
Sensogenomics and the Biological Background Underlying Musical Stimuli: Perspectives for a New Era of Musical Research




Abstract
:1. Introduction
2. Methodology
3. The Genetic Basis of Music in Human Evolution
3.1. The Origin of Musical Roots
3.2. Genetic Basis of Musical Abilities
3.3. Genomic Background Associated with Musical Traits
3.3.1. General Musical Abilities
3.3.2. Music Perception
3.3.3. Musical Memory
3.3.4. Musical Creativity
3.3.5. Musical Rhythm
3.3.6. Musical Language Production: Singing
3.4. Genetics and Music in Extreme Phenotypes
3.4.1. Genetic Research on Absolute Pitch (AP)
3.4.2. Biological Bases of Sound-Color Synesthesia
3.4.3. Williams Syndrome and Music
3.4.4. Autism Spectrum Disorder (ASD) and Music
3.4.5. Prader-Willi Syndrome, Genomic Imprinting, and Music
4. The Impact of Musical Stimuli in Our Genome
Genomics and Musical Stimuli: Impact of Music in Gene Expression
5. Sensogenomics and the Genetic Response to Musical Stimuli
6. Limitations of Present-Day Studies
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Zatorre, R.; McGill, J. Music, the food of neuroscience? Nature 2005, 434, 312–315. [Google Scholar] [CrossRef]
- Welch, G.; Ockelford, A. The importance of music in supporting the development of children with learning disabilities. Int. J. Birth Parent. Educ. 2015, 2, 21–23. [Google Scholar]
- McDermott, J. The evolution of music. Nature 2008, 453, 287–288. [Google Scholar] [CrossRef]
- Salimpoor, V.N.; Benovoy, M.; Longo, G.; Cooperstock, J.R.; Zatorre, R.J. The rewarding aspects of music listening are related to degree of emotional arousal. PLoS ONE 2009, 4, e7487. [Google Scholar] [CrossRef]
- Järvelä, I. Genomics studies on musical aptitude, music perception, and practice. Ann. N. Y. Acad. Sci. 2018, 1423, 82–91. [Google Scholar] [CrossRef]
- Blood, A.J.; Zatorre, R.J. Intensely pleasurable responses to music correlate with activity in brain regions implicated in reward and emotion. Proc. Natl. Acad. Sci. USA 2001, 98, 11818–11823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Särkämö, T.; Tervaniemi, M.; Laitinen, S.; Forsblom, A.; Soinila, S.; Mikkonen, M.; Autti, T.; Silvennoinen, H.M.; Erkkilä, J.; Laine, M.; et al. Music listening enhances cognitive recovery and mood after middle cerebral artery stroke. Brain 2008, 131, 866–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guétin, S.; Giniès, P.; Siou, D.K.; Picot, M.C.; Pommié, C.; Guldner, E.; Gosp, A.M.; Ostyn, K.; Coudeyre, E.; Touchon, J. The effects of music intervention in the management of chronic pain: A singleblind, randomized, controlled trial. Clin. J. Pain 2012, 28, 329–337. [Google Scholar] [CrossRef] [PubMed]
- Gingras, B.; Honing, H.; Peretz, I.; Trainor, L.J.; Fisher, S.E. Defining the biological bases of individual differences in musicality. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2015, 370, 20140092. [Google Scholar] [CrossRef] [Green Version]
- Tan, Y.T.; McPherson, G.E.; Peretz, I.; Berkovic, S.F.; Wilson, S.J. The genetic basis of music ability. Front. Psychol. 2014, 5, 658. [Google Scholar] [CrossRef] [Green Version]
