
Effective RNAi-Mediated Silencing of the Mismatch Repair MSH2 Gene Induces Sterility of Tomato Plants but Not an Increase in Meiotic Recombination


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plants for the Experiments
2.2. Genetic Constructs for Plant Transformation
2.3. Genetic Transformation of the Tomato Plants
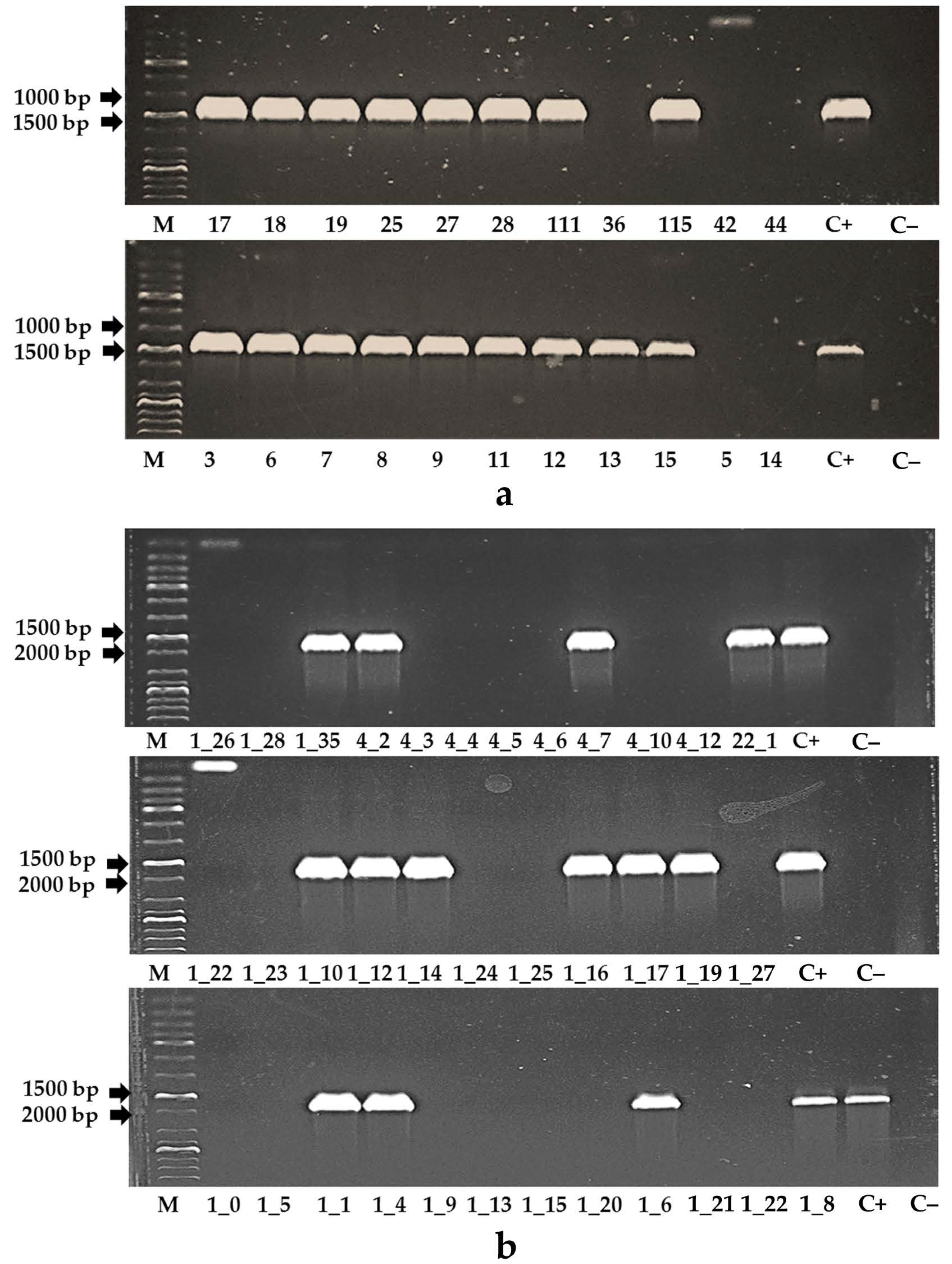
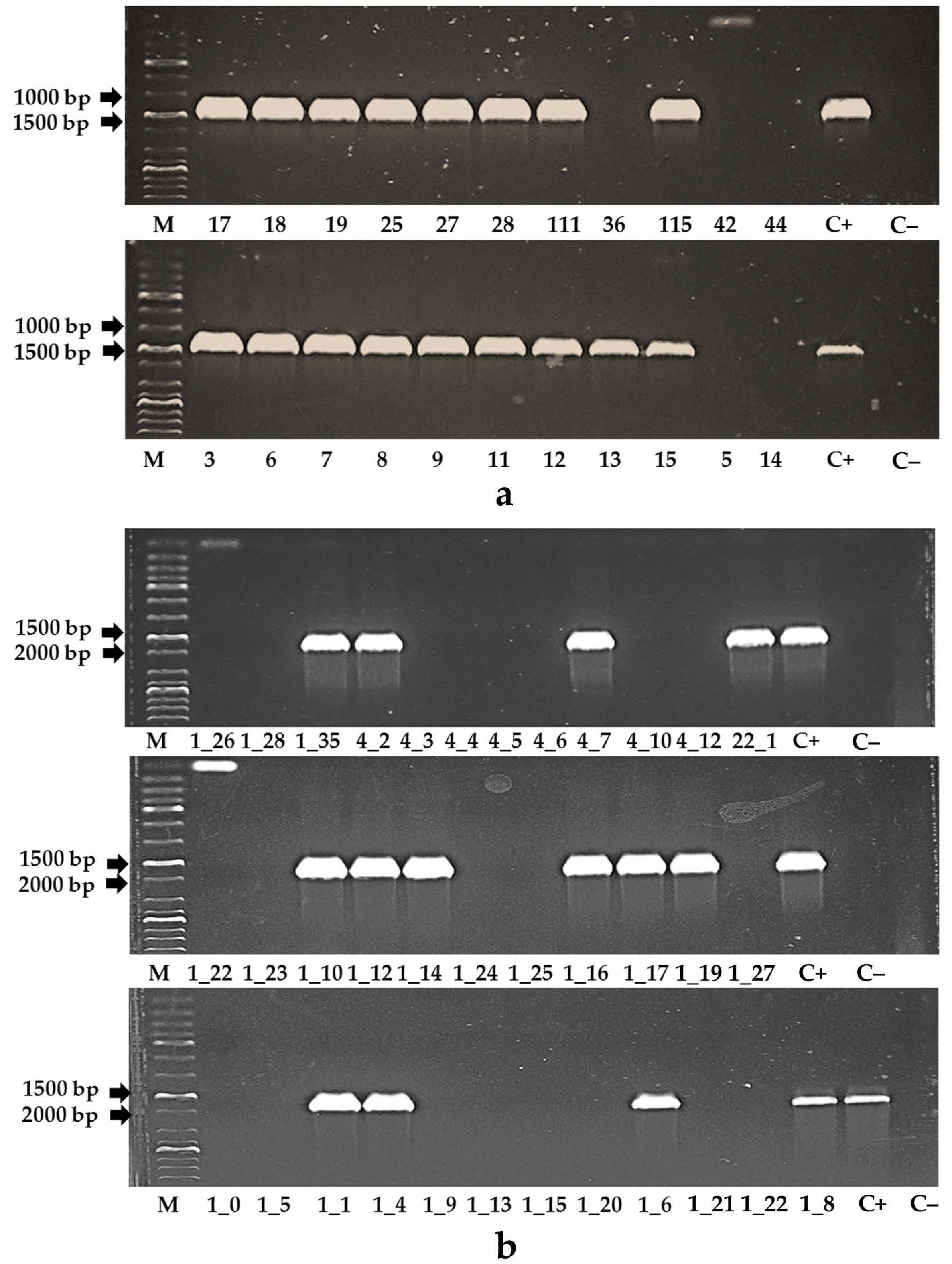
2.4. Analysis of Tomato Plants with Polymerase Chain Reaction
2.5. RNA Extraction
2.6. Statistical Data Processing
2.7. Pollen Viability
3. Results
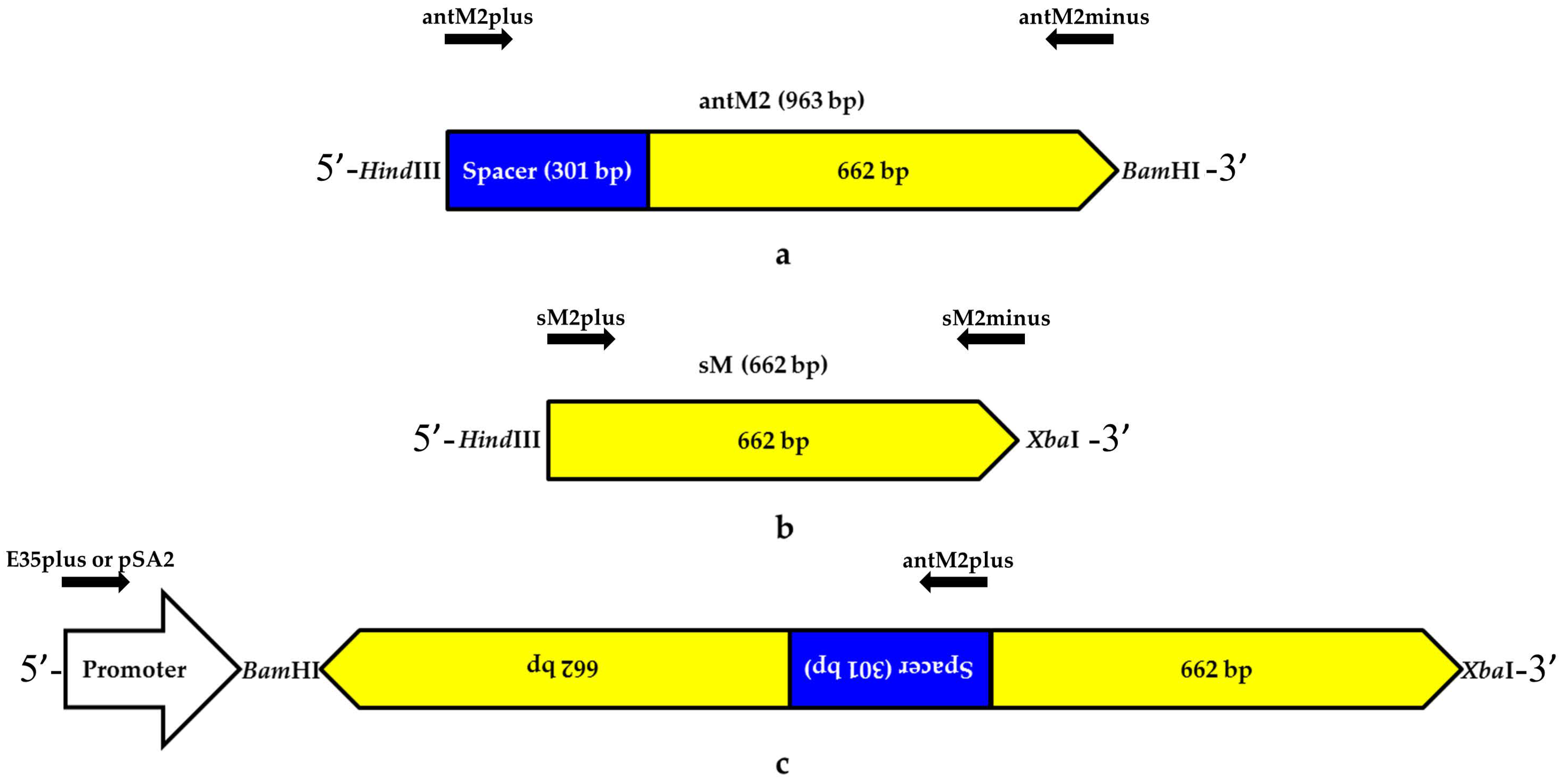
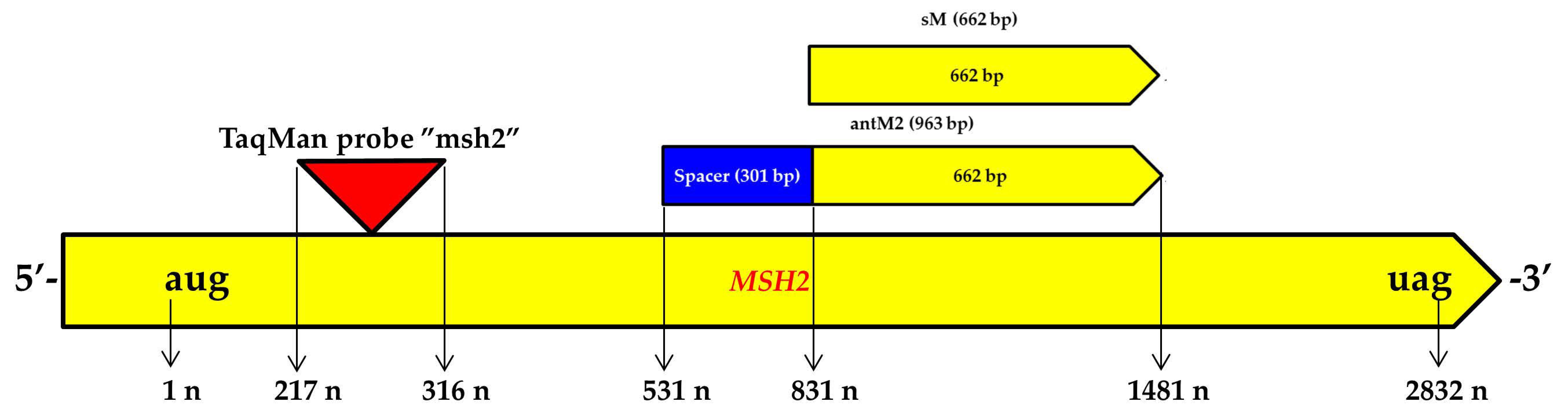
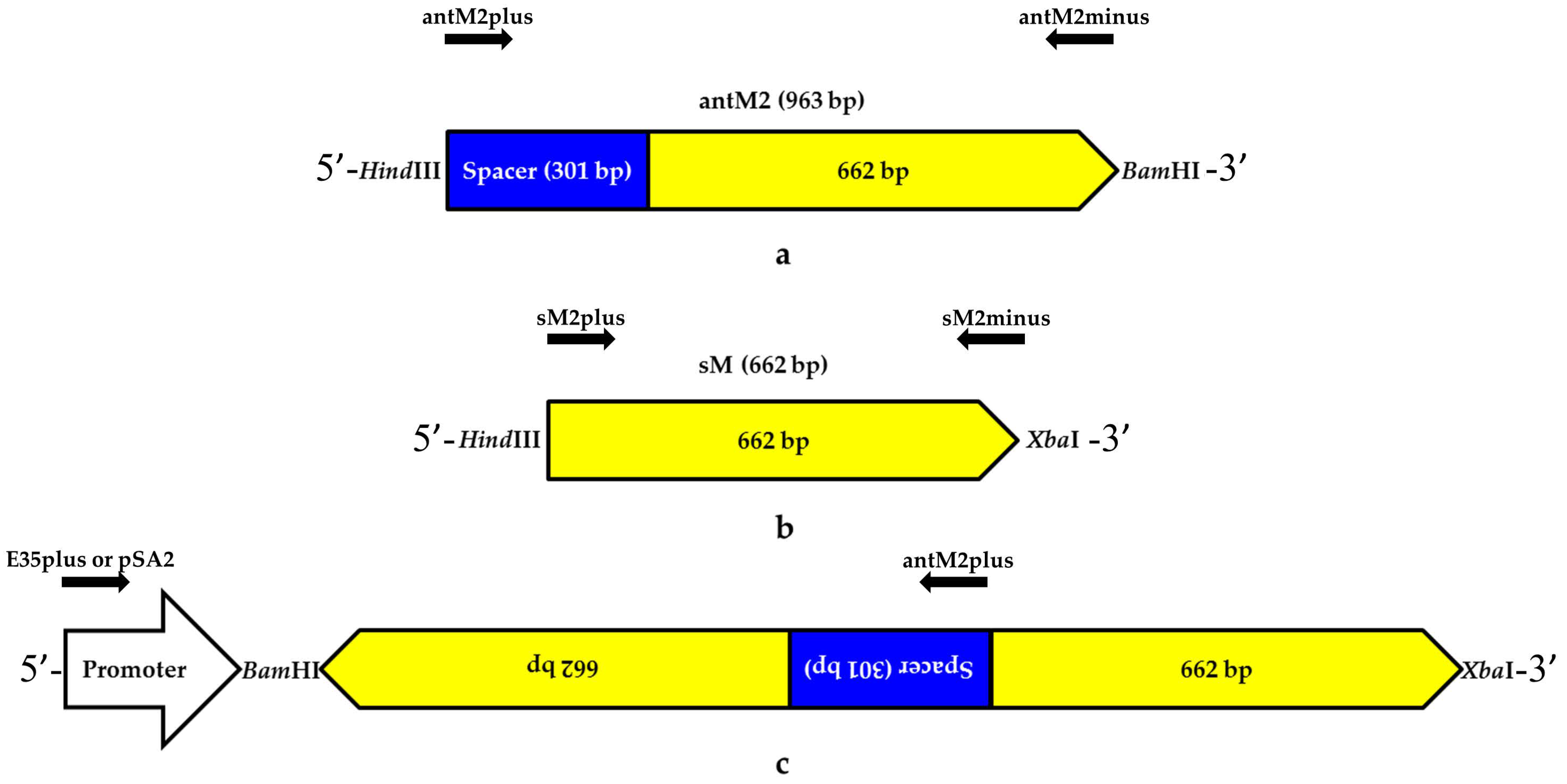
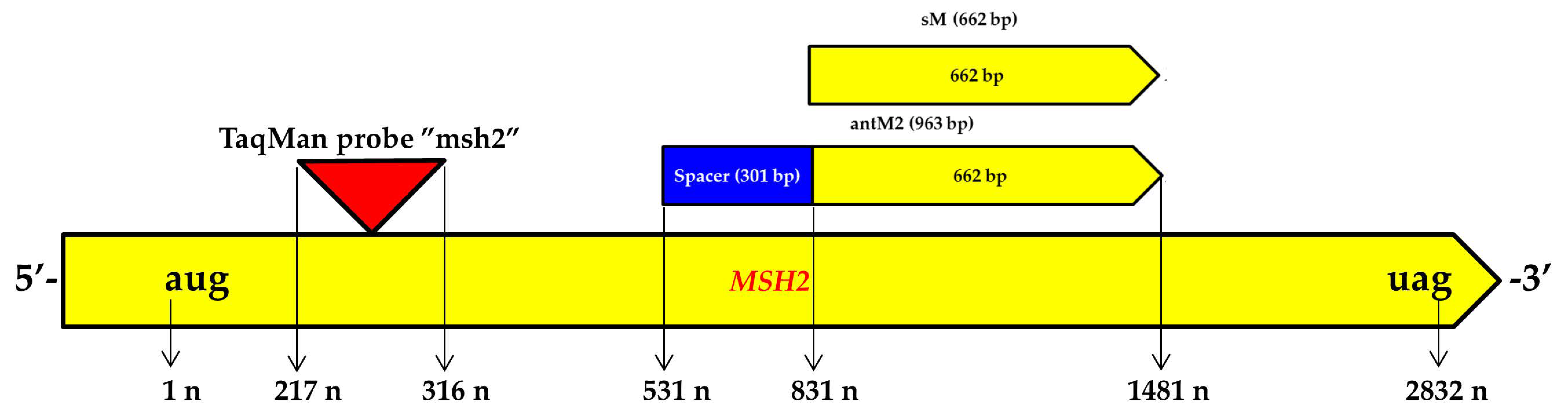
3.1. Genetic Constructs Created for RNA Interference Targeting the MSH2 Gene in Tomato Plants
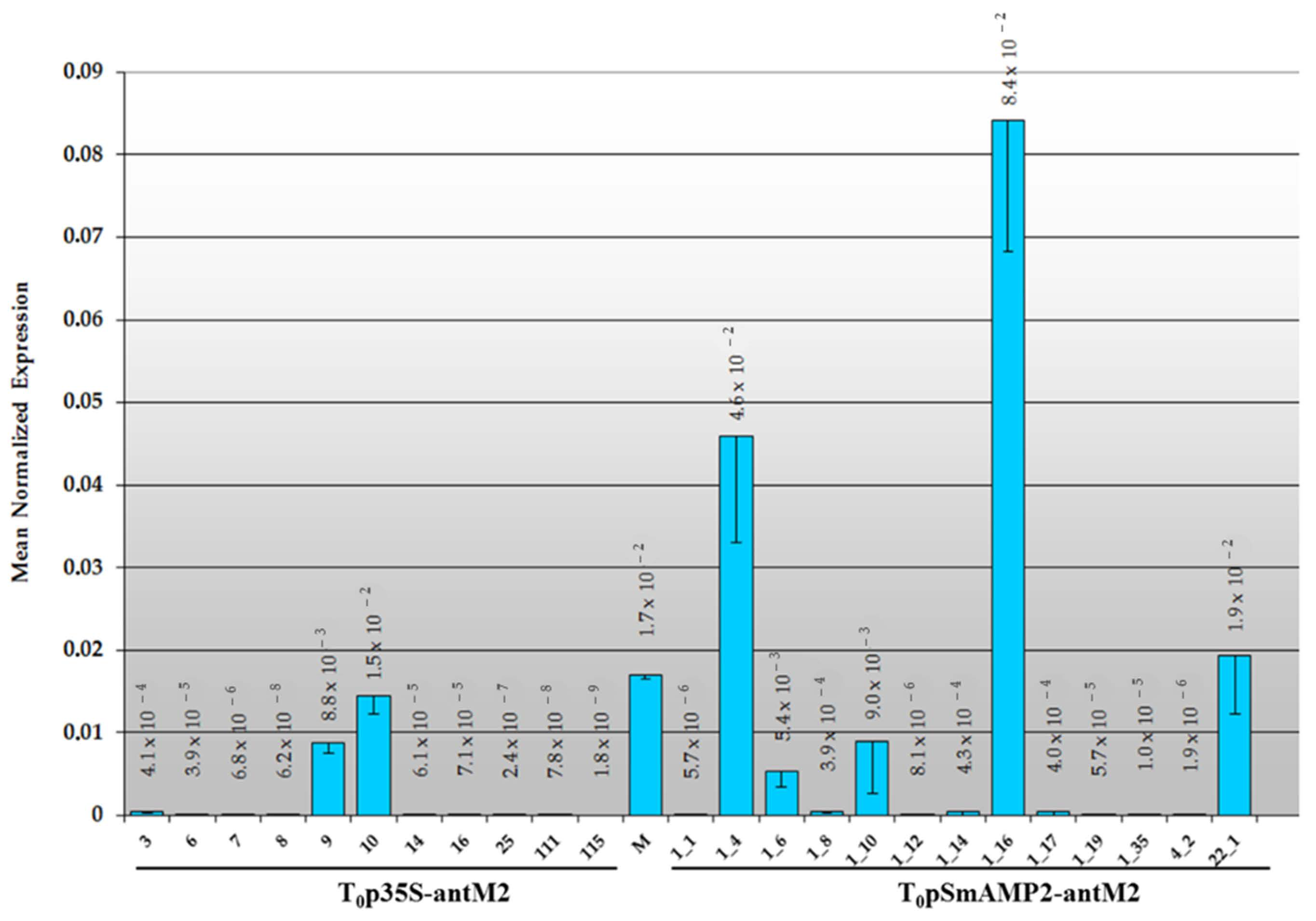
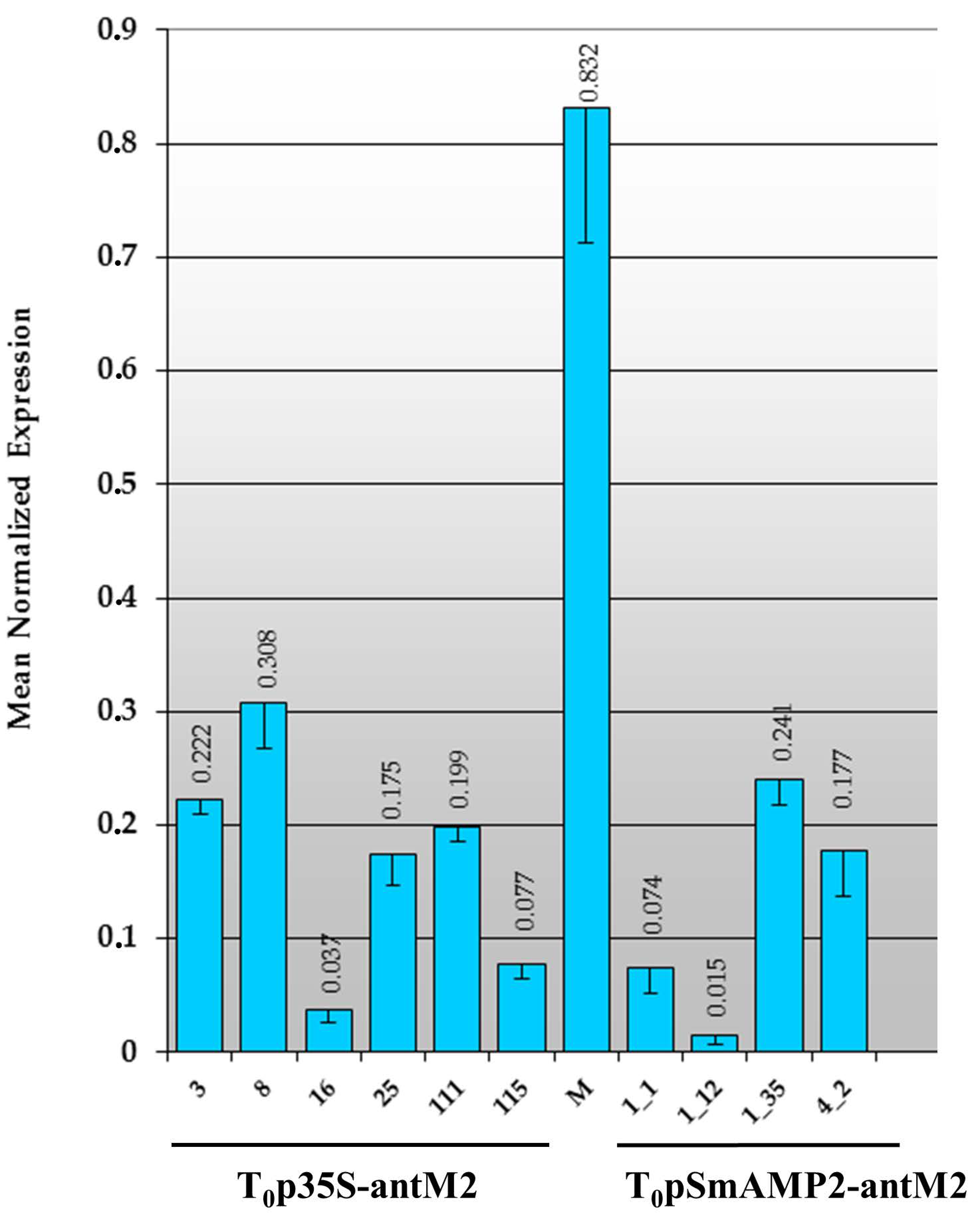
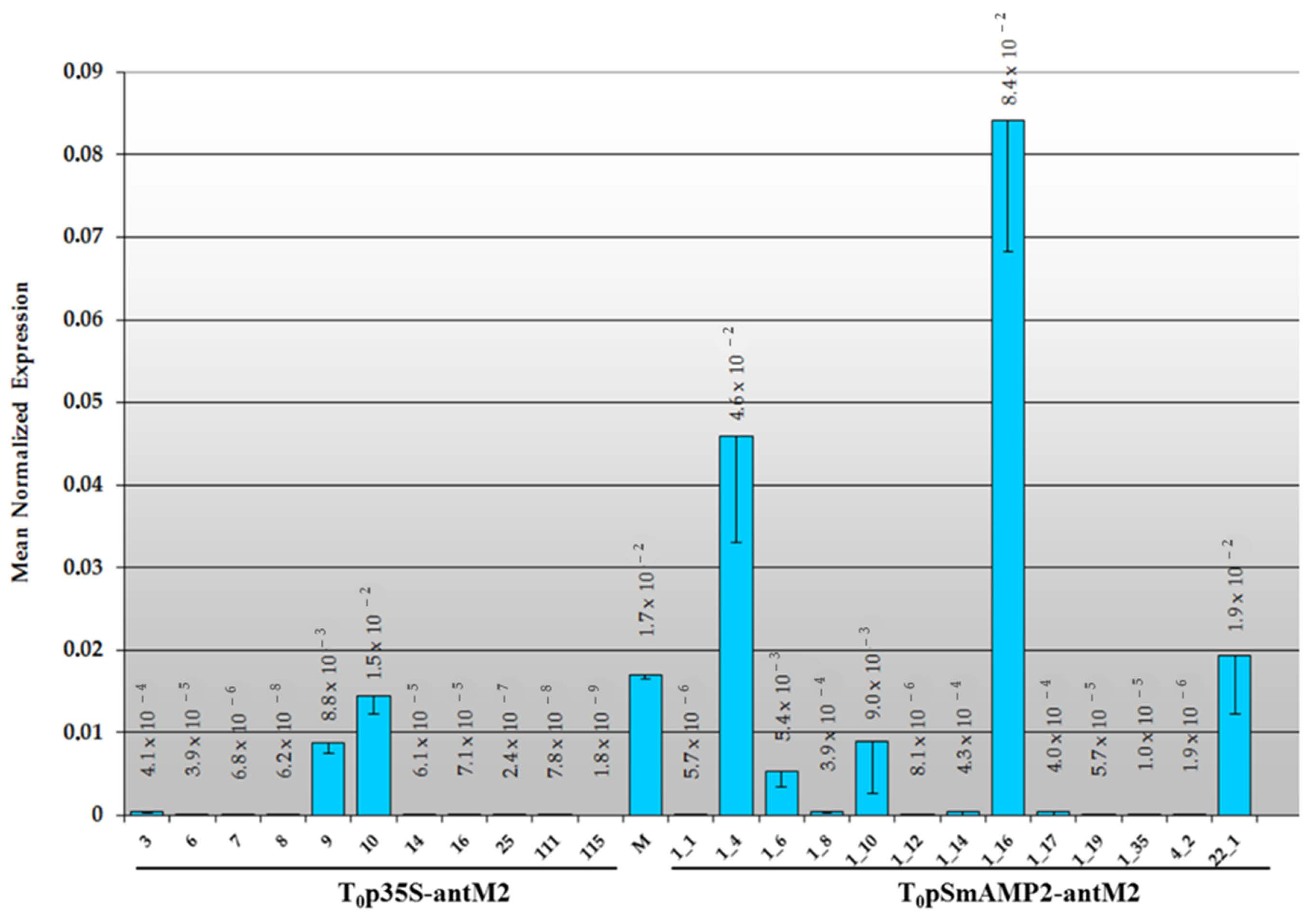
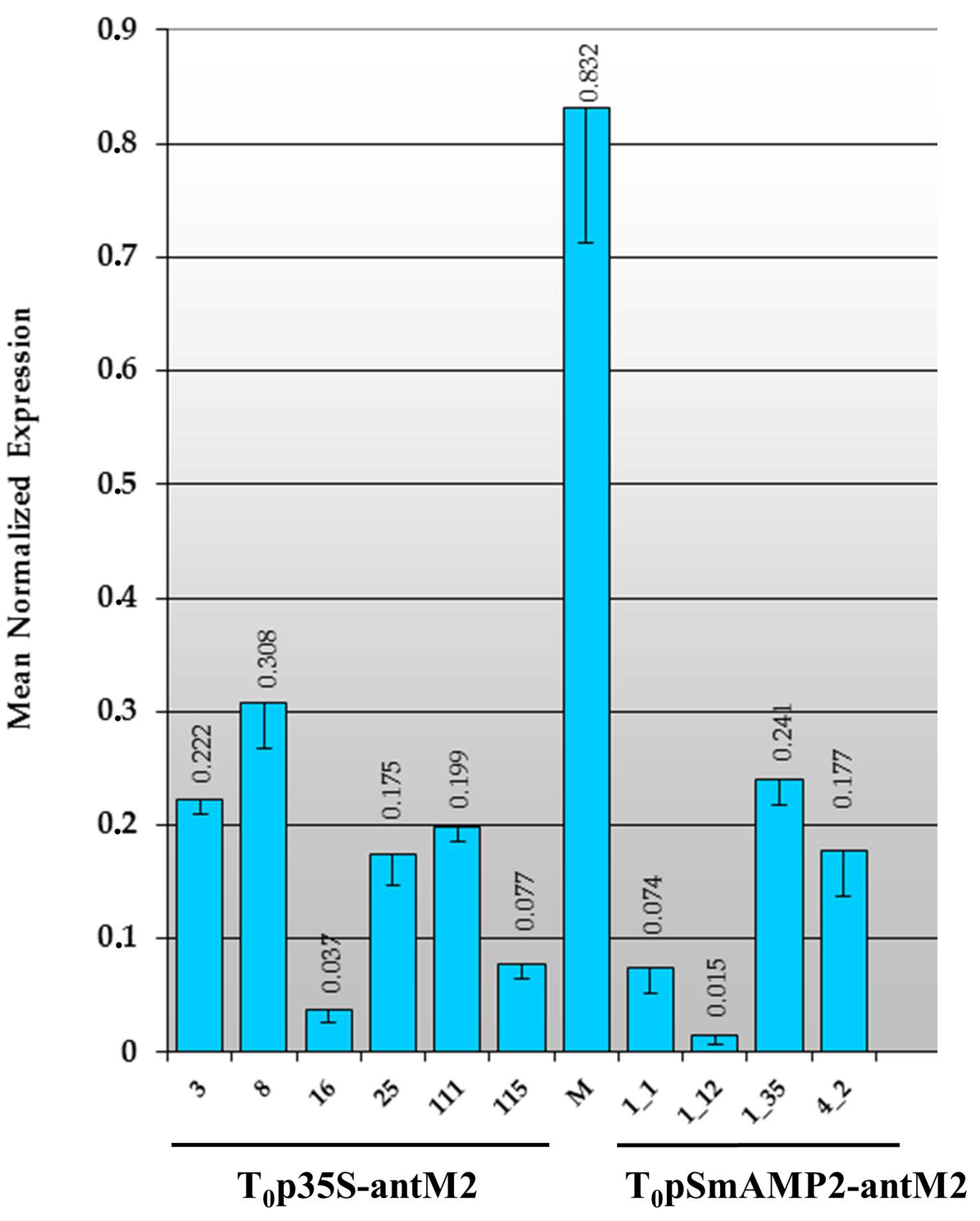
3.2. Primary Transformants of Tomato Plants Created with a Reduced Content of MSH2 Gene Transcript
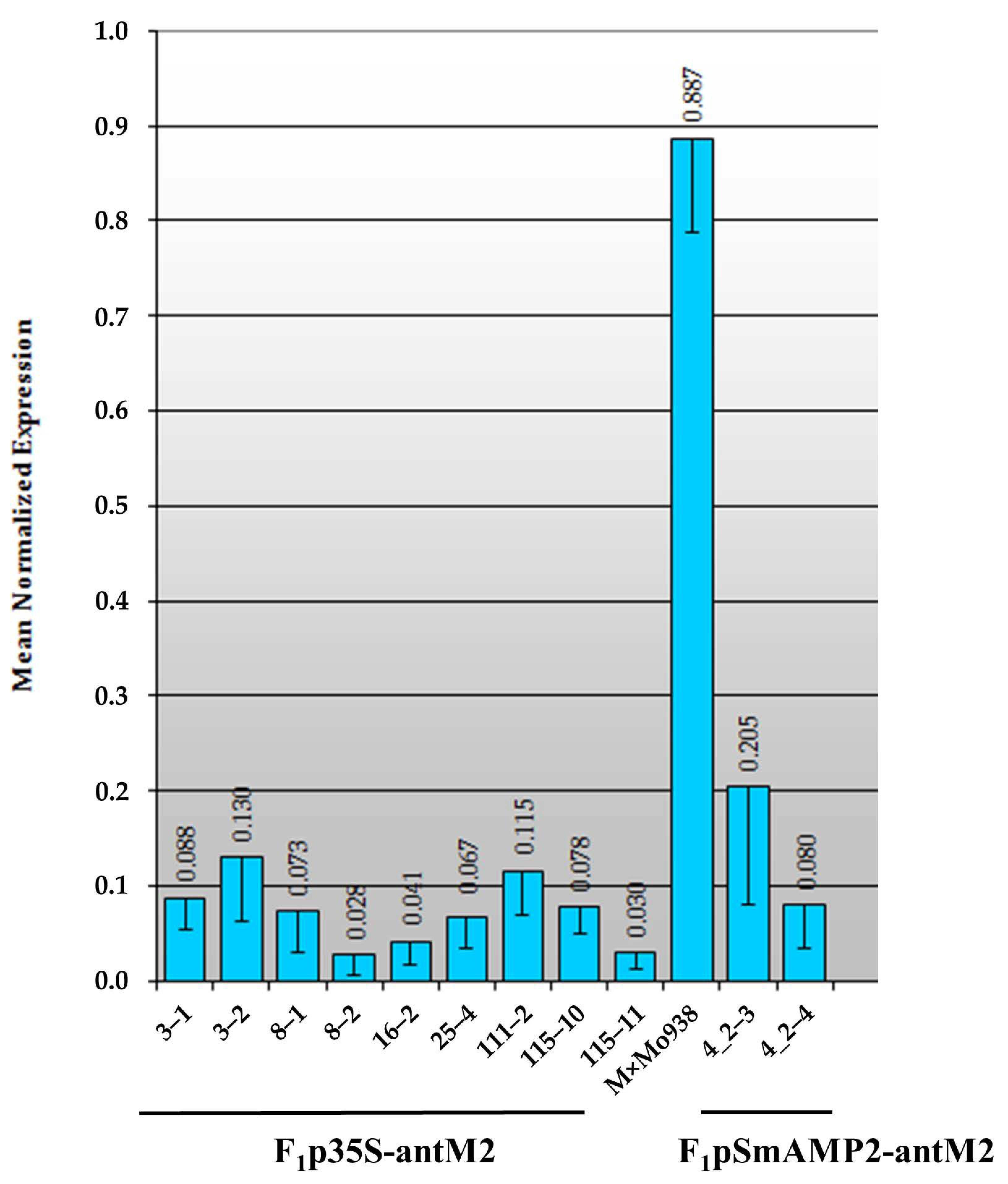
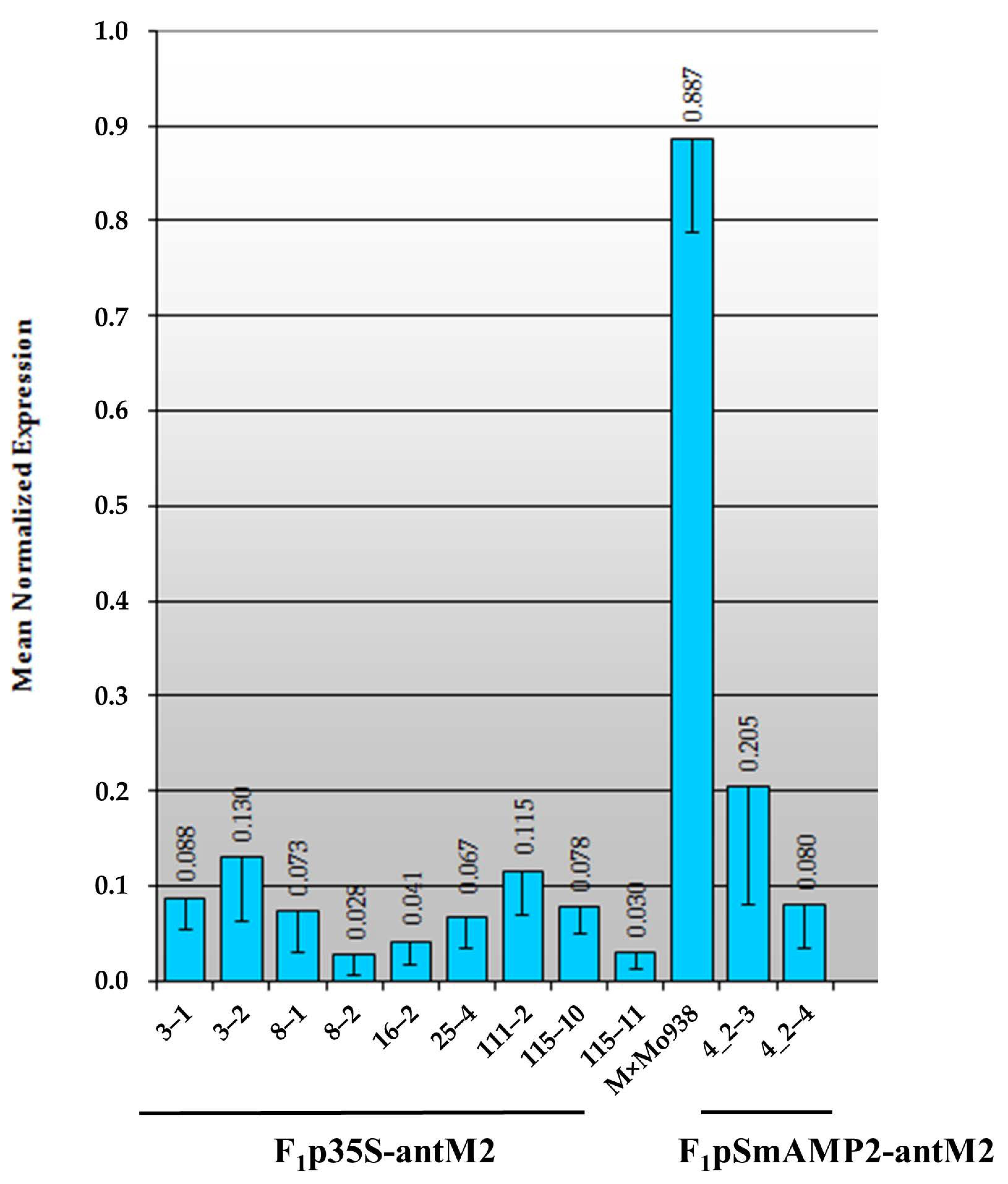
3.3. Assessment of the MSH2 Gene Transcript’s Silencing Efficiency in Tomato Hybrids
3.4. Analysis of the Meiotic Recombination Frequency between Marker Genes of Chromosome 2 in Tomato Hybrids with the MSH2 Transcript Silenced
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Cole, F.; Keeney, S.; Jasin, M. Preaching about the Converted: How Meiotic Gene Conversion Influences Genomic Diversity. Ann. N. Y. Acad. Sci. 2012, 1267, 95–102. [Google Scholar] [CrossRef] [PubMed]
- De Muyt, A.; Pereira, L.; Vezon, D.; Chelysheva, L.; Gendrot, G.; Chambon, A.; Lainé-Choinard, S.; Pelletier, G.; Mercier, R.; Nogué, F.; et al. A High Throughput Genetic Screen Identifies New Early Meiotic Recombination Functions in Arabidopsis thaliana. PLoS Genet. 2009, 5, e1000654. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijnker, E.; de Jong, H. Managing Meiotic Recombination in Plant Breeding. Trends Plant Sci. 2008, 13, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Lambing, C.; Franklin, F.C.H.; Wang, C.-J.R. Understanding and Manipulating Meiotic Recombination in Plants. Plant Physiol. 2017, 173, 1530–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tam, S.M.; Hays, J.B.; Chetelat, R.T. Effects of Suppressing the DNA Mismatch Repair System on Homeologous Recombination in Tomato. Theor. Appl. Genet. 2011, 123, 1445–1458. [Google Scholar] [CrossRef] [PubMed]
- Mézard, C.; Tagliaro Jahns, M.; Grelon, M. Where to Cross? New Insights into the Location of Meiotic Crossovers. Trends Genet. 2015, 31, 393–401. [Google Scholar] [CrossRef]
- Page, S.L.; Hawley, R.S. Chromosome Choreography: The Meiotic Ballet. Science 2003, 301, 785–789. [Google Scholar] [CrossRef] [Green Version]
- Culligan, K.M.; Hays, J.B. Arabidopsis MutS Homologs—AtMSH2, AtMSH3, AtMSH6, and a Novel AtMSH7—Form Three Distinct Protein Heterodimers with Different Specificities for Mismatched DNA. Plant Cell 2000, 12, 991–1002. [Google Scholar] [CrossRef] [Green Version]
- Sachadyn, P. Conservation and Diversity of MutS Proteins. Mutat. Res./Fundam. Mol. Mech. Mutagen. 2010, 694, 20–30. [Google Scholar] [CrossRef]
- Acharya, S.; Wilson, T.; Gradia, S.; Kane, M.F.; Guerrette, S.; Marsischky, G.T.; Kolodner, R.; Fishel, R. HMSH2 Forms Specific Mispair-Binding Complexes with HMSH3 and HMSH6. Proc. Natl. Acad. Sci. USA 1996, 93, 13629–13634. [Google Scholar] [CrossRef] [Green Version]
- Genschel, J.; Littman, S.J.; Drummond, J.T.; Modrich, P. Isolation of MutSβ from Human Cells and Comparison of the Mismatch Repair Specificities of MutSβ and MutSα. J. Biol. Chem. 1998, 273, 19895–19901. [Google Scholar] [CrossRef] [Green Version]
- Marti, T.M.; Kunz, C.; Fleck, O. DNA Mismatch Repair and Mutation Avoidance Pathways. J. Cell. Physiol. 2002, 191, 28–41. [Google Scholar] [CrossRef]
