
The Mitochondrial DNA Landscape of Modern Mexico

 , , ,
, , ,  and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Collection
2.2. MtDNA Sequencing, Haplogroup Estimation, and Quality Control
2.3. Forensic, Population Genetic, and Phylogeographic Calculations
3. Results
3.1. Quality Control
3.2. Forensic and Population Genetic Characterization of the Mexican mtDNA Dataset
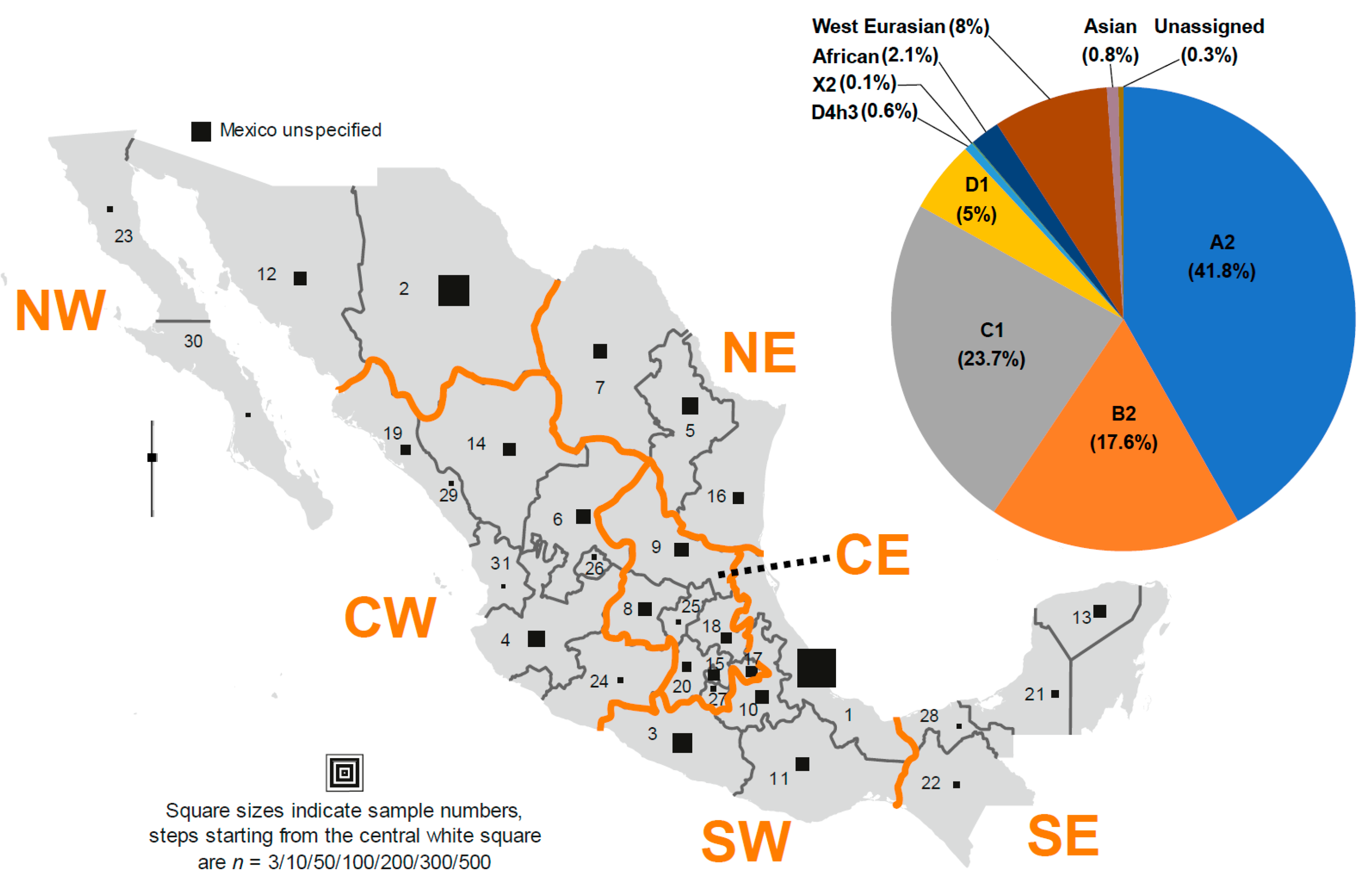
3.3. The mtDNA Haplogroups of Mexico
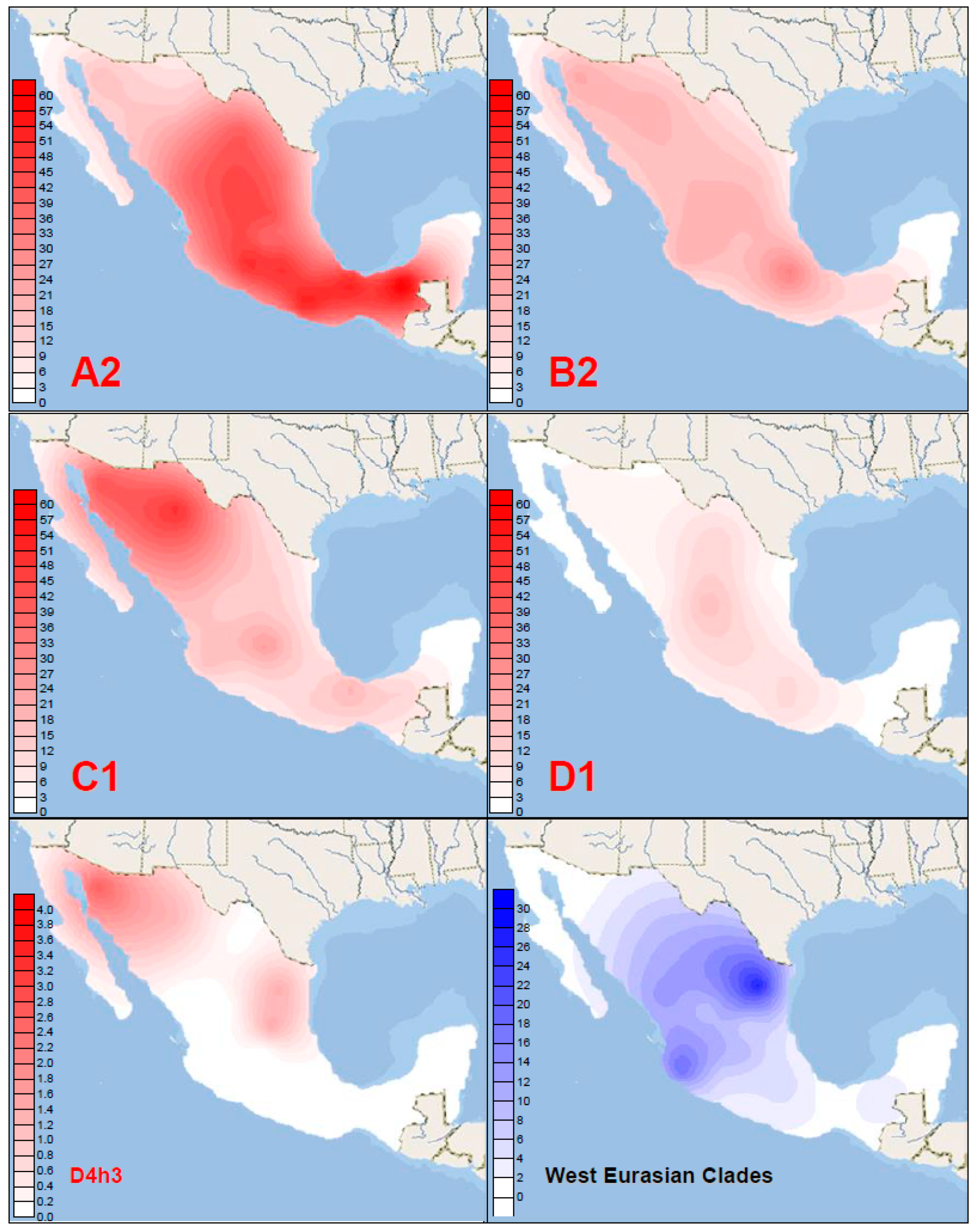
3.4. The Phylogeography of mtDNA Haplogroups in Mexico
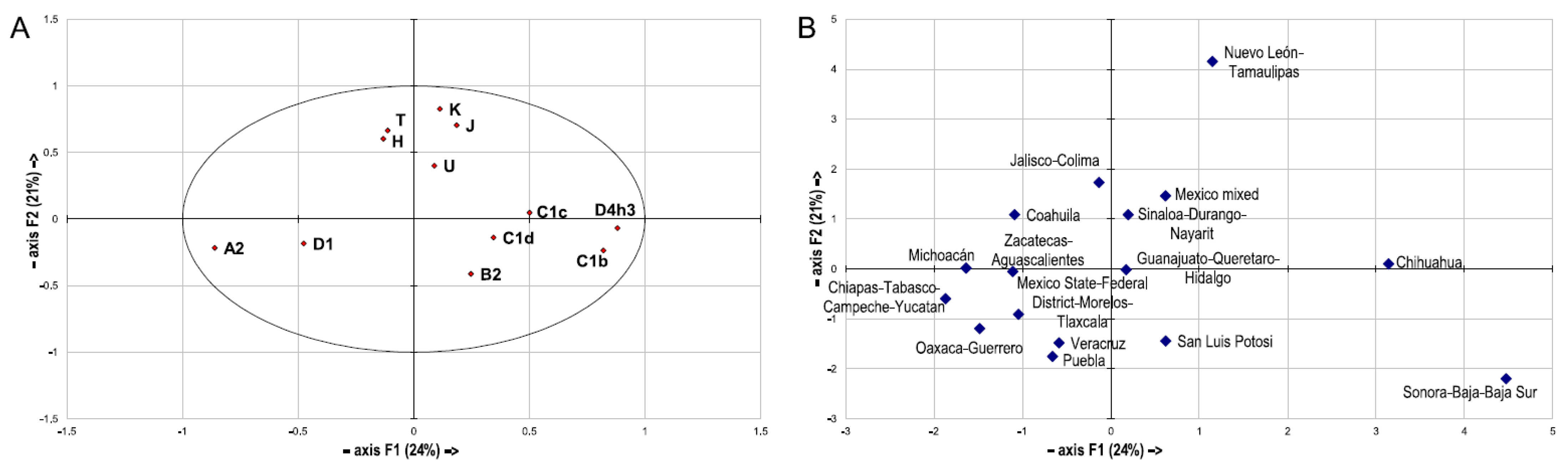
3.5. Regional mtDNA Databases for Mexico?
3.6. The Sex-Biased Genetic History of Mexico
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- UN Department of Economic and Social Affairs Statistics Division. Standard Country or Area Codes for Statistical Use; United Nations: New York, NY, USA, 1999. [Google Scholar]
- Kemp, B.M.; Gonzalez-Oliver, A.; Malhi, R.S.; Monroe, C.; Schroeder, K.B.; McDonough, J.; Rhett, G.; Resendez, A.; Penaloza-Espinosa, R.I.; Buentello-Malo, L.; et al. Evaluating the farming/language dispersal hypothesis with genetic variation exhibited by populations in the Southwest and Mesoamerica. Proc. Natl. Acad. Sci. USA 2010, 107, 6759–6764. [Google Scholar] [CrossRef]
- Gorostiza, A.; Acunha-Alonzo, V.; Regalado-Liu, L.; Tirado, S.; Granados, J.; Samano, D.; Rangel-Villalobos, H.; Gonzalez-Martin, A. Reconstructing the history of Mesoamerican populations through the study of the mitochondrial DNA control region. PLoS ONE 2012, 7, e44666. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, F.; Gojobori, J.; Wang, L.; Onishi, K.; Sugiyama, S.; Granados, J.; Gomez-Trejo, C.; Acuna-Alonzo, V.; Ueda, S. Complete mitogenome analysis of indigenous populations in Mexico: Its relevance for the origin of Mesoamericans. J. Hum. Genet. 2014, 59, 359–367. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Martin, A.; Gorostiza, A.; Regalado-Liu, L.; Arroyo-Pena, S.; Tirado, S.; Nuno-Arana, I.; Rubi-Castellanos, R.; Sandoval, K.; Coble, M.D.; Rangel-Villalobos, H. Demographic history of indigenous populations in Mesoamerica based on mtDNA sequence data. PLoS ONE 2015, 10, e0131791. [Google Scholar]
- Morales-Arce, A.Y.; Hofman, C.A.; Duggan, A.T.; Benfer, A.K.; Katzenberg, M.A.; McCafferty, G.; Warinner, C. Successful reconstruction of whole mitochondrial genomes from ancient Central America and Mexico. Sci. Rep. 2017, 7, 18100. [Google Scholar] [CrossRef]
