.; Hunt, S.E.; Riat, H.S.; Ritchie, G.R.S.; Thormann, A.; Flicek, P.; Cunningham, F. The Ensembl Variant Effect Predictor. Genome Biol. 2016, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- The GTEx Consortium; Ardlie, K.G.; DeLuca, D.S.; Segre, A.V.; Sullivan, T.J.; Young, T.R.; Gelfand, E.T.; Trowbridge, C.A.; Maller, J.B.; Tukiainen, T.; et al. The Genotype-Tissue Expression (GTEx) pilot analysis: Multitissue gene regulation in humans. Science 2015, 348, 648–660. [Google Scholar] [CrossRef]
- Ongen, H.; Buil, A.; Brown, A.; Dermitzakis, E.T.; Delaneau, O. Fast and efficient QTL mapper for thousands of molecular phenotypes. Bioinformatics 2015, 32, 1479–1485. [Google Scholar] [CrossRef]
- Storey, J.D.; Tibshirani, R. Statistical significance for genomewide studies. Proc. Natl. Acad. Sci. USA 2003, 100, 9440–9445. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.M.; Skaar, S.; Danielson, E.; Lee, S.H. Aberrant expression of S-SCAM causes the loss of GABAergic synapses in hippocampal neurons. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Shen, L.; Thompson, P.M.; Potkin, S.G.; Bertram, L.; Farrer, L.A.; Foroud, T.M.; Green, R.C.; Hu, X.; Huentelman, M.J.; Kim, S.; et al. Genetic Analysis of Quantitative Phenotypes in AD and MCI: Imaging, Cognition and Biomarkers. Brain Imaging Behav. 2014, 8, 183–207. [Google Scholar] [CrossRef] [PubMed]
- Ochi, S.; Iga, J.-I.; Funahashi, Y.; Yoshino, Y.; Yamazaki, K.; Kumon, H.; Mori, H.; Ozaki, Y.; Mori, T.; Ueno, S.-I. Identifying Blood Transcriptome Biomarkers of Alzheimer’s Disease Using Transgenic Mice. Mol. Neurobiol. 2020, 57, 4941–4951. [Google Scholar] [CrossRef] [PubMed]
- Koide, T.; Banno, M.; Aleksic, B.; Yamashita, S.; Kikuchi, T.; Kohmura, K.; Adachi, Y.; Kawano, N.; Kushima, I.; Nakamura, Y.; et al. Common Variants in MAGI2 Gene Are Associated with Increased Risk for Cognitive Impairment in Schizophrenic Patients. PLoS ONE 2012, 7, e36836. [Google Scholar] [CrossRef]
- Sahoo, T.; Theisen, A.; Rosenfeld, J.A.; Lamb, A.N.; Ravnan, J.B.; Schultz, R.A.; Torchia, B.S.; Neill, N.; Casci, I.; Bejjani, B.A.; et al. Copy number variants of schizophrenia susceptibility loci are associated with a spectrum of speech and developmental delays and behavior problems. Genet. Med. 2011, 13, 868–880. [Google Scholar] [CrossRef] [PubMed]
- Saetre, P.; Lindberg, J.; Leonard, J.; Olsson, K.; Pettersson, U.; Ellegren, H.; Bergström, T.; Vilà, C.; Jazin, E. From wild wolf to domestic dog: Gene expression changes in the brain. Mol. Brain Res. 2004, 126, 198–206. [Google Scholar] [CrossRef] [PubMed]
- Mathé, A.A.; Ågren, H.; Lindström, L.; Theodorsson, E. Increased concentration of calcitonin gene-related peptide in cerebrospinal fluid of depressed patients. A possible trait marker of major depressive disorder. Neurosci. Lett. 1994, 182, 138–142. [Google Scholar] [CrossRef]
- Matsui, A.; Tran, M.; Yoshida, A.C.; Kikuchi, S.S.; Mami, U.; Ogawa, M.; Shimogori, T. BTBD3 Controls Dendrite Orientation Toward Active Axons in Mammalian Neocortex. Science 2013, 342, 1114–1118. [Google Scholar] [CrossRef]