- Conard, N.J.; Malina, M.; Munzel, S.C. New flutes document the earliest musical tradition in southwestern Germany. Nature 2009, 460, 737–740. [Google Scholar] [CrossRef]
- Mehr, S.A.; Singh, M.; Knox, D.; Ketter, D.M.; Pickens-Jones, D.; Atwood, S.; Lucas, C.; Jacoby, N.; Egner, A.A.; Hopkins, E.J.; et al. Universality and diversity in human song. Science 2019, 366, eaax0868. [Google Scholar] [CrossRef] [Green Version]
- Merriam, A.P. The Anthropology of Music; Northwestern University Press: Evanston, IL, USA, 1964. [Google Scholar]
- Nettl, B. Native American music; Routledge: New York, NY, USA, 2017. [Google Scholar]
- Wallin, N.L.; Merker, B.; Brown, S. The Origins of Music; Bradford Books: Cambridge, MA, USA, 2001. [Google Scholar]
- Ukkola-Vuoti, L.; Kanduri, C.; Oikkonen, J.; Buck, G.; Blancher, C.; Raijas, P.; Karma, K.; Lahdesmaki, H.; Jarvela, I. Genome-wide copy number variation analysis in extended families and unrelated individuals characterized for musical aptitude and creativity in music. PLoS ONE 2013, 8, e56356. [Google Scholar] [CrossRef] [PubMed]
- Alcock, K.J.; Passingham, R.E.; Watkins, K.; Vargha-Khadem, F. Pitch and timing abilities in inherited speech and language impairment. Brain Lang. 2000, 75, 34–46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baharloo, S.; Johnston, P.A.; Service, S.K.; Gitschier, J.; Freimer, N.B. Absolute pitch: An approach for identification of genetic and nongenetic components. Am. J. Hum. Genet. 1998, 62, 224–231. [Google Scholar] [CrossRef] [Green Version]
- Drayna, D.; Manichaikul, A.; de Lange, M.; Snieder, H.; Spector, T. Genetic correlates of musical pitch recognition in humans. Science 2001, 291, 1969–1972. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Manzano, O.; Ullen, F. Same Genes, Different Brains: Neuroanatomical Differences Between Monozygotic Twins Discordant for Musical Training. Cereb. Cortex 2018, 28, 387–394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mosing, M.A.; Ullen, F. Genetic influences on musical specialization: A twin study on choice of instrument and music genre. Ann. N. Y. Acad. Sci. 2018, 1423, 427–434. [Google Scholar] [CrossRef]
- Oikkonen, J.; Järvelä, I. Genomics approaches to study musical aptitude. Bioessays 2014, 36, 1102–1108. [Google Scholar] [CrossRef] [PubMed]
- Oikkonen, J.; Onkamo, P.; Järvelä, I.; Kanduri, C. Convergent evidence for the molecular basis of musical traits. Sci. Rep. 2016, 6, 39707. [Google Scholar] [CrossRef]
- Szyfter, K.; Witt, M.P. How far musicality and perfect pitch are derived from genetic factors? J. Appl. Genet. 2020, 61, 407–414. [Google Scholar] [CrossRef]
- Beccacece, L.; Abondio, P.; Cilli, E.; Restani, D.; Luiselli, D. Human Genomics and the Biocultural Origin of Music. Int. J. Mol. Sci. 2021, 22, 5397. [Google Scholar] [CrossRef]
- Coon, H.; Carey, G. Genetic and environmental determinants of musical ability in twins. Behav. Genet. 1989, 19, 183–193. [Google Scholar] [CrossRef] [PubMed]