- Modrich, P. Mechanisms and Biological Effects of Mismatch Repair. Annu. Rev. Genet. 1991, 25, 229–253. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, A.H.; Milligan, A.S.; Langridge, P.; Able, J.A. TaMSH7: A Cereal Mismatch Repair Gene That Affects Fertility in Transgenic Barley (Hordeum vulgare L.). BMC Plant Biol. 2007, 7, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, N.; Chambers, S.R.; Louis, E.J.; Borts, R.H. The mismatch repair system contributes to meiotic sterility in an interspecific yeast hybrid. EMBO J. 1996, 15, 1726–1733. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.M.; Bollmann, S.R.; Hays, J.B. Reduction of Stability of Arabidopsis Genomic and Transgenic DNA-Repeat Sequences (Microsatellites) by Inactivation of AtMSH2 Mismatch-Repair Function. Plant Physiol. 2003, 133, 328–338. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emmanuel, E.; Yehuda, E.; Melamed-Bessudo, C.; Avivi-Ragolsky, N.; Levy, A.A. The Role of AtMSH2 in Homologous Recombination in Arabidopsis thaliana. EMBO Rep. 2006, 7, 100–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karthika, V.; Chandrashekar, B.K.; Kiranmai, K.; Ag, S.; Makarla, U.; Ramu, V.S. Disruption in the DNA Mismatch Repair Gene MSH2 by CRISPR-Cas9 in Indica Rice Can Create Genetic Variability. J. Agric. Food Chem. 2021, 69, 4144–4152. [Google Scholar] [CrossRef]
- Wilkinson, J.E.; Twell, D.; Lindsey, K. Activities of CaMV 35S and Nos Promoters in Pollen: Implications for Field Release of Transgenic Plants. J. Exp. Bot. 1997, 48, 265–275. [Google Scholar] [CrossRef] [Green Version]
- Sunilkumar, G.; Mohr, L.; Lopata-Finch, E.; Emani, C.; Rathore, K.S. Developmental and Tissue-Specific Expression of CaMV 35S Promoter in Cotton as Revealed by GFP. Plant Mol. Biol. 2002, 50, 463–479. [Google Scholar] [CrossRef]
