
Suggested Absence of Horizontal Transfer of Retrotransposons between Humans and Domestic Mammal Species

Abstract
:1. Introduction
2. Materials and Methods
3. Results and Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bourque, G.; Burns, K.H.; Gehring, M.; Gorbunova, V.; Seluanov, A.; Hammell, M.; Imbeault, M.; Izsvák, Z.; Levin, H.L.; Macfarlan, T.S.; et al. Ten things you should know about transposable elements. Genome Biol. 2018, 19, 199. [Google Scholar] [CrossRef]
- Zhang, H.-H.; Peccoud, J.; Xu, M.-R.-X.; Zhang, X.-G.; Gilbert, C. Horizontal transfer and evolution of transposable elements in vertebrates. Nat. Commun. 2020, 11, 1362. [Google Scholar] [CrossRef] [Green Version]
- Cordaux, R.; Batzer, M.A. The impact of retrotransposons on human genome evolution. Nat. Rev. Genet. 2009, 10, 691–703. [Google Scholar] [CrossRef] [Green Version]
- Panaud, O. Horizontal transfers of transposable elements in eukaryotes: The flying genes. Comptes Rendus Biol. 2016, 339, 296–299. [Google Scholar] [CrossRef] [Green Version]
- Schaack, S.; Gilbert, C.; Feschotte, C. Promiscuous DNA: Horizontal transfer of transposable elements and why it matters for eukaryotic evolution. Trends Ecol. Evol. 2010, 25, 537–546. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallau, G.L.; Vieira, C.; Loreto, É.L.S. Genetic exchange in eukaryotes through horizontal transfer: Connected by the mobilome. Mob. DNA 2018, 9, 6. [Google Scholar] [CrossRef] [PubMed]
- Martin, W.F. Too Much Eukaryote LGT. BioEssays 2017, 39, 1700115. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.A.; Li, J.; Davidson, W.S.; Davies, P.L. Smelt was the likely beneficiary of an antifreeze gene laterally transferred between fishes. BMC Evol. Biol. 2012, 12, 190. [Google Scholar] [CrossRef] [Green Version]
- Ivancevic, A.M.; Walsh, A.M.; Kortschak, R.D.; Adelson, D.L. Jumping the fine LINE between species: Horizontal transfer of transposable elements in animals catalyses genome evolution. BioEssays 2013, 35, 1071–1082. [Google Scholar] [CrossRef]
- Dotto, B.R.; Carvalho, E.L.; Silva, A.F.; Duarte Silva, L.F.; Pinto, P.M.; Ortiz, M.F.; Wallau, G.L. HTT-DB: Horizontally transferred transposable elements database. Bioinformatics 2015, 31, 2915–2917. [Google Scholar] [CrossRef] [Green Version]
- Silva, J.C.; Loreto, E.L.; Clark, J.B. Factors that affect the horizontal transfer of transposable elements. Curr. Issues Mol. Biol. 2004, 6, 57–71. Available online: https://www.ncbi.nlm.nih.gov/pubmed/14632259 (accessed on 4 June 2021). [PubMed]
- Gilbert, C.; Feschotte, C. Horizontal acquisition of transposable elements and viral sequences: Patterns and consequences. Curr. Opin. Genet. Dev. 2018, 49, 15–24. [Google Scholar] [CrossRef]
- Thomas, J.; Phillips, C.D.; Baker, R.J.; Pritham, E.J. Rolling-circle transposons catalyze genomic innovation in a mammalian lineage. Genome Biol. Evol. 2014, 6, 2595–2610. [Google Scholar] [CrossRef] [Green Version]
- Wallau, G.L.; Capy, P.; Loreto, E.; Le Rouzic, A.; Hua-Van, A. VHICA, a New Method to Discriminate between Vertical and Horizontal Transposon Transfer: Application to the Mariner Family within Drosophila. Mol. Biol. Evol. 2016, 33, 1094–1109. [Google Scholar] [CrossRef] [Green Version]