- Vinkhuyzen, A.A.; van der Sluis, S.; Posthuma, D.; Boomsma, D.I. The heritability of aptitude and exceptional talent across different domains in adolescents and young adults. Behav. Genet. 2009, 39, 380–392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theusch, E.; Basu, A.; Gitschier, J. Genome-wide study of families with absolute pitch reveals linkage to 8q24.21 and locus heterogeneity. Am. J. Hum. Genet. 2009, 85, 112–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pulli, K.; Karma, K.; Norio, R.; Sistonen, P.; Goring, H.H.; Jarvela, I. Genome-wide linkage scan for loci of musical aptitude in Finnish families: Evidence for a major locus at 4q22. J. Med. Genet. 2008, 45, 451–456. [Google Scholar] [CrossRef]
- Oikkonen, J.; Huang, Y.; Onkamo, P.; Ukkola-Vuoti, L.; Raijas, P.; Karma, K.; Vieland, V.J.; Jarvela, I. A genome-wide linkage and association study of musical aptitude identifies loci containing genes related to inner ear development and neurocognitive functions. Mol. Psychiatry 2015, 20, 275–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, H.; Lee, S.; Kim, H.J.; Ju, Y.S.; Shin, J.Y.; Hong, D.; von Grotthuss, M.; Lee, D.S.; Park, C.; Kim, J.H.; et al. Comprehensive genomic analyses associate UGT8 variants with musical ability in a Mongolian population. J. Med. Genet. 2012, 49, 747–752. [Google Scholar] [CrossRef] [Green Version]
- Ukkola-Vuoti, L.; Oikkonen, J.; Onkamo, P.; Karma, K.; Raijas, P.; Jarvela, I. Association of the arginine vasopressin receptor 1A (AVPR1A) haplotypes with listening to music. J. Hum. Genet. 2011, 56, 324–329. [Google Scholar] [CrossRef] [PubMed]
- Ukkola, L.T.; Onkamo, P.; Raijas, P.; Karma, K.; Jarvela, I. Musical aptitude is associated with AVPR1A-haplotypes. PLoS ONE 2009, 4, e5534. [Google Scholar] [CrossRef] [Green Version]
- Granot, R.Y.; Uzefovsky, F.; Bogopolsky, H.; Ebstein, R.P. Effects of arginine vasopressin on musical working memory. Front. Psychol. 2013, 4, 712. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granot, R.Y.; Frankel, Y.; Gritsenko, V.; Lerer, E.; Gritsenko, I.; Bachner-Melman, R.; Israel, S.; Ebstein, R. Provisional evidence that the arginine vasopressin 1a receptor gene is associated with musical memory. Evolut. Hum. Behav. 2007, 28, 313–318. [Google Scholar] [CrossRef]
- Morley, A.P.; Narayanan, M.; Mines, R.; Molokhia, A.; Baxter, S.; Craig, G.; Lewis, C.M.; Craig, I. AVPR1A and SLC6A4 polymorphisms in choral singers and non-musicians: A gene association study. PLoS ONE 2012, 7, e31763. [Google Scholar] [CrossRef] [Green Version]
- Seesjarvi, E.; Sarkamo, T.; Vuoksimaa, E.; Tervaniemi, M.; Peretz, I.; Kaprio, J. The Nature and Nurture of Melody: A Twin Study of Musical Pitch and Rhythm Perception. Behav. Genet. 2016, 46, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, J.H.; Stelzer, J.; Fritz, T.H.; Chetelat, G.; La Joie, R.; Turner, R. Why musical memory can be preserved in advanced Alzheimer’s disease. Brain J. Neurol. 2015, 138, 2438–2450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Israel, S.; Lerer, E.; Shalev, I.; Uzefovsky, F.; Reibold, M.; Bachner-Melman, R.; Granot, R.; Bornstein, G.; Knafo, A.; Yirmiya, N.; et al. Molecular genetic studies of the arginine vasopressin 1a receptor (AVPR1a) and the oxytocin receptor (OXTR) in human behaviour: From autism to altruism with some notes in between. Prog. Brain Res. 2008, 170, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Oikkonen, J.; Kuusi, T.; Peltonen, P.; Raijas, P.; Ukkola-Vuoti, L.; Karma, K.; Onkamo, P.; Jarvela, I. Creative activities in music--A genome-wide linkage analysis. PLoS ONE 2016, 11, e0148679. [Google Scholar] [CrossRef] [PubMed]