- Central Intelligence Agency: The World Factbook 2021; Central Intelligence Agency: Washington, DC, USA, 2021.
- Schurr, T.G. The peopling of the New World: Perspectives from molecular anthropology. Annu. Rev. Anthropol. 2004, 33, 551–583. [Google Scholar] [CrossRef]
- Bodner, M.; Perego, U.A.; Huber, G.; Fendt, L.; Rock, A.W.; Zimmermann, B.; Olivieri, A.; Gomez-Carballa, A.; Lancioni, H.; Angerhofer, N.; et al. Rapid coastal spread of first americans: Novel insights from South America’s Southern Cone mitochondrial genomes. Genome Res. 2012, 22, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Sandoval, K.; Buentello-Malo, L.; Penaloza-Espinosa, R.; Avelino, H.; Salas, A.; Calafell, F.; Comas, D. Linguistic and maternal genetic diversity are not correlated in native Mexicans. Hum. Genet. 2009, 126, 521–531. [Google Scholar] [CrossRef][Green Version]
- Chatters, J.C.; Kennett, D.J.; Asmerom, Y.; Kemp, B.M.; Polyak, V.; Blank, A.N.; Beddows, P.A.; Reinhardt, E.; Arroyo-Cabrales, J.; Bolnick, D.A.; et al. Late pleistocene human skeleton and mtDNA link Paleoamericans and modern Native Americans. Science 2014, 344, 750–754. [Google Scholar] [CrossRef] [PubMed]
- Capodiferro, M.R.; Aram, B.; Raveane, A.; Migliore, N.R.; Colombo, G.; Ongaro, L.; Rivera, J.; Mendizabal, T.; Hernandez-Mora, I.; Tribaldos, M.; et al. Archaeogenomic distinctiveness of the Isthmo-Colombian area. Cell 2021, 184, 1706–1723. [Google Scholar] [CrossRef]
- Guardado-Estrada, M.; Juarez-Torres, E.; Medina-Martinez, I.; Wegier, A.; Macias, A.; Gomez, G.; Cruz-Talonia, F.; Roman-Bassaure, E.; Pinero, D.; Kofman-Alfaro, S.; et al. A great diversity of Amerindian mitochondrial DNA ancestry is present in the Mexican Mestizo population. J. Hum. Genet. 2009, 54, 695–705. [Google Scholar] [CrossRef]
- Gonzalez-Sobrino, B.Z.; Pintado-Cortina, A.P.; Sebastian-Medina, L.; Morales-Mandujano, F.; Contreras, A.V.; Aguilar, Y.E.; Chavez-Benavides, J.; Carrillo-Rodriguez, A.; Silva-Zolezzi, I.; Medrano-Gonzalez, L. Genetic diversity and differentiation in urban and indigenous populations of Mexico: Patterns of mitochondrial DNA and Y-chromosome lineages. Biodemogr. Soc. Biol. 2016, 62, 53–72. [Google Scholar] [CrossRef]
- Gojobori, J.; Mizuno, F.; Wang, L.; Onishi, K.; Granados, J.; Gomez-Trejo, C.; Acuna-Alonzo, V.; Ueda, S. MtDNA diversity of the Zapotec in Mexico suggests a population decline long before the first contact with Europeans. J. Hum. Genet. 2015, 60, 557–559. [Google Scholar] [CrossRef]