- Krinitsina, A.; Komakhin, R. Cloning of the Mismatch Repair MSH2 Gene Fragment from Various Tomato Species. Biomics 2019, 10, 336–343. [Google Scholar] [CrossRef]
- Madzharova, N.V.; Kazakova, K.A.; Strelnikova, S.R.; Snycheva, O.A.; Vetchinkina, E.M.; Efremova, L.N.; Vysotskii, D.A.; Babakov, A.V.; Komakhin, R.A. Promoters Pro-SmAMP1 and pro-SmAMP2 from Wild Plant Stellaria media for the Biotechnology of Dicotyledons. Russ. J. Plant Physiol. 2018, 65, 750–761. [Google Scholar] [CrossRef]
- Efremova, L.N.; Strelnikova, S.R.; Gazizova, G.R.; Minkina, E.A.; Komakhin, R.A. A Synthetic Strong and Constitutive Promoter Derived from the Stellaria media Pro-SmAMP1 and pro-SmAMP2 Promoters for Effective Transgene Expression in Plants. Genes 2020, 11, 1407. [Google Scholar] [CrossRef] [PubMed]
- Komakhin, R.A.; Vysotskii, D.A.; Shukurov, R.R.; Voblikova, V.D.; Komakhina, V.V.; Strelnikova, S.R.; Vetchinkina, E.M.; Babakov, A.V. Novel Strong Promoter of Antimicrobial Peptides Gene Pro-SmAMP2 from Chickweed (Stellaria media). BMC Biotechnol. 2016, 16, 43. [Google Scholar] [CrossRef] [Green Version]
- Komakhin, R.A.; Strelnikova, S.R.; Zhuchenko, A.A. Genetic Features of the Tomato Marker Line Mo938. Russ. J. Genet. 2019, 55, 52–60. [Google Scholar] [CrossRef]
- Komakhina, V.V.; Krinitsina, A.A.; Milyukova, N.A.; Komakhin, R.A. Expression of Recombinant SPO11 Genes Locally Alters Crossing over in Tomato. Russ. J. Genet. 2020, 56, 1079–1089. [Google Scholar] [CrossRef]
- Komakhin, R.A.; Abdeeva, I.A.; Salehi Dzhuzani, G.R.; Goldenkova, I.V.; Zhuchenko, A.A. Thermostable Lichenase as a Translational Reporter. Russ. J. Genet. 2005, 41, 23–31. [Google Scholar] [CrossRef]
- Komakhin, R.A.; Komakhina, V.V.; Milyukova, N.A.; Goldenkova-Pavlova, I.V.; Fadina, O.A.; Zhuchenko, A.A. Transgenic Tomato Plants Expressing RecA and NLS-RecA-LicBM3 Genes as a Model for Studying Meiotic Recombination. Russ. J. Genet. 2010, 46, 1440–1448. [Google Scholar] [CrossRef]