- Donaldson, Z.R.; Young, L.J. Oxytocin, vasopressin, and the neurogenetics of sociality. Science 2008, 322, 900–904. [Google Scholar] [CrossRef] [Green Version]
- Niarchou, M.; Sathirapongsasuti, J.F.; Jacoby, N.; Bell, E.; McArthur, E.; Straub, P.; Team, T.A.R.; McAuley, J.D.; Capra, J.A.; Ullén, F.; et al. Unravelling the genetic architecture of musical rhythm. bioRxiv 2019. [Google Scholar]
- Lai, C.S.; Fisher, S.E.; Hurst, J.A.; Vargha-Khadem, F.; Monaco, A.P. A forkhead-domain gene is mutated in a severe speech and language disorder. Nature 2001, 413, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Campbell, P.; Reep, R.L.; Stoll, M.L.; Ophir, A.G.; Phelps, S.M. Conservation and diversity of Foxp2 expression in muroid rodents: Functional implications. J. Comp. Neurol. 2009, 512, 84–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atkinson, E.G.; Audesse, A.J.; Palacios, J.A.; Bobo, D.M.; Webb, A.E.; Ramachandran, S.; Henn, B.M. No Evidence for Recent Selection at FOXP2 among Diverse Human Populations. Cell 2018, 174, 1424–1435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, S.E. Human Genetics: The Evolving Story of FOXP2. Curr. Biol. 2019, 29, R65–R67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furlong, R. FOXP2 Tells a Cautionary Tale. Nat. Rev. Genet. 2018, 19, 592–593. [Google Scholar] [CrossRef] [PubMed]
- Miyazaki, K. Musical pitch identification by absolute pitch possessors. Percept. Psychophys. 1988, 44, 501–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregersen, P.K. Instant recognition: The genetics of pitch perception. Am. J. Hum. Genet. 1998, 62, 221–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Profita, J.; Bidder, T.G. Perfect pitch. Am. J. Med. Genet. 1988, 29, 763–771. [Google Scholar] [CrossRef]
- Takeuchi, A.H.; Hulse, S.H. Absolute pitch. Psychol. Bull. 1993, 113, 345–361. [Google Scholar] [CrossRef] [PubMed]
- Bachem, A. Absolute pitch. J. Acoust. Soc. Am. 1955, 27, 1180–1185. [Google Scholar] [CrossRef]
- Wellek, A. Musikpsychologie und Musikasthetik: Grundriss der systematischen Musikwissenschaft; Akademischer: Frankfurt, Germany, 1963. [Google Scholar]
- Revesz, G. Introduction to the Psychology of Music; Longmans Green: London, UK, 1953. [Google Scholar]
- Gregersen, P.K.; Kowalsky, E.; Kohn, N.; Marvin, E.W. Absolute pitch: Prevalence, ethnic variation, and estimation of the genetic component. Am. J. Hum. Genet. 1999, 65, 911–913. [Google Scholar] [CrossRef] [Green Version]
- Baharloo, S.; Service, S.K.; Risch, N.; Gitschier, J.; Freimer, N.B. Familial aggregation of absolute pitch. Am. J. Hum. Genet. 2000, 67, 755–758. [Google Scholar] [CrossRef] [Green Version]
- Gregersen, P.K.; Kowalsky, E.; Kohn, N.; Marvin, E.W. Early childhood music education and predisposition to absolute pitch: Teasing apart genes and environment. Am. J. Med. Genet. 2001, 98, 280–282. [Google Scholar] [CrossRef]
- Sacks, O. Musicophilia: Tales of Music and the Brain; Alfred A. Knopf: New York, NY, USA, 2007. [Google Scholar]