- Green, L.D.; Derr, J.N.; Knight, A. MtDNA affinities of the peoples of North-Central Mexico. Am. J. Hum. Genet. 2000, 66, 989–998. [Google Scholar] [CrossRef]
- Gonzalez-Oliver, A.; Pineda-Vazquez, D.; Garfias-Morales, E.; La Cruz-Laina, I.; Medrano-Gonzalez, L.; Marquez-Morfin, L.; Ortega-Munoz, A. Genetic overview of the Maya populations: Mitochondrial DNA haplogroups. Hum. Biol. 2018, 90, 281–300. [Google Scholar] [CrossRef]
- Martinez-Cortes, G.; Salazar-Flores, J.; Haro-Guerrero, J.; Rubi-Castellanos, R.; Velarde-Felix, J.S.; Munoz-Valle, J.F.; Lopez-Casamichana, M.; Carrillo-Tapia, E.; Canseco-Avila, L.M.; Bravi, C.M.; et al. Maternal admixture and population structure in Mexican-Mestizos based on mtDNA haplogroups. Am. J. Phys. Anthropol. 2013, 151, 526–537. [Google Scholar] [CrossRef]
- Vidal, O.; Brusca, R.C. Mexico’s biocultural diversity in peril. Rev. Biol. Trop. 2020, 68, 669–691. [Google Scholar] [CrossRef]
- Kumar, S.; Bellis, C.; Zlojutro, M.; Melton, P.E.; Blangero, J.; Curran, J.E. Large scale mitochondrial sequencing in Mexican Americans suggests a reappraisal of Native American origins. BMC Evol. Biol 2011, 11, 293. [Google Scholar] [CrossRef]
- Mitchell, S.L.; Goodloe, R.; Brown-Gentry, K.; Pendergrass, S.A.; Murdock, D.G.; Crawford, D.C. Characterization of mitochondrial haplogroups in a large population-based sample from the United States. Hum. Genet. 2014, 133, 861–868. [Google Scholar] [CrossRef] [PubMed]
- Gorodezky, C.; Alaez, C.; Vazquez-Garcia, M.N.; de la Rosa, G.; Infante, E.; Balladares, S.; Toribio, R.; Perez-Luque, E.; Munoz, L. The genetic structure of Mexican Mestizos of different locations: Tracking back their origins through MHC genes, blood group systems, and microsatellites. Hum. Immunol. 2001, 62, 979–991. [Google Scholar] [CrossRef]
- Salas, A.; Acosta, A.; Alvarez-Iglesias, V.; Cerezo, M.; Phillips, C.; Lareu, M.V.; Carracedo, A. The mtdna ancestry of admixed Colombian populations. Am. J. Hum. Biol. 2008, 20, 584–591. [Google Scholar] [CrossRef]
- Instituto Nacional de Estadística y Geografía (México). Panorama Sociodemográfico de México—Censo de Población y Vivienda 2020; INEGI: Aguascalientes, México, 2021. [Google Scholar]
- Instituto Nacional de Estadística y Geografía (México). Clasificación de Lenguas Indígenas 2010; INEGI: Aguascalientes, México, 2016. [Google Scholar]
- Stoneking, M.; Hedgecock, D.; Higuchi, R.G.; Vigilant, L.; Erlich, H.A. Population variation of human mtDNA control region sequences detected by enzymatic amplification and sequence-specific oligonucleotide probes. Am. J. Hum. Genet. 1991, 48, 370–382. [Google Scholar]