- Daniels, S.B.; Peterson, K.R.; Strausbaugh, L.D.; Kidwell, M.G.; Chovnick, A. Evidence for horizontal transmission of the P transposable element between Drosophila species. Genetics 1990, 124, 339–355. Available online: https://www.ncbi.nlm.nih.gov/pubmed/2155157 (accessed on 15 June 2021). [CrossRef] [PubMed]
- Peccoud, J.; Loiseau, V.; Cordaux, R.; Gilbert, C. Massive horizontal transfer of transposable elements in insects. Proc. Natl. Acad. Sci. USA 2017, 114, 4721–4726. [Google Scholar] [CrossRef] [Green Version]
- Ivancevic, A.M.; Kortschak, R.D.; Bertozzi, T.; Adelson, D.L. Horizontal transfer of BovB and L1 retrotransposons in eukaryotes. Genome Biol. 2018, 19, 85. [Google Scholar] [CrossRef] [Green Version]
- Walsh, A.M.; Kortschak, R.D.; Gardner, M.G.; Bertozzi, T.; Adelson, D.L. Widespread horizontal transfer of retrotransposons. Proc. Natl. Acad. Sci. USA 2013, 110, 1012–1016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, C.; Chateigner, A.; Ernenwein, L.; Barbe, V.; Bézier, A.; Herniou, E.A.; Cordaux, R. Population genomics supports baculoviruses as vectors of horizontal transfer of insect transposons. Nat. Commun. 2014, 5, 3348. [Google Scholar] [CrossRef] [Green Version]
- Larson, G.; Fuller, D.Q. The Evolution of Animal Domestication. Annu. Rev. Ecol. Evol. Syst. 2014, 45, 115–136. [Google Scholar] [CrossRef] [Green Version]
- MacEachern, S.; Hayes, B.; McEwan, J.; Goddard, M. An examination of positive selection and changing effective population size in Angus and Holstein cattle populations (Bos taurus) using a high density SNP genotyping platform and the contribution of ancient polymorphism to genomic diversity in Domestic cattle. BMC Genom. 2009, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- Suh, A.; Churakov, G.; Ramakodi, M.P.; Platt, R.N.; Jurka, J.; Kojima, K.K.; Caballero, J.; Smit, A.F.; Vliet, K.A.; Hoffmann, F.G.; et al. Multiple lineages of ancient CR1 retroposons shaped the early genome evolution of amniotes. Genome Biol. Evol. 2014, 7, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Waterston, R.H.; Pachter, L. Initial sequencing and comparative analysis of the mouse genome. Nature 2002, 420, 520–562. [Google Scholar] [CrossRef] [PubMed]
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, D.A.; Pagan, H.J.T.; Thompson, M.L.; Stevens, R.D. Bats with hATs: Evidence for recent DNA transposon activity in genus Myotis. Mol. Biol. Evol. 2007, 24, 632–639. [Google Scholar] [CrossRef] [PubMed]
- Renault, S.; Genty, M.; Gabori, A.; Boisneau, C.; Esnault, C.; Dugé de Bernonville, T.; Augé-Gouillou, C. The epigenetic regulation of HsMar1, a human DNA transposon. BMC Genet. 2019, 20, 17. [Google Scholar] [CrossRef] [PubMed]
- Cordaux, R.; Udit, S.; Batzer, M.A.; Feschotte, C. Birth of a chimeric primate gene by capture of the transposase gene from a mobile element. Proc. Natl. Acad. Sci. USA 2006, 103, 8101–8106. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Terrones, D.; Torres-Padilla, M.-E. Nimble and Ready to Mingle: Transposon Outbursts of Early Development. Trends Genet. 2018, 34, 806–820. [Google Scholar] [CrossRef]
- Feschotte, C.; Pritham, E.J. DNA transposons and the evolution of eukaryotic genomes. Annu. Rev. Genet. 2007, 41, 331–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagès, H.; Aboyoun, P.; Gentleman, R.; DebRoy, S. Biostrings: Efficient Manipulation of Biological Strings. R Package Version 2.54.0. 2019. Available online: https://rdrr.io/bioc/Biostrings/ (accessed on 21 May 2021).