- Yu, L.; Fan, Y.; Deng, Z.; Huang, D.; Wang, S.; Zhang, Y. Pitch Processing in Tonal-Language-Speaking Children with Autism: An Event-Related Potential Study. J. Autism Dev. Disord. 2015, 45, 3656–3667. [Google Scholar] [CrossRef] [PubMed]
- Heaton, P. Pitch memory, labelling and disembedding in autism. J. Child Psychol. Psychiatry 2003, 44, 543–551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heaton, P. Assessing musical skills in autistic children who are not savants. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2009, 364, 1443–1447. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gregersen, P.K.; Kowalsky, E.; Lee, A.; Baron-Cohen, S.; Fisher, S.E.; Asher, J.E.; Ballard, D.; Freudenberg, J.; Li, W. Absolute pitch exhibits phenotypic and genetic overlap with synesthesia. Hum. Mol. Genet. 2013, 22, 2097–2104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilot, A.K.; Kucera, K.S.; Vino, A.; Asher, J.E.; Baron-Cohen, S.; Fisher, S.E. Rare variants in axonogenesis genes connect three families with sound-color synesthesia. Proc. Natl. Acad. Sci. USA 2018, 115, 3168–3173. [Google Scholar] [CrossRef] [Green Version]
- Baron-Cohen, S.; Burt, L.; Smith-Laittan, F.; Harrison, J.; Bolton, P. Synaesthesia: Prevalence and familiality. Perception 1996, 25, 1073–1079. [Google Scholar] [CrossRef]
- Melero, H. Sinestesia, Bases Neuroanatómicas y Cognitivas. Universiry Complutense of Madrid: Madrid, Spain, Unpublished work. 2015. [Google Scholar]
- Asher, J.E.; Lamb, J.A.; Brocklebank, D.; Cazier, J.B.; Maestrini, E.; Addis, L.; Sen, M.; Baron-Cohen, S.; Monaco, A.P. A whole-genome scan and fine-mapping linkage study of auditory-visual synesthesia reveals evidence of linkage to chromosomes 2q24, 5q33, 6p12, and 12p12. Am. J. Hum. Genet. 2009, 84, 279–285. [Google Scholar] [CrossRef] [Green Version]
- Lashkari, A.; Smith, A.K.; Graham, J.M., Jr. Williams-Beuren syndrome: An update and review for the primary physician. Clin. Pediatr. 1999, 38, 189–208. [Google Scholar] [CrossRef]
- Levitin, D.J. Musical behavior in a neurogenetic developmental disorder: Evidence from Williams Syndrome. Ann. N. Y. Acad. Sci. 2005, 1060, 325–334. [Google Scholar] [CrossRef]
- Levitin, D.J.; Menon, V.; Schmitt, J.E.; Eliez, S.; White, C.D.; Glover, G.H.; Kadis, J.; Korenberg, J.R.; Bellugi, U.; Reiss, A.L. Neural correlates of auditory perception in Williams syndrome: An fMRI study. Neuroimage 2003, 18, 74–82. [Google Scholar] [CrossRef] [Green Version]
- Wengenroth, M.; Blatow, M.; Bendszus, M.; Schneider, P. Leftward lateralization of auditory cortex underlies holistic sound perception in Williams syndrome. PLoS ONE 2010, 5, e12326. [Google Scholar] [CrossRef]
- Canales, C.P.; Wong, A.C.; Gunning, P.W.; Housley, G.D.; Hardeman, E.C.; Palmer, S.J. The role of GTF2IRD1 in the auditory pathology of Williams-Beuren Syndrome. Eur. J. Hum. Genet. 2015, 23, 774–780. [Google Scholar] [CrossRef] [Green Version]