- Horai, S.; Kondo, R.; Nakagawahattori, Y.; Hayashi, S.; Sonoda, S.; Tajima, K. Peopling of the america, founded by 4 major lineages of mitochondrial DNA. Mol. Biol. Evol. 1993, 10, 23–47. [Google Scholar]
- Torroni, A.; Schurr, T.G.; Cabell, M.F.; Brown, M.D.; Neel, J.V.; Larsen, M.; Smith, D.G.; Vullo, C.M.; Wallace, D.C. Asian affinities and continental radiation of the 4 founding native-american mtDNAs. Am. J. Hum. Genet. 1993, 53, 563–590. [Google Scholar] [PubMed]
- Torroni, A.; Chen, Y.S.; Semino, O.; Santachiarabeneceretti, A.S.; Scott, C.R.; Lott, M.T.; Winter, M.; Wallace, D.C. MtDNA and Y-chromosome polymorphisms in 4 native-american populations from Southern Mexico. Am. J. Hum. Genet. 1994, 54, 303–318. [Google Scholar]
- Bonilla, C.; Gutierrez, G.; Parra, E.J.; Kline, C.; Shriver, M.D. Admixture analysis of a rural population of the state of Guerrero, Mexico. Am. J. Phys. Anthropol. 2005, 128, 861–869. [Google Scholar] [CrossRef]
- Rosas, R.C.B.; Sesma, A.M.; Ortega, L.H.; Gonzalez, L.H.; Avalos, J.V.; Cruz, A.A.A. The utility of genomic public databases to mitochondrial haplotyping in contemporary Mestizo population of Mexican origin. Mitochondrial DNA A 2019, 30, 567–572. [Google Scholar] [CrossRef] [PubMed]
- Parra, J.C.V.; Rosas, R.C.B.; Leon, D.C.Y. Inequality in medical-genetic research towards minority groups an approach to the case of people of African descent in Mexico. Res. J. Med. Sci. 2018, 12, 46–51. [Google Scholar]
- Avila-Arcos, M.C.; McManus, K.F.; Sandoval, K.; Rodriguez-Rodriguez, J.E.; Villa-Islas, V.; Martin, A.R.; Luisi, P.; Penaloza-Espinosa, R.I.; Eng, C.; Huntsman, S.; et al. Population history and gene divergence in Native Mexicans inferred from 76 human exomes. Mol. Biol. Evol. 2020, 37, 994–1006. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Ordonez, I.; Perez-Villatoro, F.; Garcia-Ortiz, H.; Barajas-Olmos, F.; Ballesteros-Villascan, J.; Gonzalez-Buenfil, R.; Fresno, C.; Garciarrubio, A.; Fernandez-Lopez, J.C.; Tovar, H.; et al. Whole genome variation in 27 Mexican indigenous populations, demographic and biomedical insights. PLoS ONE 2021, 16, e0249773. [Google Scholar] [CrossRef] [PubMed]
- Campos-Sanchez, R.; Barrantes, R.; Silva, S.; Escamilla, M.; Ontiveros, A.; Nicolini, H.; Mendoza, R.; Munoz, R.; Raventos, H. Genetic structure analysis of three Hispanic populations from Costa Rica, Mexico, and the Southwestern United States using Y-chromosome STR markers and mtDNA sequences. Hum. Biol. 2006, 78, 551–563. [Google Scholar] [CrossRef]