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Subramanian, S. Mutation rates in mammalian genomes. Proc. Natl. Acad. Sci. USA 2002, 99, 803–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blanchette, M.; Kent, W.J.; Riemer, C.; Elnitski, L.; Smit, A.F.A.; Roskin, K.M.; Baertsch, R.; Rosenbloom, K.; Clawson, H.; Green, E.D.; et al. Aligning multiple genomic sequences with the threaded blockset aligner. Genome Res. 2004, 14, 708–715. [Google Scholar] [CrossRef] [Green Version]
- Drew, A.C.; Brindley, P.J. A retrotransposon of the non-long terminal repeat class from the human blood fluke Schistosoma mansoni. Similarities to the chicken-repeat-1-like elements of vertebrates. Mol. Biol. Evol. 1997, 14, 602–610. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandergon, T.L.; Reitmant, M. Evolution of Chicken Repeat (CRl) Elements: Evidence for Ancient Subfamilies and Multiple Progenitors. Mol. Biol. Evol. 1994, 11, 886–898. [Google Scholar]
- Axelsson, E.; Ratnakumar, A.; Arendt, M.-L.; Maqbool, K.; Webster, M.T.; Perloski, M.; Liberg, O.; Arnemo, J.M.; Hedhammar, Å.; Lindblad-Toh, K. The genomic signature of dog domestication reveals adaptation to a starch-rich diet. Nature 2013, 495, 360–364. [Google Scholar] [CrossRef] [PubMed]
- Pitt, D.; Sevane, N.; Nicolazzi, E.L.; MacHugh, D.E.; Park, S.D.E.; Colli, L.; Martinez, R.; Bruford, M.W.; Orozco-terWengel, P. Domestication of cattle: Two or three events? Evol. Appl. 2019, 12, 123–136. [Google Scholar] [CrossRef]
- Schubert, M.; Jónsson, H.; Chang, D.; Der Sarkissian, C.; Ermini, L.; Ginolhac, A.; Albrechtsen, A.; Dupanloup, I.; Foucal, A.; Petersen, B.; et al. Prehistoric genomes reveal the genetic foundation and cost of horse domestication. Proc. Natl. Acad. Sci. USA 2014, 111, E5661–E5669. [Google Scholar] [CrossRef] [Green Version]
- Chessa, B.; Pereira, F.; Arnaud, F.; Amorim, A.; Goyache, F.; Mainland, I.; Kao, R.R.; Pemberton, J.M.; Beraldi, D.; Stear, M.J.; et al. Revealing the history of sheep domestication using retrovirus integrations. Science 2009, 324, 532–536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stahl, P.W. Animal Domestication in South America. In The Handbook of South American Archaeology; Silverman, H., Isbell, W.H., Eds.; Springer: New York, NY, USA, 2008; pp. 121–130. [Google Scholar] [CrossRef]
- Hu, Y.; Hu, S.; Wang, W.; Wu, X.; Marshall, F.B.; Chen, X.; Hou, L.; Wang, C. Earliest evidence for commensal processes of cat domestication. Proc. Natl. Acad. Sci. USA 2014, 111, 116–120. [Google Scholar] [CrossRef] [Green Version]
- Kadwell, M.; Fernandez, M.; Stanley, H.F.; Baldi, R.; Wheeler, J.C.; Rosadio, R.; Bruford, M.W. Genetic analysis reveals the wild ancestors of the llama and the alpaca. Proc. Biol. Sci. 2001, 268, 2575–2584. [Google Scholar] [CrossRef] [Green Version]
- Thornton, P.C.; Wright, P.A.; Sacra, P.J.; Goodiert, T.E.W. The ferret, Mustela putorius furo, as a new species in toxicology. Lab. Anim. 1979, 13, 119–124. [Google Scholar] [CrossRef]
- Ji, R.; Cui, P.; Ding, F.; Geng, J.; Gao, H.; Zhang, H.; Yu, J.; Hu, S.; Meng, H. Monophyletic origin of domestic bactrian camel (Camelus bactrianus) and its evolutionary relationship with the extant wild camel (Camelus bactrianus ferus). Anim. Genet. 2009, 40, 377–382. [Google Scholar] [CrossRef] [Green Version]
- Orlando, L. Back to the roots and routes of dromedary domestication. Proc. Natl. Acad. Sci. USA 2016, 113, 6588–6590. [Google Scholar] [CrossRef] [Green Version]
- Utsunomiya, Y.T.; Milanesi, M.; Fortes, M.R.S.; Porto-Neto, L.R.; Utsunomiya, A.T.H.; Silva, M.V.G.B.; Garcia, J.F.; Ajmone-Marsan, P. Genomic clues of the evolutionary history of Bos indicus cattle. Anim. Genet. 2019, 50, 557–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kordis, D.; Gubensek, F. Horizontal transfer of non-LTR retrotransposons in vertebrates. Genetica 1999, 107, 121–128. [Google Scholar] [CrossRef]
- Galbraith, J.D.; Ludington, A.J.; Suh, A. New Environment, New Invaders—Repeated Horizontal Transfer of LINEs to Sea Snakes. Genome Biol. Evol. 2020, 12, 2370–2383. [Google Scholar] [CrossRef] [PubMed]