- Mulle, J.G.; Pulver, A.E.; McGrath, J.A.; Wolyniec, P.S.; Dodd, A.F.; Cutler, D.J.; Sebat, J.; Malhotra, D.; Nestadt, G.; Conrad, D.F.; et al. Reciprocal duplication of the Williams-Beuren syndrome deletion on chromosome 7q11.23 is associated with schizophrenia. Biol. Psychiatry 2014, 75, 371–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, L.; Carter, C.S.; Ying, J.; Bellugi, U.; Pournajafi-Nazarloo, H.; Korenberg, J.R. Oxytocin and vasopressin are dysregulated in Williams Syndrome, a genetic disorder affecting social behavior. PLoS ONE 2012, 7, e38513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Remington, A.; Fairnie, J. A sound advantage: Increased auditory capacity in autism. Cognition 2017, 166, 459–465. [Google Scholar] [CrossRef] [PubMed]
- Molnar-Szakacs, I.; Heaton, P. Music: A unique window into the world of autism. Ann. N. Y. Acad. Sci. 2012, 1252, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Hammock, E.A.; Young, L.J. Oxytocin, vasopressin and pair bonding: Implications for autism. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 2006, 361, 2187–2198. [Google Scholar] [CrossRef] [Green Version]
- Yirmiya, N.; Rosenberg, C.; Levi, S.; Salomon, S.; Shulman, C.; Nemanov, L.; Dina, C.; Ebstein, R.P. Association between the arginine vasopressin 1a receptor (AVPR1a) gene and autism in a family-based study: Mediation by socialization skills. Mol. Psychiatry 2006, 11, 488–494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jacob, S.; Brune, C.W.; Carter, C.S.; Leventhal, B.L.; Lord, C.; Cook, E.H., Jr. Association of the oxytocin receptor gene (OXTR) in Caucasian children and adolescents with autism. Neurosci. Lett. 2007, 417, 6–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehr, S.A.; Kotler, J.; Howard, R.M.; Haig, D.; Krasnow, M.M. Genomic imprinting is implicated in the psychology of music. Psychol. Sci. 2017, 28, 1455–1467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallam, S. The Power of Music: A Research Synthesis of the Impact of Actively Making Music on the Intellectual, Social and Personal Development of Children and Young People; Institute of Education University College: London, UK, 2015. [Google Scholar]
- Bittman, B.; Berk, L.; Shannon, M.; Sharaf, M.; Westengard, J.; Guegler, K.J.; Ruff, D.W. Recreational music-making modulates the human stress response: A preliminary individualized gene expression strategy. Med. Sci. Monit. 2005, 11, BR31–BR40. [Google Scholar] [PubMed]
- Qu, S.; Olafsrud, S.M.; Meza-Zepeda, L.A.; Saatcioglu, F. Rapid gene expression changes in peripheral blood lymphocytes upon practice of a comprehensive yoga program. PLoS ONE 2013, 8, e61910. [Google Scholar] [CrossRef] [Green Version]
- Kanduri, C.; Raijas, P.; Ahvenainen, M.; Philips, A.K.; Ukkola-Vuoti, L.; Lahdesmaki, H.; Jarvela, I. The effect of listening to music on human transcriptome. PeerJ 2015, 3, e830. [Google Scholar] [CrossRef] [Green Version]
- Nair, P.S.; Raijas, P.; Ahvenainen, M.; Philips, A.K.; Ukkola-Vuoti, L.; Jarvela, I. Music-listening regulates human microRNA expression. Epigenetics 2020, 16, 554–566. [Google Scholar] [CrossRef]
- Kanduri, C.; Kuusi, T.; Ahvenainen, M.; Philips, A.K.; Lahdesmaki, H.; Jarvela, I. The effect of music performance on the transcriptome of professional musicians. Sci. Rep. 2015, 5, 9506. [Google Scholar] [CrossRef] [Green Version]