- Gonzalez-Oliver, A.; Garfias-Morales, E.; Smith, D.G.; Quinto-Sanchez, M. Mitochondrial DNA analysis of Mazahua and Otomi indigenous populations from Estado de Mexico suggests a distant common ancestry. Hum. Biol. 2017, 89, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Parson, W.; Gusmao, L.; Hares, D.R.; Irwin, J.A.; Mayr, W.R.; Morling, N.; Pokorak, E.; Prinz, M.; Salas, A.; Schneider, P.M.; et al. DNA commission of the International Society for Forensic Genetics: Revised and extended guidelines for mitochondrial DNA typing. Forensic Sci. Int. Genet. 2014, 13, 134–142. [Google Scholar] [CrossRef] [PubMed]
- Kemp, B.M.; Reséndez, A.; Berrelleza, J.A.R.B.; Malhi, R.S.; Smith, D.G. An analysis of ancient aztec mtdna from tlatelolco: Pre-Columbian relations and the spread of Uto-Aztecan. In Biomolecular Archaeology: Genetic Approaches to the Past; Reed, D.M., Ed.; Center for Archaeological Investigation, Southern Illinois University: Carbondale, IL, USA, 2005. [Google Scholar]
- Alvarez-Sandoval, B.A.; Manzanilla, L.R.; Gonzalez-Ruiz, M.; Malgosa, A.; Montiel, R. Genetic evidence supports the multiethnic character of Teopancazco, a neighborhood center of Teotihuacan, Mexico (AD 200-600). PLoS ONE 2015, 10, e0132371. [Google Scholar] [CrossRef] [PubMed]
- Watkins, W.S.; Xing, J.C.; Huff, C.; Witherspoon, D.J.; Zhang, Y.H.; Perego, U.A.; Woodward, S.R.; Jorde, L.B. Genetic analysis of ancestry, admixture and selection in Bolivian and Totonac populations of the New World. BMC Genet. 2012, 13, 39. [Google Scholar] [CrossRef]
- Cardoso, S.; Alfonso-Sanchez, M.A.; Valverde, L.; Sanchez, D.; Zarrabeitia, M.T.; Odriozola, A.; Martinez-Jarreta, B.; de Pancorbo, M.M. Genetic uniqueness of the Waorani tribe from the Ecuadorian Amazon. Heredity 2012, 108, 609–615. [Google Scholar] [CrossRef]
- Mizuno, F.; Kumagai, M.; Kurosaki, K.; Hayashi, M.; Sugiyama, S.; Ueda, S.; Wang, L. Imputation approach for deducing a complete mitogenome sequence from low-depth-coverage next-generation sequencing data: Application to ancient remains from the Moon pyramid, Mexico. J. Hum. Genet. 2017, 62, 631–635. [Google Scholar] [CrossRef]
- Migration Data Portal: Migrant Deaths and Disappearances. Available online: https://migrationdataportal.org/themes/migrant-deaths-and-disappearances (accessed on 16 August 2021).
