
Essential Role of the 14q32 Encoded miRNAs in Endocrine Tumors


Abstract
1. Introduction
2. Materials and Methods
3. miRNAs in Endocrine Tumors
3.1. Neuroendocrine Tumors (NET)
3.2. Pheochromocytoma-Paraganglioma (PPGL)
3.3. Adrenocortical Tumors
3.4. Pituitary Neuroendocrine Tumors (PitNET)
3.5. Thyroid Carcinoma
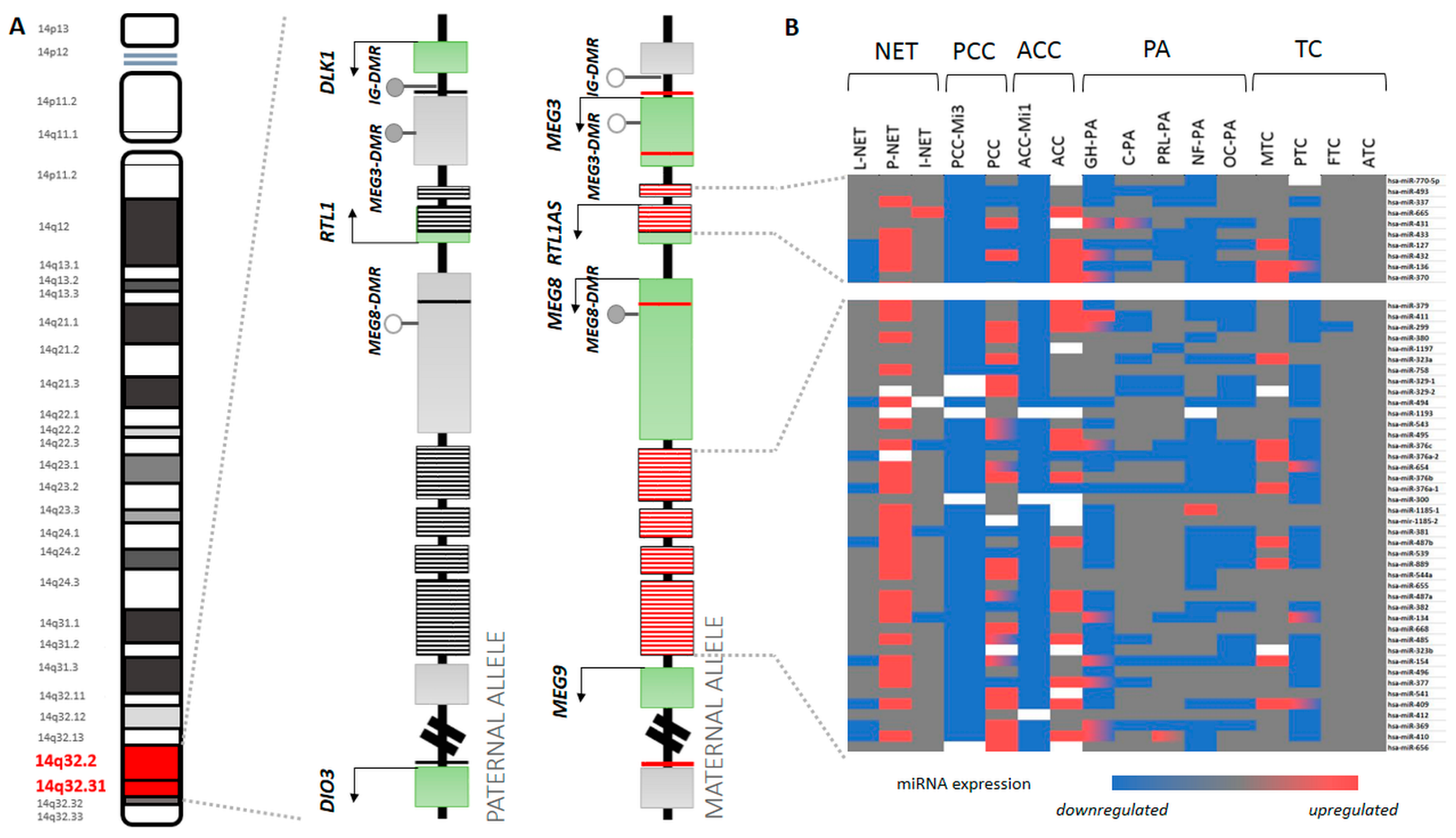
4. Different Expression of 14q32 miRNA Cluster Members
5. Functional Impact of 14q32 miRNAs
6. Summary and Discussion
7. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yates, L.A.; Norbury, C.J.; Gilbert, R.J.C. The Long and Short of MicroRNA. Cell 2013, 153, 516–519. [Google Scholar] [CrossRef]
- Valinezhad Orang, A.; