
Highly Conservative Pattern of Sex Chromosome Synapsis and Recombination in Neognathae Birds

 , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Specimens
2.2. SC Spreading and Immunostaining
2.3. Generation of the Microdissected DNA Probe and FISH
2.4. Microscopic Analysis
2.5. Chromosome Measurements and Generation of Recombination Maps of GRCs
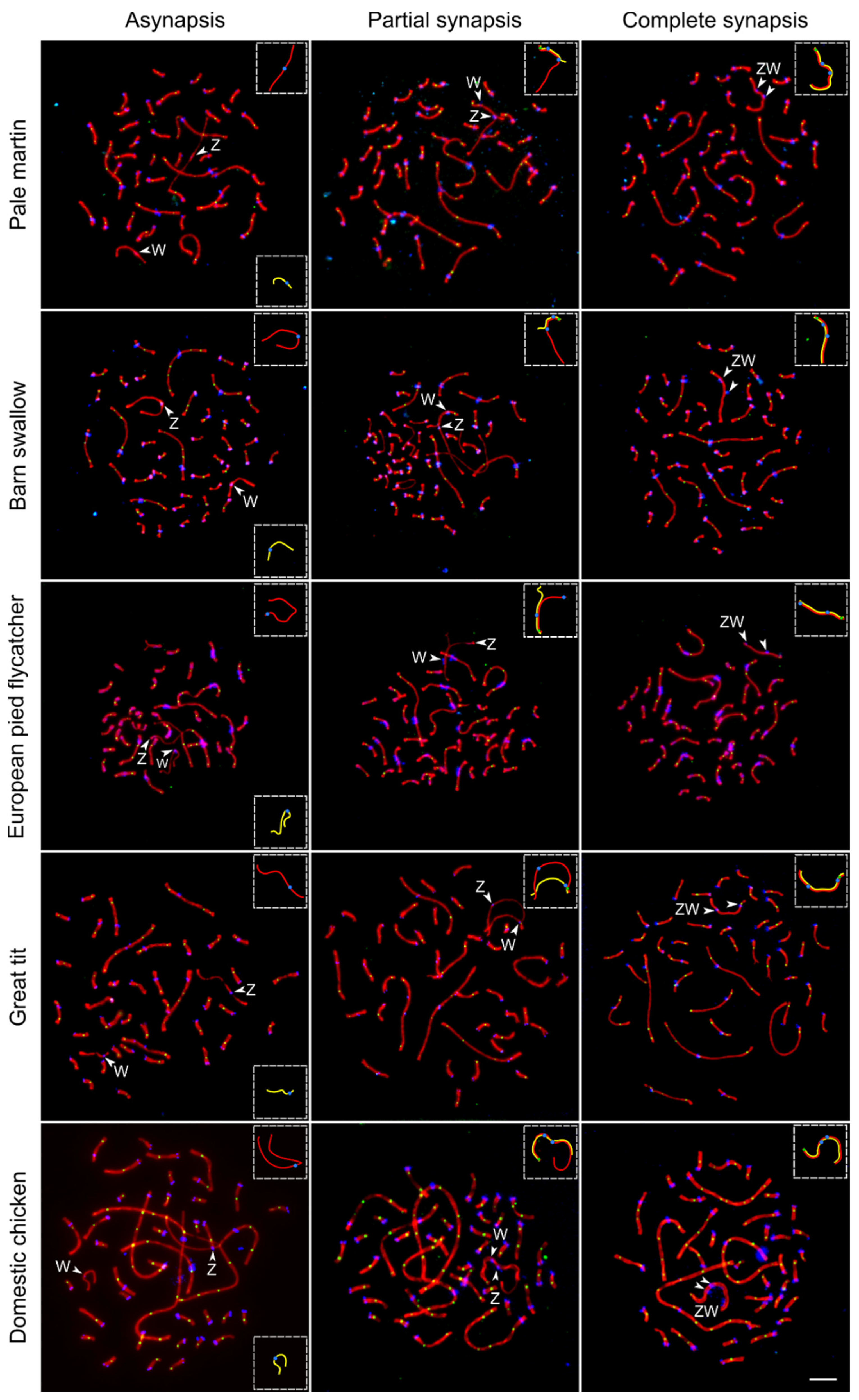
3. Results
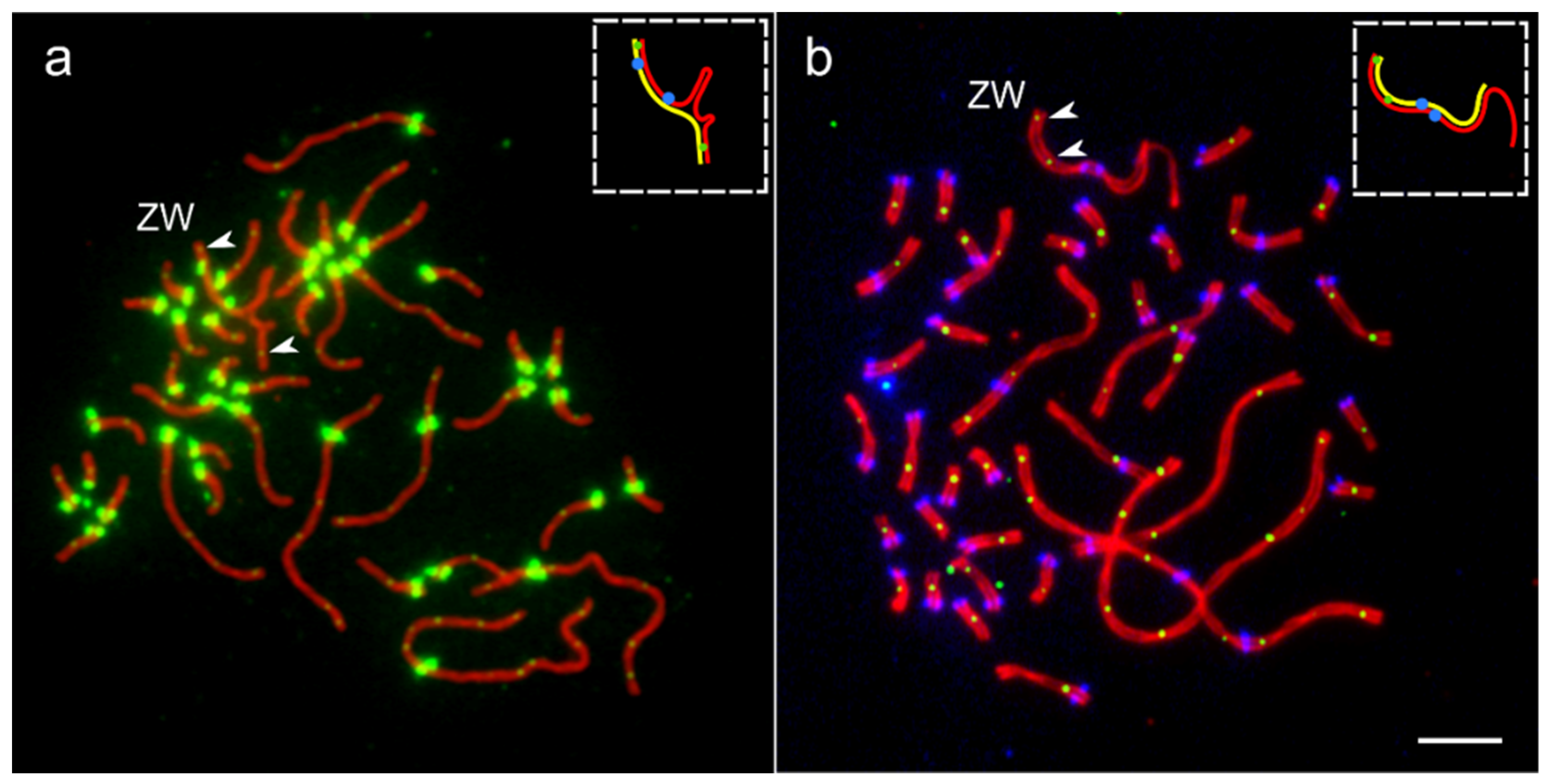
3.1. Common Features of ZW Synapsis
3.2. MLH1 Foci Distribution along ZW Bivalents
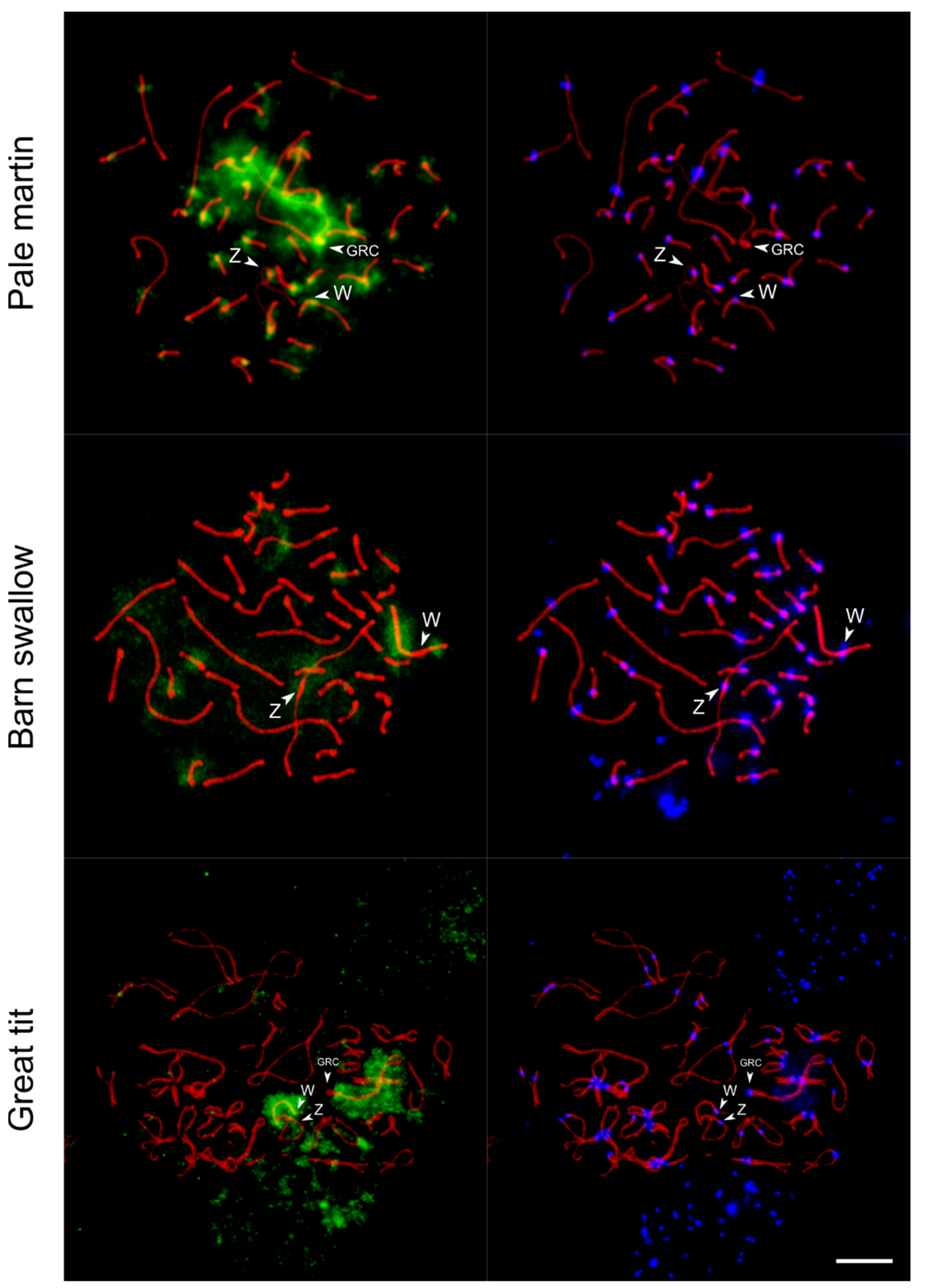
3.3. FISH of Whole-Chromosome Probe of the Pale Martin GRC with Pachytene Chromosomes of Three Songbird Species
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fridolfsson, A.K.; Cheng, H.; Copeland, N.G.; Jenkins, N.A.; Liu, H.C.; Raudsepp, T.; Woodage, T.; Chowdhary, B.; Halverson, J.; Ellegren, H. Evolution of the avian sex chromosomes from an ancestral pair of autosomes. Proc. Natl. Acad. Sci. USA 1998, 95, 8147–8152. [Google Scholar] [CrossRef]
- Pigozzi, M.I. The chromosomes of birds during meiosis. Cytogenet. Genome Res. 2016, 150, 128–138. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Zhang, J.; Bachtrog, D.; An, N.; Huang, Q.; Jarvis, E.D.; Gilbert, M.T.P.; Zhang, G. Complex evolutionary trajectories of sex chromosomes across bird taxa. Science 2014, 346, 1246338. [Google Scholar] [CrossRef]
- Pigozzi, M.I.; Solari, A.J. Extreme axial equalization and wide distribution of recombination nodules in the primitive ZW pair of Rhea americana (Aves, Ratitae). Chromosom. Res. 1997, 5, 421–428. [Google Scholar] [CrossRef] [PubMed]
- Pigozzi, M.I.; Solari, A.J. The ZW pairs of two paleognath birds from two orders show transitional stages of sex chromosome differentiation. Chromosom. Res. 1999, 7, 541–551. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.E.; Harrison, P.W.; Montgomery, S.H.; Pointer, M.A.; Mank, J.E. Independent stratum formation on the avian sex chromosomes reveals inter-chromosomal gene conversion and predominance of purifying selection on the w chromosome. Evolution 2014, 68, 3281–3295. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Wright, A.E.; Moghadam, H.K.; Mank, J.E. Trade-off between selection for dosage compensation and masculinization on the avian Z chromosome. Genetics 2012, 192, 1433–1445. [Google Scholar] [CrossRef]
- Xu, L.; Auer, G.; Peona, V.; Suh, A.; Deng, Y.; Feng, S.; Zhang, G.; Blom, M.P.K.; Christidis, L.; Prost, S.; et al. Dynamic evolutionary history and gene content of sex chromosomes across diverse songbirds. Nat. Ecol. Evol. 2019, 3, 834–844. [Google Scholar] [CrossRef] [PubMed]
- Pigozzi, M.I.; Solari, A.J. Recombination nodule mapping and chiasma distribution in spermatocytes of the pigeon, Columba livia. Genome 1999, 42, 308–314. [Google Scholar] [CrossRef]
- Solari, A.J. Equalization of Z and W axes in chicken and quail oocytes. Cytogenet. Genome Res. 1992, 59, 52–56. [Google Scholar] [CrossRef]
- Solari, A.J.; Pigozzi, M.I. Recombination nodules and axial equalization in the ZW pairs of the Peking duck and the guinea fowl. Cytogenet Cell Genet 1993, 64, 268–272. [Google Scholar] [CrossRef]
- Pigozzi, M.I.; del Priore, L. Meiotic recombination analysis in female ducks (Anas platyrhynchos). Genetica 2016, 144, 307–312. [Google Scholar] [CrossRef]
- Torgasheva, A.A.; Borodin, P.M. Immunocytological analysis of meiotic recombination in the gray goose (Anser anser). Cytogenet. Genome Res. 2017, 151, 27–35. [Google Scholar] [CrossRef]
- Semenov, G.A.; Basheva, E.A.; Borodin, P.M.; Torgasheva, A.A. High rate of meiotic recombination and its implications for intricate speciation patterns in the white wagtail (Motacilla alba). Biol. J. Linn. Soc. 2018, 125, 600–612. [Google Scholar] [CrossRef]
- Lisachov, A.P.; Malinovskaya, L.P.; Druzyaka, A.V.; Borodin, P.M.; Torgasheva, A.A. Synapsis and recombination of autosomes and sex chromosomes in two terns (Sternidae, Charadriiformes, Aves). Vavilov J. Genet. Breed. 2017, 21, 259–268. [Google Scholar] [CrossRef]
- Pigozzi, M.I. Distribution of MLH1 foci on the synaptonemal complexes of chicken oocytes. Cytogenet Cell Genet 2001, 95, 129–133. [Google Scholar] [CrossRef] [PubMed]
- del Priore, L.; Pigozzi, M.I. Sex-specific recombination maps for individual macrochromosomes in the Japanese quail (Coturnix japonica). Chromosom. Res. 2015, 23, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Torgasheva, A.A.; Malinovskaya, L.P.; Zadesenets, K.S.; Karamysheva, T.V.; Kizilova, E.A.; Akberdina, E.A.; Pristyazhnyuk, I.E.; Shnaider, E.P.; Volodkina, V.A.; Saifitdinova, A.F.; et al. Germline-restricted chromosome (GRC) is widespread among songbirds. Proc. Natl. Acad. Sci. USA 2019, 116, 11650–11845. [Google Scholar] [CrossRef] [PubMed]
- Pigozzi, M.I.; Solari, A.J. The germ-line-restricted chromosome in the zebra finch: Recombination in females and elimination in males. Chromosoma 2005, 114, 403–409. [Google Scholar] [CrossRef] [PubMed]
- Pigozzi, M.I.; Solari, A.J. Germ cell restriction and regular transmission of an accessory chromosome that mimics a sex body in the zebra finch, Taeniopygia guttata. Chromosom. Res. 1998, 6, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Malinovskaya, L.P.; Zadesenets, K.S.; Karamysheva, T.V.; Akberdina, E.A.; Kizilova, E.A.; Romanenko, M.V.; Shnaider, E.P.; Scherbakova, M.M.; Korobitsyn, I.G.; Rubtsov, N.B.; et al. Germline-restricted chromosome (GRC) in the sand martin and the pale martin (Hirundinidae, Aves): Synapsis, recombination and copy number variation. Sci. Rep. 2020, 10, 1058. [Google Scholar] [CrossRef] [PubMed]
- Pei, Y.; Forstmeier, W.; Ruiz-Ruano, F.J.; Mueller, J.C.; Cabrero, J.; Camacho, J.P.M.; Alché, J.D.; Franke, A.; Hoeppner, M.; Börno, S.; et al. Occasional paternal inheritance of the germline-restricted chromosome in songbirds. bioRxiv 2021. [Google Scholar] [CrossRef]
- Kinsella, C.M.; Ruiz-Ruano, F.J.; Dion-Côté, A.M.; Charles, A.J.; Gossmann, T.I.; Cabrero, J.; Kappei, D.; Hemmings, N.; Simons, M.J.P.; Camacho, J.P.M.; et al. Programmed DNA elimination of germline development genes in songbirds. Nat. Commun. 2019, 10, 5468. [Google Scholar] [CrossRef] [PubMed]
- Stöck, M.; Kratochvíl, L.; Kuhl, H.; Rovatsos, M.; Evans, B.J.; Suh, A.; Valenzuela, N.; Veyrunes, F.; Zhou, Q.; Gamble, T.; et al. A brief review of vertebrate sex evolution with a pledge for integrative research: Towards ‘sexomics’. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200426. [Google Scholar] [CrossRef] [PubMed]
- Peters, A.H.; Plug, A.W.; van Vugt, M.J.; de Boer, P. A drying-down technique for the spreading of mammalian meiocytes from the male and female germline. Chromosom. Res. 1997, 5, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Anderson, L.K.; Reeves, A.; Webb, L.M.; Ashley, T. Distribution of crossing over on mouse synaptonemal complexes using immunofluorescent localization of MLH1 protein. Genetics 1999, 151, 1569–1579. [Google Scholar] [CrossRef] [PubMed]
- Zadesenets, K.S.; Schärer, L.; Rubtsov, N.B. New insights into the karyotype evolution of the free-living flatworm Macrostomum lignano (Platyhelminthes, Turbellaria). Sci. Rep. 2017, 7, 6066. [Google Scholar] [CrossRef]
- Trifonov, V.; Vorobieva, N.; Serdyukova, N.; Rens, W. FISH with and without COT1 DNA. In Fluorescence In Situ Hybridization (FISH)—Application Guide.; Liehr, T., Ed.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 123–133. [Google Scholar]
- Reeves, A. MicroMeasure: A new computer program for the collection and analysis of cytogenetic data. Genome 2001, 44, 439–443. [Google Scholar] [CrossRef]
- Pigozzi, M.I.; Solari, A.J. Equal frequencies of recombination nodules in both sexes of the pigeon suggest a basic difference with eutherian mammals. Genome 1999, 42, 315–321. [Google Scholar] [CrossRef]
- Zickler, D.; Kleckner, N. A few of our favorite things: Pairing, the bouquet, crossover interference and evolution of meiosis. Semin. Cell Dev. Biol. 2016, 54, 135–148. [Google Scholar] [CrossRef]
- Solari, A.J. Synaptic behaviour and recombination nodules in the human XY pair. Genetica 1988, 77, 149–158. [Google Scholar] [CrossRef]
- Li, L.; Hamer, D.H. Recombination and allelic association in the Xq/Yq homology region. Hum. Mol. Genet. 1995, 4, 2013–2016. [Google Scholar] [CrossRef]
- Sun, F.; Oliver-Bonet, M.; Liehr, T.; Starke, H.; Ko, E.; Rademaker, A.; Navarro, J.; Benet, J.; Martin, R.H. Human male recombination maps for individual chromosomes. Am. J. Hum. Genet. 2004, 74, 521–531. [Google Scholar] [CrossRef]
- Pala, I.; Naurin, S.; Stervander, M.; Hasselquist, D.; Bensch, S.; Hansson, B. Evidence of a neo-sex chromosome in birds. Heredity 2012, 108, 264–272. [Google Scholar] [CrossRef]
- Sigeman, H.; Ponnikas, S.; Hansson, B. Whole-genome analysis across 10 songbird families within Sylvioidea reveals a novel autosome–sex chromosome fusion. Biol. Lett. 2020, 16, 20200082. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Furo, I.; Peona, V.; Liu, J.; Gomes, A.J.B.; Cen, W.; Huang, H.; Zhang, Y.; Chen, D.; Ting, X.; et al. Recurrent chromosome reshuffling and the evolution of neo-sex chromosomes in parrots. bioRxiv 2021. [Google Scholar] [CrossRef]
- Gan, H.M.; Falk, S.; Morales, H.E.; Austin, C.M.; Sunnucks, P.; Pavlova, A. Genomic evidence of neo-sex chromosomes in the eastern yellow robin. Gigascience 2019, 8, 1–10. [Google Scholar] [CrossRef]
- Sigeman, H.; Ponnikas, S.; Chauhan, P.; Dierickx, E.; Brooke, M.d.L.; Hansson, B. Repeated sex chromosome evolution in vertebrates supported by expanded avian sex chromosomes. Proc. R. Soc. B Biol. Sci. 2019, 286, 20192051. [Google Scholar] [CrossRef]
- Itoh, Y.; Kampf, K.; Pigozzi, M.I.; Arnold, A.P. Molecular cloning and characterization of the germline-restricted chromosome sequence in the zebra finch. Chromosoma 2009, 118, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Hansson, B. On the origin and evolution of germline chromosomes in songbirds. Proc. Natl. Acad. Sci. USA 2019, 116, 11570–11572. [Google Scholar] [CrossRef] [PubMed]
- Peona, V.; Palacios-Gimenez, O.M.; Blommaert, J.; Liu, J.; Haryoko, T.; Jønsson, K.A.; Irestedt, M.; Zhou, Q.; Jern, P.; Suh, A. The avian W chromosome is a refugium for endogenous retroviruses with likely effects on female-biased mutational load and genetic incompatibilities. Philos. Trans. R. Soc. B Biol. Sci. 2021, 376, 20200186. [Google Scholar] [CrossRef] [PubMed]
- Asalone, K.C. Analysis of the Mysterious Germline Restricted Chromosome in Zebra Finch; American University: Washington, DC, USA, 2021. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Domestic Chicken | Grey Goose | Black Tern | Common Tern | Pale Martin | Barn Swallow | European Pied Flycatcher | Great Tit | White Wagtail |
|---|---|---|---|---|---|---|---|---|---|
| Number of birds | 3 | 3 | 1 | 2 | 3 | 3 | 2 | 4 | 2 |
| Pairing arms | Zp-Wp | Zq-Wp | Zp-Wp | Zp-Wp | Zp-Wp | Zp-Wp | Zq-Wp | Zq-Wp | Zp-Wp and Zq-Wq |
| Complete synapsis | |||||||||
| N * | 57 | 50 | 89 | 150 | 61 | 128 | 80 | 152 | 57 |
| Absolute length of ZW, µm | 10.7 ± 3.3 | 14.6 ± 4.0 | 15.6 ± 3.3 | 12.5 ± 2.1 | 9.1 ± 2.5 | 11.3 ± 1.8 | 12.3 ± 3.4 | 9.1 ± 1.6 | 10.3 ± 1.7 |
| Relative length of ZW **, % | 4.7 | 4.9 | 5.1 | 5 | 5.1 | 5.7 | 5.4 | 4.6 | 5.3 |
| CI of Z | 0.47 ± 0.03 | 0.36 ± 0.06 | 0.51 ± 0.06 | 0.52 ± 0.04 | 0.48 ± 0.03 | 0.49 ± 0.05 | 0.02 ± 0.03 | 0.29 ± 0.05 | 0.29 ± 0.05 |
| CI of W | 0.50 ± 0.06 | 0.50 ± 0.07 | 0.35 ± 0.04 | 0.34 ± 0.04 | 0.30 ± 0.04 | 0.24 ± 0.04 | 0.39 ± 0.03 | 0.16 ± 0.04 | 0.17 ± 0.04 |
| Asynapsis (A) or partial synapsis (P) | |||||||||
| Synapsis | A/P | P | P | A | A | A | A | A | A |
| N | 25 | 6 | 28 | 19 | 43 | 59 | 16 | 28 | 15 |
| Length of Z, µm | 17.1 ± 4.1 | 29.5 ± 6.8 | 23.0 ± 5.7 | 23.0 ± 4.6 | 14.1 ± 2.1 | 16.5 ± 4.4 | 16.7 ± 3.8 | 11.9 ± 3.6 | 15.8 ± 2.3 |
| CI of Z | 0.45 ± 0.04 | 0.35 ± 0.04 | 0.46 ± 0.07 | 0.44 ± 0.05 | 0.48 ± 0.01 | 0.47 ± 0.02 | 0.03 ± 0.02 | 0.31 ± 0.05 | 0.32 ± 0.02 |
| Length of W, µm | 8.68 ± 1.7 | 10.2 ± 3.1 | 16.1 ± 2.9 | 9.9 ± 2.1 | 7.6 ± 1.0 | 8.8 ± 1.9 | 11.3 ± 1.8 | 7.0 ± 1.41 | 4.7 ± 0.8 |