- Perego, U.A.; Lancioni, H.; Tribaldos, M.; Angerhofer, N.; Ekins, J.E.; Olivieri, A.; Woodward, S.R.; Pascale, J.M.; Cooke, R.; Motta, J.; et al. Decrypting the mitochondrial gene pool of modern Panamanians. PLoS ONE 2012, 7, e38337. [Google Scholar] [CrossRef]
- Andrews, R.M.; Kubacka, I.; Chinnery, P.F.; Lightowlers, R.N.; Turnbull, D.M.; Howell, N. Reanalysis and revision of the Cambridge Reference Sequence for human mitochondrial DNA. Nat. Genet. 1999, 23, 147. [Google Scholar] [CrossRef]
- Parson, W.; Dür, A. EMPOP-a forensic mtDNA database. Forensic Sci. Int. Genet. 2007, 1, 88–92. [Google Scholar] [CrossRef]
- Huber, N.; Parson, W.; Dür, A. Next generation database search algorithm for forensic mitogenome analyses. Forensic Sci. Int. Genet. 2018, 37, 204–214. [Google Scholar] [CrossRef]
- Van Oven, M.; Kayser, M. Updated comprehensive phylogenetic tree of global human mitochondrial DNA variation. Hum. Mutat. 2009, 30, E386–E394. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef]
- Kiesler, K.M.; Coble, M.D.; Hall, T.A.; Vallone, P.M. Comparison of base composition analysis and sanger sequencing of mitochondrial DNA for four U.S. Population groups. Forensic Sci. Int. Genet. 2014, 8, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Modi, A.; Lancioni, H.; Cardinali, I.; Capodiferro, M.R.; Rambaldi Migliore, N.; Hussein, A.; Strobl, C.; Bodner, M.; Schnaller, L.; Xavier, C.; et al. The mitogenome portrait of Umbria in Central Italy as depicted by contemporary inhabitants and pre-Roman remains. Sci. Rep. 2020, 10, 10700. [Google Scholar] [CrossRef] [PubMed]
- Olivieri, A.; Sidore, C.; Achilli, A.; Angius, A.; Posth, C.; Furtwangler, A.; Brandini, S.; Capodiferro, M.R.; Gandini, F.; Zoledziewska, M.; et al. Mitogenome diversity in Sardinians: A genetic window onto an island’s past. Mol. Biol. Evol. 2017, 34, 1230–1239. [Google Scholar] [CrossRef]
- Perego, U.A.; Angerhofer, N.; Pala, M.; Olivieri, A.; Lancioni, H.; Hooshiar Kashani, B.; Carossa, V.; Ekins, J.E.; Gomez-Carballa, A.; Huber, G.; et al. The initial peopling of the Americas: A growing number of founding mitochondrial genomes from Beringia. Genome Res. 2010, 20, 1174–1179. [Google Scholar] [CrossRef] [PubMed]
- Perego, U.A.; Achilli, A.; Angerhofer, N.; Accetturo, M.; Pala, M.; Olivieri, A.; Hooshiar Kashani, B.; Ritchie, K.H.; Scozzari, R.; Kong, Q.P.; et al. Distinctive paleo-indian migration routes from Beringia marked by two rare mtDNA haplogroups. Curr. Biol. 2009, 19, 1–8. [Google Scholar] [CrossRef]
- Bodner, M.; Iuvaro, A.; Strobl, C.; Nagl, S.; Huber, G.; Pelotti, S.; Pettener, D.; Luiselli, D.; Parson, W. Helena, the hidden beauty: Resolving the most common West Eurasian mtdna control region haplotype by massively parallel sequencing an Italian population sample. Forensic Sci. Int. Genet. 2015, 15, 21–26. [Google Scholar] [CrossRef]
- Lorenz, J.G.; Smith, D.G. Distribution of four founding mtDNA haplogroups among Native North Americans. Am. J. Phys. Anthropol. 1996, 101, 307–323. [Google Scholar] [CrossRef]
- Penaloza-Espinosa, R.I.; Arenas-Aranda, D.; Cerda-Flores, R.M.; Buentello-Malo, L.; Gonzalez-Valencia, G.; Torres, J.; Alvarez, B.; Mendoza, I.; Flores, M.; Sandoval, L.; et al. Characterization of mtDNA haplogroups in 14 Mexican indigenous populations. Hum. Biol. 2007, 79, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Boiso, A.; Penaloza-Espinosa, R.I.; Castro-Sierra, E.; Cerda-Flores, R.M.; Buentello-Malo, L.; Sanchez-Urbina, R.; Ortiz-de-Luna, R.I.; Rodriguez-Espino, B.A.; Salamanca-Gomez, F.A.; Flores-Ayon, M.P.; et al. Genetic structure of three native Mexican communities based on mtDNA haplogroups, and ABO and Rh blood group systems. Rev. Investig. Clin. 2011, 63, 614–620. [Google Scholar]
- Gonzalez-Oliver, A.; Marquez-Morfin, L.; Jimenez, J.C.; Torre-Blanco, A. Founding amerindian mitochondrial DNA lineages in ancient Maya from Xcaret, Quintana Roo. Am. J. Phys. Anthropol. 2001, 116, 230–235. [Google Scholar] [CrossRef] [PubMed]