- Muller, P.Y.; Miserez, A.R.; Dobbie, Z. Processing of Gene Expression Data Generated by Quantitative Real-Time RT-PCR. BioTechniques 2002, 32, 6. [Google Scholar]
- Suzuki, D.T.; Griffiths, A.J.F. Introduction to Genetic Analysis; W. H. Freeman: New York, NY, USA, 1976. [Google Scholar]
- Kosambi, D.D. The Estimation of Map Distances from Recombination Values. In Selected Works in Mathematics and Statistics; Ramaswamy, R., Ed.; Springer: New Delhi, India, 2016; pp. 125–130. ISBN 978-81-322-3676-4. [Google Scholar]
- Chetelat, R.T. Overcoming Sterility and Unilateral Incompatibility of Solanum lycopersicum × S. sitiens Hybrids. Euphytica 2016, 207, 319–330. [Google Scholar] [CrossRef]
- Hoffman, P.D.; Leonard, J.M.; Lindberg, G.E.; Bollmann, S.R.; Hays, J.B. Rapid Accumulation of Mutations during Seed-to-Seed Propagation of Mismatch-Repair-Defective Arabidopsis. Genes Dev. 2004, 18, 2676–2685. [Google Scholar] [CrossRef] [Green Version]
- Rakosy-Tican, E.; Lörincz-Besenyei, E.; Molnár, I.; Thieme, R.; Hartung, F.; Sprink, T.; Antonova, O.; Famelaer, I.; Angenon, G.; Aurori, A. New Phenotypes of Potato Co-Induced by Mismatch Repair Deficiency and Somatic Hybridization. Front. Plant Sci. 2019, 10, 3. [Google Scholar] [CrossRef] [Green Version]
- Sarma, S.; Pandey, A.K.; Sharma, K.; Ravi, M.; Sreelakshmi, Y.; Sharma, R. MutS-Homolog2 Silencing Generates Tetraploid Meiocytes in Tomato (Solanum lycopersicum). Plant Direct 2018, 2, e00017. [Google Scholar] [CrossRef] [Green Version]
- Serra, H.; Svačina, R.; Baumann, U.; Whitford, R.; Sutton, T.; Bartoš, J.; Sourdille, P. Ph2 encodes the mismatch repair protein MSH7-3D that inhibits wheat homoeologous recombination. Nat. Commun. 2021, 12, 803. [Google Scholar] [CrossRef] [PubMed]
- Frohnmeyer, H.; Hahlbrock, K.; Schafer, E. A Light-Responsive in Vitro Transcription System from Evacuolated Parsley Protoplasts. Plant J. 1994, 5, 437–449. [Google Scholar] [CrossRef]