- Smit, A.F.; Riggs, A.D. MIRs are classic, tRNA-derived SINEs that amplified before the mammalian radiation. Nucleic Acids Res. 1995, 23, 98–102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirakawa, M.; Nishihara, H.; Kanehisa, M.; Okada, N. Characterization and evolutionary landscape of AmnSINE1 in Amniota genomes. Gene 2009, 441, 100–110. [Google Scholar] [CrossRef]
- Han, G.; Zhang, N.; Jiang, H.; Meng, X.; Qian, K.; Zheng, Y.; Xu, J.; Wang, J. Diversity of short interspersed nuclear elements (SINEs) in lepidopteran insects and evidence of horizontal SINE transfer between baculovirus and lepidopteran hosts. BMC Genom. 2021, 22, 226. [Google Scholar] [CrossRef]
- Wang, J.; Wang, A.; Han, Z.; Zhang, Z.; Li, F.; Li, X. Characterization of three novel SINE families with unusual features in Helicoverpa armigera. PLoS ONE 2012, 7, e31355. [Google Scholar] [CrossRef]
- Luchetti, A.; Mantovani, B. Rare horizontal transmission does not hide long-term inheritance of SINE highly conserved domains in the metazoan evolution. Curr. Zool. 2016, 62, 667–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamada, H.; Matthews, S.G. Prenatal programming of stress responsiveness and behaviours: Progress and perspectives. J. Neuroendocrinol. 2019, 31, e12674. [Google Scholar] [CrossRef]
- Palazzo, A.; Lorusso, P.; Miskey, C.; Walisko, O.; Gerbino, A.; Marobbio, C.M.T.; Ivics, Z.; Marsano, R.M. Transcriptionally promiscuous “blurry” promoters in Tc1/mariner transposons allow transcription in distantly related genomes. Mob. DNA 2019, 10, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palazzo, A.; Caizzi, R.; Viggiano, L.; Marsano, R.M. Does the Promoter Constitute a Barrier in the Horizontal Transposon Transfer Process? Insight from Bari Transposons. Genome Biol. Evol. 2017, 9, 1637–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
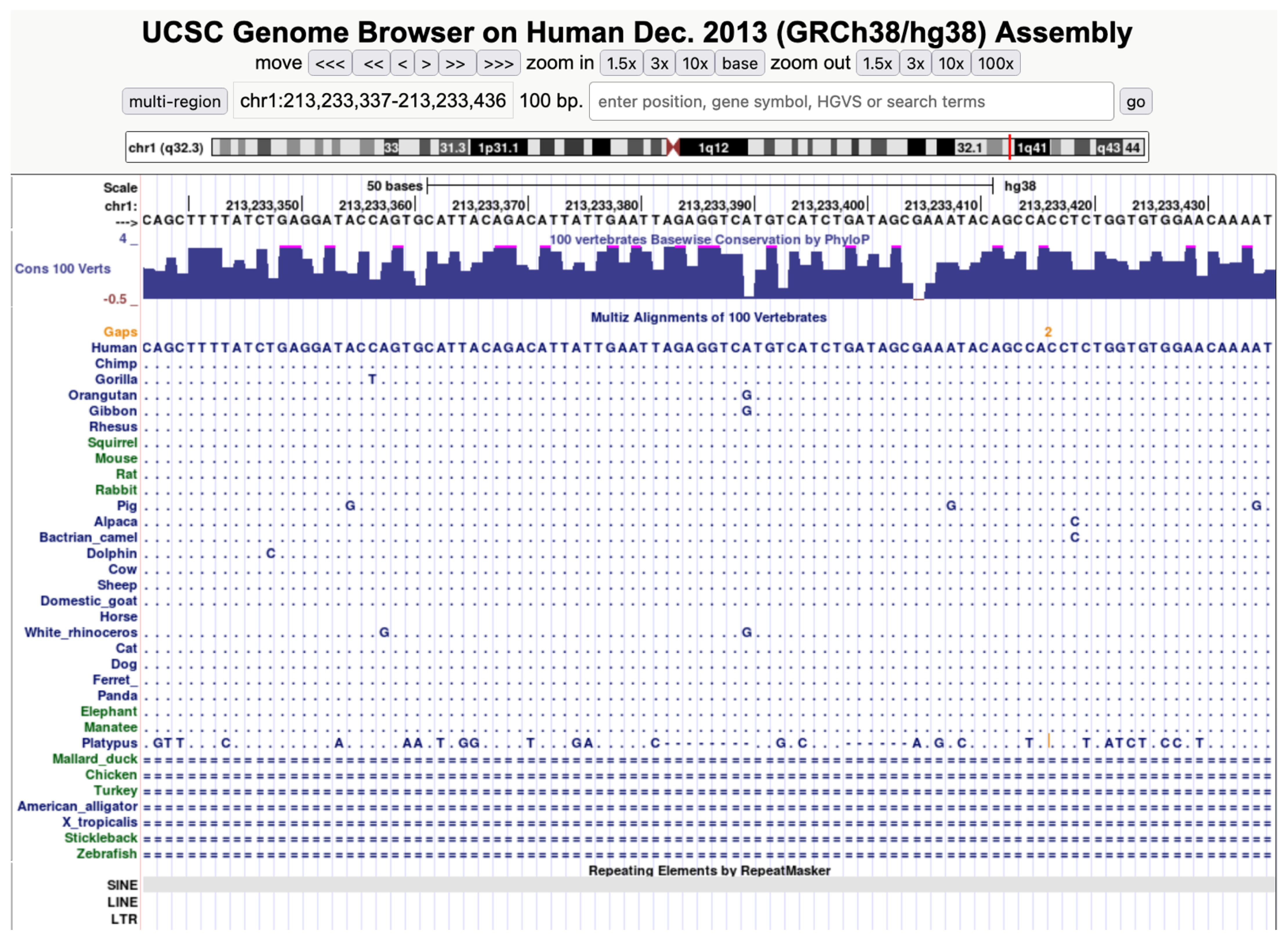