- Mata-Miguez, J.; Overholtzer, L.; Rodriguez-Alegria, E.; Kemp, B.M.; Bolnick, D.A. The genetic impact of Aztec imperialism: Ancient mitochondrial DNA evidence from Xaltocan, Mexico. Am. J. Phys. Anthropol. 2012, 149, 504–516. [Google Scholar] [CrossRef]
- Ochoa-Lugo, M.I.; Munoz, M.L.; Perez-Ramirez, G.; Beaty, K.G.; Lopez-Armenta, M.; Cervini-Silva, J.; Moreno-Galeana, M.; Meza, A.M.; Ramos, E.; Crawford, M.H.; et al. Genetic affiliation of pre-Hispanic and contemporary Mayas through maternal linage. Hum. Biol. 2016, 88, 136–167. [Google Scholar] [CrossRef] [PubMed]
- Monroe, C.; Kemp, B.M.; Smith, D.G. Exploring prehistory in the North American Southwest with mitochondrial DNA diversity exhibited by Yumans and Athapaskans. Am. J. Phys. Anthropol. 2013, 150, 618–631. [Google Scholar] [CrossRef]
- Malhi, R.S.; Mortensen, H.M.; Eshleman, J.A.; Kemp, B.M.; Lorenz, J.G.; Kaestle, F.A.; Johnson, J.R.; Gorodezky, C.; Smith, D.G. Native american mtDNA prehistory in the American Southwest. Am. J. Phys. Anthropol. 2003, 120, 108–124. [Google Scholar] [CrossRef]
- Rubi-Castellanos, R.; Martinez-Cortes, G.; Munoz-Valle, J.F.; Gonzalez-Martin, A.; Cerda-Flores, R.M.; Anaya-Palafox, M.; Rangel-Villalobos, H. Pre-Hispanic Mesoamerican demography approximates the present-day ancestry of Mestizos throughout the territory of Mexico. Am. J. Phys. Anthropol. 2009, 139, 284–294. [Google Scholar] [CrossRef]
- Bobillo, M.C.; Zimmermann, B.; Sala, A.; Huber, G.; Rock, A.; Bandelt, H.J.; Corach, D.; Parson, W. Amerindian mitochondrial DNA haplogroups predominate in the population of Argentina: Towards a first nationwide forensic mitochondrial DNA sequence database. Int. J. Leg. Med. 2010, 124, 263–268. [Google Scholar] [CrossRef]
- Gusmao, L.; Butler, J.M.; Linacre, A.; Parson, W.; Roewer, L.; Schneider, P.M.; Carracedo, A. Revised guidelines for the publication of genetic population data. Forensic Sci. Int. Genet. 2017, 30, 160–163. [Google Scholar] [CrossRef]
- Brenner, C.H. Fundamental problem of forensic mathematics-the evidential value of a rare haplotype. Forensic Sci. Int. Genet. 2010, 4, 281–291. [Google Scholar] [CrossRef]
- Irwin, J.A.; Ikramov, A.; Saunier, J.; Bodner, M.; Amory, S.; Rock, A.; O’Callaghan, J.; Nuritdinov, A.; Atakhodjaev, S.; Mukhamedov, R.; et al. The mtDNA composition of Uzbekistan: A microcosm of Central Asian patterns. Int. J. Leg. Med. 2010, 124, 195–204. [Google Scholar] [CrossRef]
- Rangel-Villalobos, H.; Munoz-Valle, J.F.; Gonzalez-Martin, A.; Gorostiza, A.; Magana, M.T.; Paez-Riberos, L.A. Genetic admixture, relatedness, and structure patterns among Mexican populations revealed by the Y-chromosome. Am. J. Phys. Anthropol. 2008, 135, 448–461. [Google Scholar] [CrossRef]
- Salazar-Flores, J.; Zuniga-Chiquette, F.; Rubi-Castellanos, R.; Alvarez-Miranda, J.L.; Zetina-Hernandez, A.; Martinez-Sevilla, V.M.; Gonzalez-Andrade, F.; Corach, D.; Vullo, C.; Alvarez, J.C.; et al. Admixture and genetic relationships of Mexican Mestizos regarding Latin American and Caribbean populations based on 13 CODIS-STRs. Homo 2015, 66, 44–59. [Google Scholar] [CrossRef] [PubMed]
- Cerda-Flores, R.M.; Budowle, B.; Jin, L.; Barton, S.A.; Deka, R.; Chakraborty, R. Maximum likelihood estimates of admixture in Northeastern Mexico using 13 short tandem repeat loci. Am. J. Hum. Biol. 2002, 14, 429–439. [Google Scholar] [CrossRef] [PubMed]
- Bryc, K.; Velez, C.; Karafet, T.; Moreno-Estrada, A.; Reynolds, A.; Auton, A.; Hammer, M.; Bustamante, C.D.; Ostrer, H. Genome-wide patterns of population structure and admixture among Hispanic/Latino populations. Proc. Natl. Acad. Sci. USA 2010, 107, 8954–8961. [Google Scholar] [CrossRef] [PubMed]