- Cooker, D.E.; Webb, K.J. Stability of CaMV 35S-Gus Gene Expression in (Bird’s Foot Trefoil) Hairy Root Cultures under Different Growth Conditions. Plant Cell Tissue Organ Cult. 1997, 47, 163–168. [Google Scholar] [CrossRef]
- Yamamoto, Y.Y.; Kondo, Y.; Kato, A.; Tsuji, H.; Obokata, J. Light-Responsive Elements of the Tobacco PSI-D Gene Are Located Both Upstream and within the Transcribed Region. Plant J. 1997, 12, 255–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kurata, H.; Takemura, T.; Furusaki, S.; Kado, C.I. Light-Controlled Expression of a Foreign Gene Using the Chalcone Synthase Promoter in Tobacco BY-2 Cells. J. Ferment. Bioeng. 1998, 86, 317–323. [Google Scholar] [CrossRef]
- Zambre, M.; Terryn, N.; De Clercq, J.; De Buck, S.; Dillen, W.; Van Montagu, M.; Van Der Straeten, D.; Angenon, G. Light Strongly Promotes Gene Transfer from Agrobacterium tumefaciens to Plant Cells. Planta 2003, 216, 580–586. [Google Scholar] [CrossRef]
- Ellis, D.D.; McCabe, D.; Russell, D.; Martinell, B.; McCown, B.H. Expression of Inducible Angiosperm Promoters in a Gymnosperm, Picea Glauca (White Spruce). Plant Mol. Biol. 1991, 17, 19–27. [Google Scholar] [CrossRef]
- Schnurr, J.A.; Guerra, D.J. The CaMV-35S Promoter Is Sensitive to Shortened Photoperiod in Transgenic Tobacco. Plant Cell Rep. 2000, 19, 279–282. [Google Scholar] [CrossRef] [PubMed]
- Zuker, A.; Ahroni, A.; Tzfira, T.; Ben-Meir, H.; Vainstein, A. Wounding by Bombardment Yields Highly Efficient Agrobacterium-Mediated Transformation of Carnation (Dianthus caryophyllus L.). Mol. Breed. 1999, 5, 367–375. [Google Scholar] [CrossRef]
- Bovy, A.; Van Den Berg, C.; De Vrieze, G.; Thompson, W.F.; Weisbeek, P.; Smeekens, S. Light-Regulated Expression of the Arabidopsis thaliana Ferredoxin Gene Requires Sequences Upstream and Downstream of the Transcription Initiation Site. Plant Mol. Biol. 1995, 27, 27–39. [Google Scholar] [CrossRef] [PubMed]