- Nair, P.S.; Kuusi, T.; Ahvenainen, M.; Philips, A.K.; Jarvela, I. Music-performance regulates microRNAs in professional musicians. PeerJ 2019, 7, e6660. [Google Scholar] [CrossRef] [Green Version]
- Ramirez-Rivera, S.; Bernal, G. Music Is Capable of Inducing Changes in Gene Expression in Gastric Cancer Cells. J. Gastrointest. Cancer 2019, 50, 175–180. [Google Scholar] [CrossRef]
- Neisser, U. Cognitive Psychology; Psychology Press: New York, NY, USA, 1967. [Google Scholar]
- Peretz, I.; Zatorre, R.J. Brain organization for music processing. Annu. Rev. Psychol. 2005, 56, 89–114. [Google Scholar] [CrossRef] [Green Version]
- Honing, H. On the biological basis of musicality. Ann. N. Y. Acad. Sci. 2018, 1423, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Munafo, M.R.; Clark, T.G.; Flint, J. Assessing publication bias in genetic association studies: Evidence from a recent meta-analysis. Psychiatry Res. 2004, 129, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Hewitt, J.K. Editorial policy on candidate gene association and candidate gene-by-environment interaction studies of complex traits. Behav. Genet. 2012, 42, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Jacoby, N.; Margulis, E.H.; Clayton, M.; Hannon, E.; Honing, H.; Iversen, J.; Klein, T.R.; Mehr, S.A.; Pearson, L.; Peretz, I.; et al. Cross-cultural work in music cognition: Challenges, insights, and recommendations. Music Percept. 2020, 37, 185–195. [Google Scholar] [CrossRef] [Green Version]
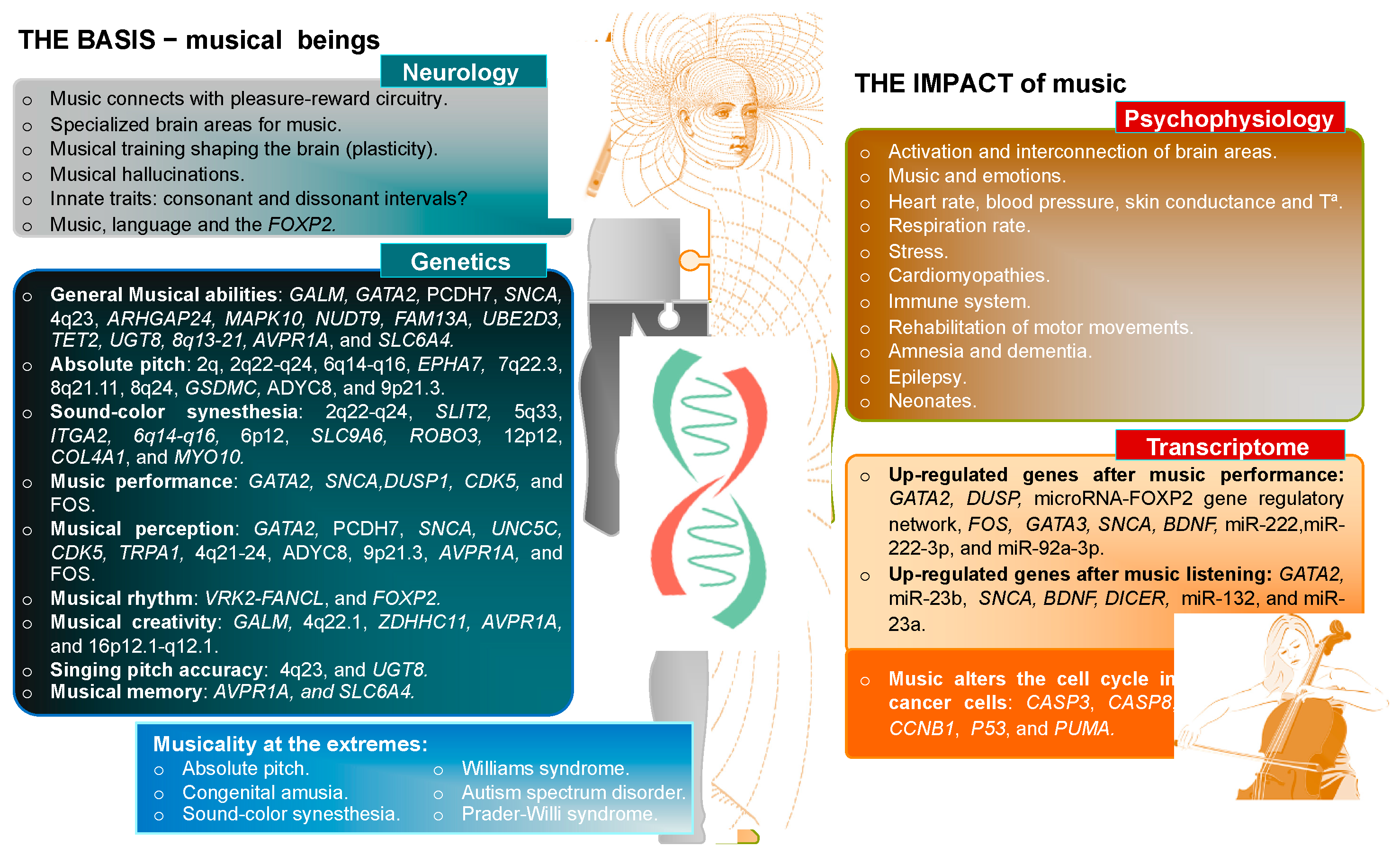

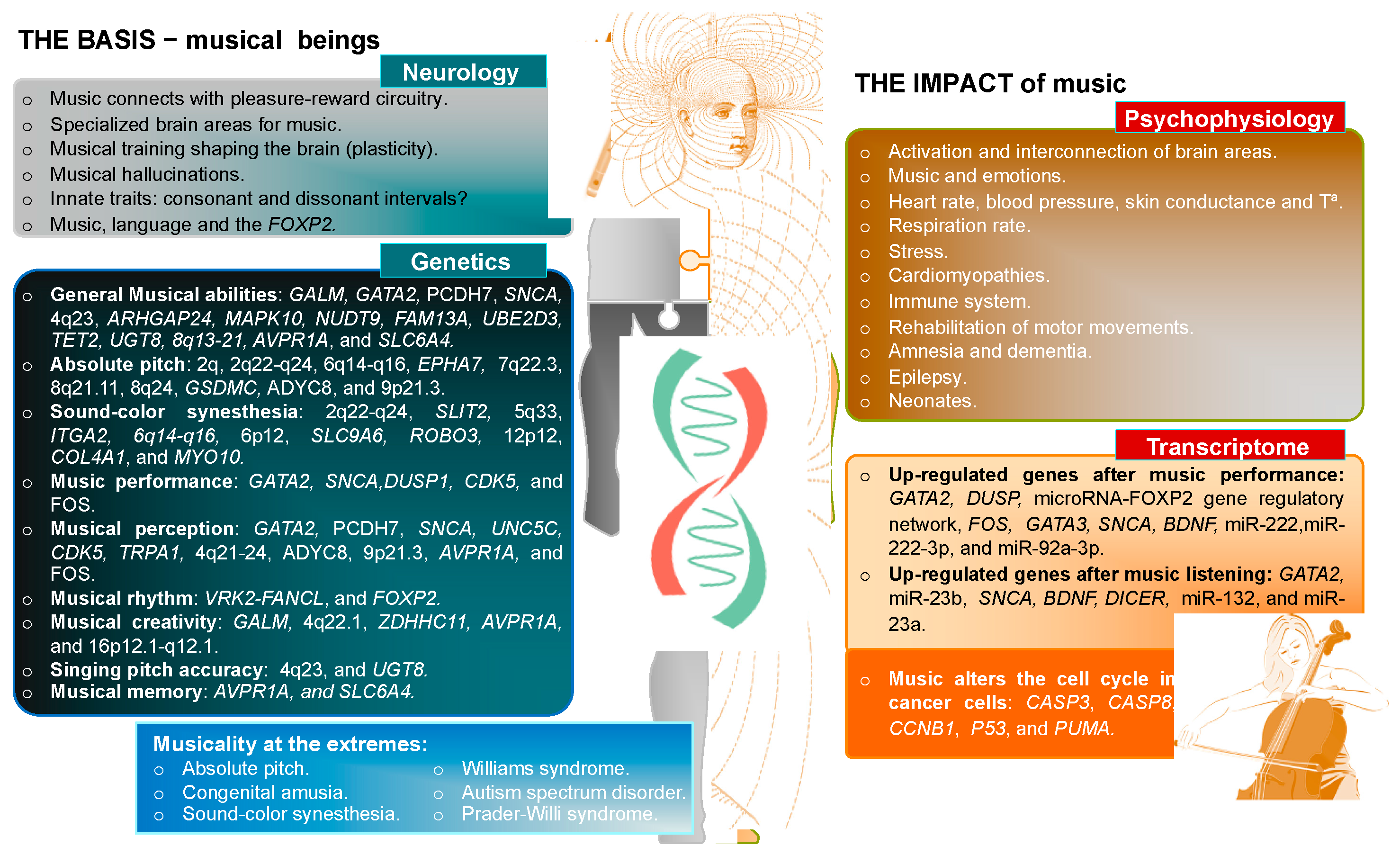{kind=link}
| STUDIES | FOCUS | TYPE OF STUDY | CASES/CONTROLS | |
|---|---|---|---|---|
| DNA: MUSICAL TRAITS | Coon and Carey (1989) | Musical abilities | Twin study | n = 98 MZ and 70 DZ males + 93 MZ and 132 DZ females |
| Vinkhuyzen et al., (2009) | Musical abilities | Twin study | n = 1685 twin pairs | |
| Drayna et al., (2001) | Music perception | Twin study | n = 136 MZ + 148 DZ | |
| Pulli et al., (2008) | Music perception | GWLS/pedigree study | n = 234 from 15 families | |
| Seeajärvi et al., (2016) | Music perception | Twin study | n = 384 (69MZ, 44DZ + 70MZ, and 88DZ, without a co-twin) | |
| Ukkola-Vuoti et al., (2009) | Music perception/creativity | Candidate genes/pedigree study | n = 343 from 19 families | |
| Ukkola-Vuoti et al., (2011) | Music perception | Candidate genes | n = 437 from 31 families | |
| Ukkola-Vuoti et al., (2013) | Music perception/creativity | Genome-wide CNVs | n = 170 cases from 5 families + 172 controls | |
| Oikkonen et al., (2015) | Music perception | GWLS/GWAS | n = 767 from 76 families | |
| Oikkonen et al., (2016a) | Musical creativity | GWLS | n = 474 cases from 79 families + 103 controls | |
| Granot et al., (2007) | Musical memory | Candidate genes | n = 82 cases | |
| Park et al., (2012) | Musical language production | GWLS/GWAS/exome sequencing/aCGH | n = 1008 from 73 families | |
| Morley et al., (2012) | Musical language production | Candidate genes | n = 262 cases + 261 controls | |
| Niarchou et al., (2019) | Rhythm | GWAS | n = 606,825 | |
| DNA AND EXTRE PHENOTYPES | Profita and Bidder (1988) | AP | Family segregation analysis | n = 35 AP from 19 families |