- Stewart, S.E.; Yu, D.; Scharf, J.M.; Neale, B.M.; Fagerness, J.A.; Mathews, C.A.; Arnold, P.D.; Evans, P.D.; Gamazon, E.R.; Osiecki, L.; et al. Genome-wide association study of obsessive-compulsive disorder. Mol. Psychiatry 2013, 18, 788–798. [Google Scholar] [CrossRef] [PubMed]
- Yokota, M.; Kakuta, S.; Shiga, T.; Ishikawa, K.-I.; Okano, H.; Hattori, N.; Akamatsu, W.; Koike, M. Establishment of an in vitro model for analyzing mitochondrial ultrastructure in PRKN-mutated patient iPSC-derived dopaminergic neurons. Mol. Brain 2021, 14, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Milanowski, Ł.M.; Lindemann, J.A.; Hoffman-Zacharska, D.; Soto-Beasley, A.I.; Barcikowska, M.; Boczarska-Jedynak, M.; Deutschlander, A.; Kłodowska, G.; Dulski, J.; Fedoryshyn, L.; et al. Frequency of mutations in PRKN, PINK1, and DJ1 in Patients With Early-Onset Parkinson Disease from neighboring countries in Central Europe. Park. Relat. Disord. 2021, 86, 48–51. [Google Scholar] [CrossRef] [PubMed]
- Lubbe, S.J.; I Bustos, B.; Hu, J.; Krainc, D.; Joseph, T.; Hehir, J.; Tan, M.; Zhang, W.; Escott-Price, V.; Williams, N.M.; et al. Assessing the relationship between monoallelic PRKN mutations and Parkinson’s risk. Hum. Mol. Genet. 2021, 30, 78–86. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Phenotype | Nearest Gene (Protein Coding) | Chr | Start Position | End Position | Top SNP | p-Value | Number of SNPs with p-Value < 1 × 10−5 |
|---|---|---|---|---|---|---|---|
| NS3 (Extravagance) | MAGI2 | 7 | 77646393 | 79082890 | rs13242288 | 9.14 × 10−8 | 24 |
| NS4 (Disorderliness) | CALCB | 11 | 14926543 | 15103888 | rs10832317 | 4.56 × 10−6 | 81 |
| HA4 (Fatigability) | BTBD3 | 20 | 11871371 | 11907257 | rs4814036 | 2.15 × 10−8 | 72 |
| RD (Reward Dependence) | PARK2 | 6 | 161768452 | 163148803 | rs10455897 | 8.27 × 10−9 | 27 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Concas, M.P.; Minelli, A.; Aere, S.; Morgan, A.; Tesolin, P.; Gasparini, P.; Gennarelli, M.; Girotto, G. Genetic Dissection of Temperament Personality Traits in Italian Isolates. Genes 2022, 13, 4. https://doi.org/10.3390/genes13010004
Concas MP, Minelli A, Aere S, Morgan A, Tesolin P, Gasparini P, Gennarelli M, Girotto G. Genetic Dissection of Temperament Personality Traits in Italian Isolates. Genes. 2022; 13(1):4. https://doi.org/10.3390/genes13010004
Chicago/Turabian StyleConcas, Maria Pina, Alessandra Minelli, Susanna Aere, Anna Morgan, Paola Tesolin, Paolo Gasparini, Massimo Gennarelli, and Giorgia Girotto. 2022. "Genetic Dissection of Temperament Personality Traits in Italian Isolates" Genes 13, no. 1: 4. https://doi.org/10.3390/genes13010004
APA StyleConcas, M. P., Minelli, A., Aere, S., Morgan, A., Tesolin, P., Gasparini, P., Gennarelli, M., & Girotto, G. (2022). Genetic Dissection of Temperament Personality Traits in Italian Isolates. Genes, 13(1), 4. https://doi.org/10.3390/genes13010004