- Dickey, L.; Gallo-Meagher, M.; Thompson, W. Light Regulatory Sequences are Located within the 5′ Portion of the Fed-1 Message Sequence. EMBO J. 1992, 11, 2311–2317. [Google Scholar] [CrossRef] [PubMed]
- Okayama, T.; Furukawa, H.; Okamura, K.; Murase, H. The Effect of Photoperiod on β-Glucuronidase gene Expression under Control CaMV-35S Promoter in Transgenic Lettuce. Environ. Control. Biol. 2010, 48, 1–8. [Google Scholar] [CrossRef]
- Saidi, Y.; Schaefer, D.G.; Goloubinoff, P.; Zrÿd, J.-P.; Finka, A. The CaMV 35S Promoter Has a Weak Expression Activity in Dark Grown Tissues of Moss Physcomitrella patens. Plant Signal. Behav. 2009, 4, 457–459. [Google Scholar] [CrossRef] [Green Version]
- Ehlenfeldt, M.K.; Ortiz, R. Evidence on the Nature and Origins of Endosperm Dosage Requirements in Solanum and Other Angiosperm Genera. Sex. Plant Reprod. 1995, 8, 189–196. [Google Scholar] [CrossRef]
- Nilsson, E. Some experiments with tetraploid tomatoes. Hereditas 1950, 36, 181–204. [Google Scholar] [CrossRef]
- Ren, B.; Cam, H.; Takahashi, Y.; Volkert, T.; Terragni, J.; Young, R.A.; Dynlacht, B.D. E2F integrates cell cycle progression with DNA repair, replication, and G2/M checkpoints. Genes Dev. 2002, 16, 245–256. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Wang, H.; Zhuang, D.; Zhu, H.; Du, Y.; Cheng, Z.; Cui, W.; Rogers, H.J.; Zhang, Q.; Jia, C.; et al. Roles of MSH2 and MSH6 in cadmium-induced G2/M checkpoint arrest in Arabidopsis roots. Chemosphere 2018, 201, 586–594. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Sequence | Primer | 5′-3′ | Amplicon, bp |
|---|---|---|---|
| MSH2 | antM2plus | aagctttgatagccacttcaccaatt | 963 |
| antM2minus | ggatccgtttgtttgtgcaaattatg | ||
| sM2plus | aagcttagcaattatggaaactacacagtc | 662 | |
| sM2minus | tctagagtttgtttgtgcaaattatgaatc | ||
| TaqMan-msh | FAMttctcttggagaggatggatcgBHQ1 | - | |
| msh2 + 1 | tgcgctttccagtgttagtg | 109 * | |
| msh2-1 | tgaaccactgccctcataca | ||
| CaMV35S | E35plus | aacaaagggtaatatccggaaacc | - |