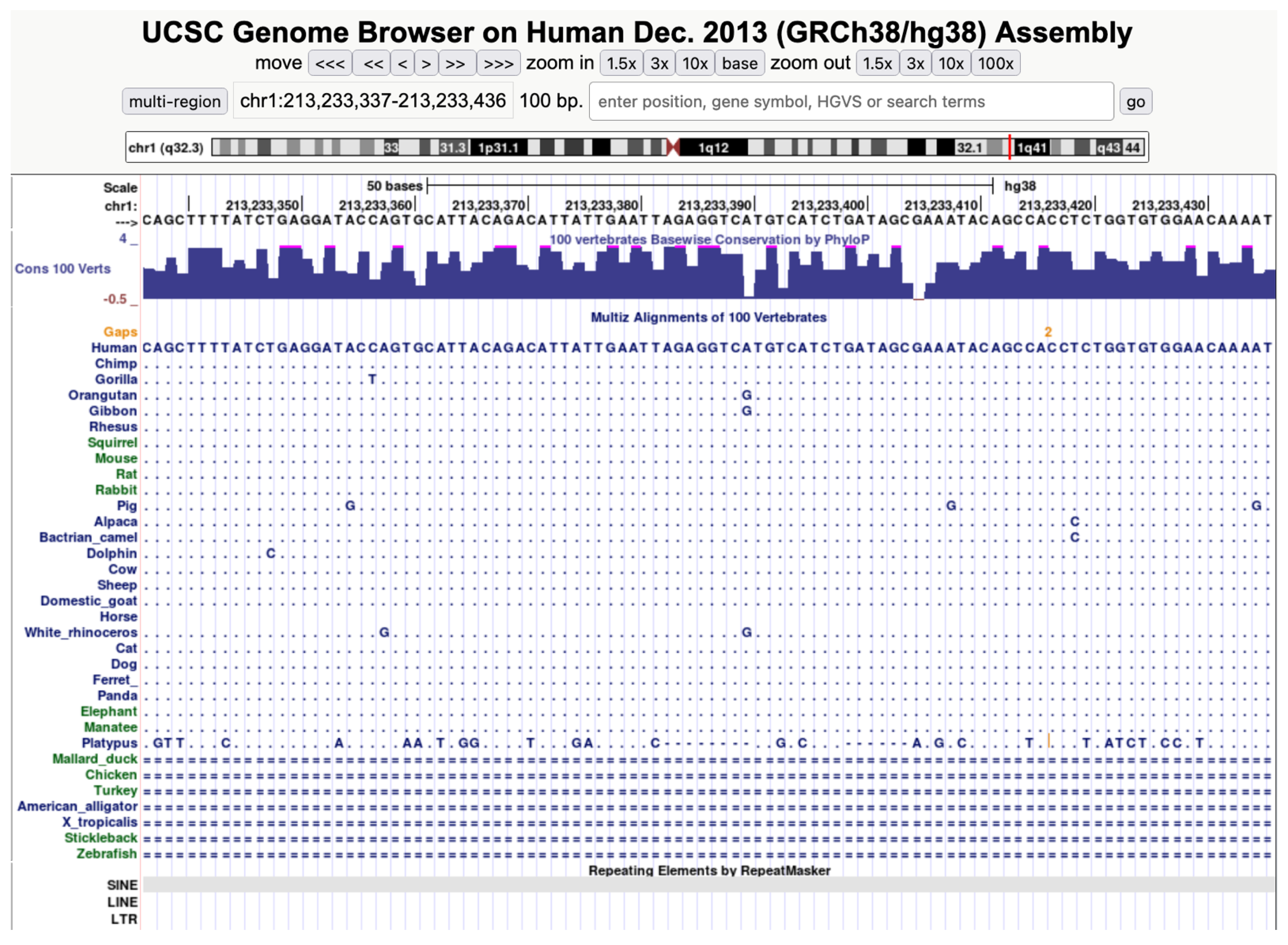{kind=link}
| Animal | Genome | Approximate Domestication (kya) | n Queried (mRNA) | n Hits (mRNA) | Mean Hit Length (mRNA) | n Queried (TE) | n Hits (TE) | Mean Hit Length (TE) |
|---|---|---|---|---|---|---|---|---|
| Human | hg38 | 3,402,790 | ||||||
| Dog | canFam5 | 12–33 [36] | 1,748,840 | 19 | 138 | |||
| Cow | bosTau9 | >10 [37] | 14,530 | 132 | 220 | 3,117,396 | 12 | 135 |
| Horse | equCab3 | 5.5 [38] | 1878 | 117 | 228 | 2,034,000 | 21 | 128 |
| Sheep | oviAri4 | 11 [39] | 1018 | 25 | 85.2 | 2,960,232 | 9 | 135 |
| Pig | susScr11 | 9 [20] | 4851 | 98 | 90 | 1,789,659 | 13 | 144 |
| Guinea pig | cavPor3 | 5 [40] | 491 | 51 | 121 | 1,721,346 | 5 | 122 |
| Cat | felCat9 | 4 [41] | 419 | 0 | 1,855,102 | 20 | 138 | |
| Alpaca | vicPac2 | 7 [42] | 17 | 0 | 1,739,851 | 18 | 140 | |
| Ferret | musFur1 | 2 [43] | 2,545,632 | 4 | 125 | |||
| Bactrian camel | GCF_000767855.1 | 5 [44] | 1,586,346 | 15 | 134 | |||
| Dromedary | GCF_000803125.2 | 4 [45] | 1,663,540 | 18 | 134 | |||
| Goat | GCF_001704425.1 | 11 [39] | 2,153,565 | 9 | 133 | |||
| Zebu | GCF_000247795.1 | 8 [46] | 3,034,305 | 11 | 130 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wanner, N.M.; Faulk, C. Suggested Absence of Horizontal Transfer of Retrotransposons between Humans and Domestic Mammal Species. Genes 2021, 12, 1223. https://doi.org/10.3390/genes12081223
Wanner NM, Faulk C. Suggested Absence of Horizontal Transfer of Retrotransposons between Humans and Domestic Mammal Species. Genes. 2021; 12(8):1223. https://doi.org/10.3390/genes12081223
Chicago/Turabian StyleWanner, Nicole M., and Christopher Faulk. 2021. "Suggested Absence of Horizontal Transfer of Retrotransposons between Humans and Domestic Mammal Species" Genes 12, no. 8: 1223. https://doi.org/10.3390/genes12081223
APA StyleWanner, N. M., & Faulk, C. (2021). Suggested Absence of Horizontal Transfer of Retrotransposons between Humans and Domestic Mammal Species. Genes, 12(8), 1223. https://doi.org/10.3390/genes12081223