| Gregersen and Kumar (1996) | AP | Familiar aggregation | n = 101 AP | |
| Baharloo et al., (1998) | AP | Familiar aggregation | n = 612 (92 self-reported AP + 520 non-AP/controls) | |
| Gregersen et al., (1999) | AP | Familiar aggregation | n = 2707 music students/249 AP | |
| Baharloo et al., (2000) | AP | Familiar aggregation | n = 74 AP with 113 siblings + 625 controls | |
| Gregersen et al., (2001) | AP | Familiar aggregation | n = 1067 cases (Asian/no Asian) | |
| Theusch et al., (2009) | AP | GWLS | n = 281 from 73 AP families with at least 2 AP possessors | |
| Theusch and Gitschier (2011) | AP | Family segregation analysis/twin study | n = 1463 families + 14 MZ + 31 DZ | |
| Gregersen et al., (2013) | AP/synesthesia | GWLS/exome sequencing | n = 768 AP subjects/151 reported synesthesia | |
| Asher et al., (2009) | Sound-color synesthesia | Whole-genome scan/fine-mapping linkage study | n = 196 from 43 multiplex families | |
| Tilot et al., (2018) | Sound-color synesthesia | WES | n = 3 families | |
| Peretz et al., (2007) | Congenital amusia | Family aggregation study | n = 71 cases + 75 controls | |
| Kalmus and Fry (1980) | Congenital amusia | Pedigree study/twin–sibling study | n = 600 | |
| Lai et al., (2001) | Speech and language disorder | Gene expression analysis/DNA sequencing | n = 3 generation pedigree | |
| MUSIC AND GENE EXPRESSION | Bittman et al., (2005) | Music performance and stress | Gene expression analysis | n = 32 (16 cases + 16 control) |
| QU et al., (2013) | Music listening and stress | Gene expression analysis | n = 14 cases | |
| Kanduri et al., (2015a) | Music listening | Gene expression analysis | n = 48 cases/audience + 15 controls | |
| Kanduri et al., (2015b) | Music performance | Gene expression analysis | n = 10 professional musicians + music-free control session | |
| Nair et al., (2020) | Music listening | Sequencing microRNAs | n = 48 cases/audience + 15 controls | |
| Nair et al., (2019) | Music performance | Sequencing microRNAs | n = 10 professional musicians + music-free control session | |
| Ramirez-Rivera and Bernal (2019) | Musical impact | Gene expression analysis | n = 5.000 gastric cancer cells |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Navarro, L.; Martinón-Torres, F.; Salas, A. Sensogenomics and the Biological Background Underlying Musical Stimuli: Perspectives for a New Era of Musical Research. Genes 2021, 12, 1454. https://doi.org/10.3390/genes12091454
Navarro L, Martinón-Torres F, Salas A. Sensogenomics and the Biological Background Underlying Musical Stimuli: Perspectives for a New Era of Musical Research. Genes. 2021; 12(9):1454. https://doi.org/10.3390/genes12091454
Chicago/Turabian StyleNavarro, Laura, Federico Martinón-Torres, and Antonio Salas. 2021. "Sensogenomics and the Biological Background Underlying Musical Stimuli: Perspectives for a New Era of Musical Research" Genes 12, no. 9: 1454. https://doi.org/10.3390/genes12091454
APA StyleNavarro, L., Martinón-Torres, F., & Salas, A. (2021). Sensogenomics and the Biological Background Underlying Musical Stimuli: Perspectives for a New Era of Musical Research. Genes, 12(9), 1454. https://doi.org/10.3390/genes12091454