| pro-SmAMP2 | pSA2 | acggaattcgtaccaacgtgagtaactat | - |
| Actin (U60482) | TaqMan-act | [26] | 65 * |
| Fwd-act | |||
| Rev-act |
| F1 Hybrid | F2 Phenotypic Classes | χ2 (3:1) | rf, % | ||||
|---|---|---|---|---|---|---|---|
| WvD | wvD | Wvd | wvd | ||||
| Wv:wv | D:d | ||||||
| F1p35S-antM2 No. 3-1 | 260 | 43 | 43 | 64 | 0.26 | 0.26 | 23.3 ± 0.2 |
| F1p35S-antM2 No. 16-1 | 465 | 65 | 58 | 104 | 0.12 | 0.93 | 20.3 ± 2.6 |
| F1p35S-antM2 No. 111-2 | 261 | 40 | 35 | 58 | 0 | 0.41 | 22.5 ± 2.2 |
| F1p35S-antM2 No. 115-11 | 513 | 74 | 79 | 104 | 1.46 | 0.63 | 23.0 ± 0.9 |
| Mean F1p35S-antM2 | 1499 | 222 | 215 | 330 | 1.09 | 0.49 | 22.3 ± 0.7 |
| F1pSmAMP2-antM2 No. 4-2-4 | 536 | 85 | 93 | 114 | 0.41 | 0 | 24.6 ± 1.0 |
| F1 (Marglobe × Mo938) | 393 | 65 | 55 | 72 | 0.1 | 3.3 | 23.8 ± 1.8 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Strelnikova, S.R.; Krinitsina, A.A.; Komakhin, R.A. Effective RNAi-Mediated Silencing of the Mismatch Repair MSH2 Gene Induces Sterility of Tomato Plants but Not an Increase in Meiotic Recombination. Genes 2021, 12, 1167. https://doi.org/10.3390/genes12081167
Strelnikova SR, Krinitsina AA, Komakhin RA. Effective RNAi-Mediated Silencing of the Mismatch Repair MSH2 Gene Induces Sterility of Tomato Plants but Not an Increase in Meiotic Recombination. Genes. 2021; 12(8):1167. https://doi.org/10.3390/genes12081167
Chicago/Turabian StyleStrelnikova, Svetlana R., Anastasiya A. Krinitsina, and Roman A. Komakhin. 2021. "Effective RNAi-Mediated Silencing of the Mismatch Repair MSH2 Gene Induces Sterility of Tomato Plants but Not an Increase in Meiotic Recombination" Genes 12, no. 8: 1167. https://doi.org/10.3390/genes12081167
APA StyleStrelnikova, S. R., Krinitsina, A. A., & Komakhin, R. A. (2021). Effective RNAi-Mediated Silencing of the Mismatch Repair MSH2 Gene Induces Sterility of Tomato Plants but Not an Increase in Meiotic Recombination. Genes, 12(8), 1167. https://doi.org/10.3390/genes12081167