- Silva-Zolezzi, I.; Hidalgo-Miranda, A.; Estrada-Gil, J.; Fernandez-Lopez, J.C.; Uribe-Figueroa, L.; Contreras, A.; Balam-Ortiz, E.; del Bosque-Plata, L.; Velazquez-Fernandez, D.; Lara, C.; et al. Analysis of genomic diversity in Mexican Mestizo populations to develop genomic medicine in Mexico. Proc. Natl. Acad. Sci. USA 2009, 106, 8611–8616. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Estrada, A.; Gignoux, C.R.; Fernandez-Lopez, J.C.; Zakharia, F.; Sikora, M.; Contreras, A.V.; Acuna-Alonzo, V.; Sandoval, K.; Eng, C.; Romero-Hidalgo, S.; et al. Human genetics. The genetics of Mexico recapitulates Native American substructure and affects biomedical traits. Science 2014, 344, 1280–1285. [Google Scholar] [CrossRef] [PubMed]
- Huerta-Chagoya, A.; Moreno-Macias, H.; Fernandez-Lopez, J.C.; Ordonez-Sanchez, M.L.; Rodriguez-Guillen, R.; Contreras, A.; Hidalgo-Miranda, A.; Alfaro-Ruiz, L.A.; Salazar-Fernandez, E.P.; Moreno-Estrada, A.; et al. A panel of 32 AIMs suitable for population stratification correction and global ancestry estimation in Mexican Mestizos. BMC Genet. 2019, 20, 5. [Google Scholar] [CrossRef] [PubMed]
- Lopopolo, M.; Borsting, C.; Pereira, V.; Morling, N. A study of the peopling of Greenland using next generation sequencing of complete mitochondrial genomes. Am. J. Phys. Anthropol. 2016, 161, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Belbin, G.M.; Nieves-Colon, M.A.; Kenny, E.E.; Moreno-Estrada, A.; Gignoux, C.R. Genetic diversity in populations across Latin America: Implications for population and medical genetic studies. Curr. Opin. Genet. Dev. 2018, 53, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Colistro, V.; Rojas-Martinez, A.; Carracedo, A.; CHIBCA Consortium; Tomlinson, I.; Carvajal-Carmona, L.; Cruz, R.; Sans, M. CHIBCA Consortium; Population structure and relatedness estimates in a Mexican sample. Ann. Hum. Genet. 2021. [Google Scholar] [CrossRef] [PubMed]
- D’Amato, M.E.; Bodner, M.; Butler, J.M.; Gusmao, L.; Linacre, A.; Parson, W.; Schneider, P.M.; Vallone, P.; Carracedo, A. Ethical publication of research on genetics and genomics of biological material: Guidelines and recommendations. Forensic Sci. Int. Genet. 2020, 48, 102299. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Mexico | North | NW | NE | Center | CW | CE | South | SW | SE | West | East | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | 2021 | 567 | 388 | 179 | 622 | 360 | 262 | 709 | 629 | 80 | 1377 | 521 |
| A2 | 41.8 | 22.2 | 14.7 | 38.5 | 43.6 | 42.5 | 45.0 | 57.0 | 55.5 | 68.8 | 40.6 | 46.4 |
| B2 | 17.6 | 18.5 | 20.4 | 14.5 | 18.3 | 17.5 | 19.5 | 16.8 | 17.3 | 12.5 | 18.2 | 16.7 |
| C1 | 23.7 | 38.8 | 49.5 | 15.6 | 16.1 | 14.2 | 18.7 | 17.8 | 18.4 | 12.5 | 26.1 | 16.7 |
| D1 | 5.5 | 3.7 | 3.1 | 5.0 | 7.2 | 6.9 | 7.6 | 5.4 | 5.9 | 1.3 | 5.4 | 5.8 |
| D4h3(a) | 0.6 | 1.6 | 1.8 | 1.1 | 0.2 | -- | 0.4 | 0.1 | 0.2 | -- | 0.6 | 0.6 |
| X2(a) | 0.1 | 0.0 | 0.0 | -- | -- | -- | -- | -- | -- | -- | 0.0 | -- |
| West Eurasian | 8.0 | 12.0 | 7.2 | 22.3 | 10.6 | 13.6 | 6.5 | 1.6 | 1.6 | 1.3 | 6.3 | 11.1 |
| African | 2.1 | 2.1 | 1.8 | 2.8 | 3.7 | 5.0 | 1.9 | 1.0 | 1.1 | -- | 2.3 | 1.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bodner, M.; Perego, U.A.; Gomez, J.E.; Cerda-Flores, R.M.; Rambaldi Migliore, N.; Woodward, S.R.; Parson, W.; Achilli, A. The Mitochondrial DNA Landscape of Modern Mexico. Genes 2021, 12, 1453. https://doi.org/10.3390/genes12091453
Bodner M, Perego UA, Gomez JE, Cerda-Flores RM, Rambaldi Migliore N, Woodward SR, Parson W, Achilli A. The Mitochondrial DNA Landscape of Modern Mexico. Genes. 2021; 12(9):1453. https://doi.org/10.3390/genes12091453
Chicago/Turabian StyleBodner, Martin, Ugo A. Perego, J. Edgar Gomez, Ricardo M. Cerda-Flores, Nicola Rambaldi Migliore, Scott R. Woodward, Walther Parson, and Alessandro Achilli. 2021. "The Mitochondrial DNA Landscape of Modern Mexico" Genes 12, no. 9: 1453. https://doi.org/10.3390/genes12091453
APA StyleBodner, M., Perego, U. A., Gomez, J. E., Cerda-Flores, R. M., Rambaldi Migliore, N., Woodward, S. R., Parson, W., & Achilli, A. (2021). The Mitochondrial DNA Landscape of Modern Mexico. Genes, 12(9), 1453. https://doi.org/10.3390/genes12091453