| CI of W | 0.44 ± 0.05 | 0.47 ± 0.07 | 0.36 ± 0.07 | 0.36 ± 0.04 | 0.28 ± 0.04 | 0.28 ± 0.05 | 0.41 ± 0.04 | 0.16 ± 0.04 | 0.19 ± 0.05 |
| Distance to MLH1 focus from the telomere in completely synapsed ZW | |||||||||
| N | 57 | 50 | 89 | 150 | 61 | 128 | 80 | 152 | 57 |
| Average distance, µm | 0.12±0.18 | 0.45 ± 0.17 | 0.79 ± 0.24 | 0.62 ± 0.17 | 0.45 ± 0.14 | 0.53 ± 0.13 | 0.37 ± 0.12 | 0.82 ± 0.26 | 0.37 ± 16 |
| Average distance ***, % | 1.0 ± 1.5 | 3.4 ± 1.3 | 5.2 ± 1.6 | 5.1 ± 1.5 | 5.0 ± 1.5 | 4.8 ± 1.2 | 3.3 ± 1.3 | 9.0 ± 2.9 | 3.6 ± 1.6 |
| Min-max distance, µm | 0–0.58 | 0.10–0.83 | 0.31–1.85 | 0.18–0.97 | 0.16–0.89 | 0.27–0.87 | 0.15–0.68 | 0.20–1.70 | 0.17–1.24 |
| Min-max distance, % | 0–4.7 | 0.9–5.7 | 1.4–10.4 | 1.2–8.4 | 1.5–8.3 | 2.5–8.4 | 1.0–6.6 | 2.0–15.6 | 1.4–12.5 |
| % of ZW occupied by MLH1 foci | 4.7 | 4.8 | 9 | 7.2 | 6.8 | 5.9 | 5.6 | 13.6 | 11.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Torgasheva, A.; Malinovskaya, L.; Zadesenets, K.S.; Slobodchikova, A.; Shnaider, E.; Rubtsov, N.; Borodin, P. Highly Conservative Pattern of Sex Chromosome Synapsis and Recombination in Neognathae Birds. Genes 2021, 12, 1358. https://doi.org/10.3390/genes12091358
Torgasheva A, Malinovskaya L, Zadesenets KS, Slobodchikova A, Shnaider E, Rubtsov N, Borodin P. Highly Conservative Pattern of Sex Chromosome Synapsis and Recombination in Neognathae Birds. Genes. 2021; 12(9):1358. https://doi.org/10.3390/genes12091358
Chicago/Turabian StyleTorgasheva, Anna, Lyubov Malinovskaya, Kira S. Zadesenets, Anastasia Slobodchikova, Elena Shnaider, Nikolai Rubtsov, and Pavel Borodin. 2021. "Highly Conservative Pattern of Sex Chromosome Synapsis and Recombination in Neognathae Birds" Genes 12, no. 9: 1358. https://doi.org/10.3390/genes12091358
APA StyleTorgasheva, A., Malinovskaya, L., Zadesenets, K. S., Slobodchikova, A., Shnaider, E., Rubtsov, N., & Borodin, P. (2021). Highly Conservative Pattern of Sex Chromosome Synapsis and Recombination in Neognathae Birds. Genes, 12(9), 1358. https://doi.org/10.3390/genes12091358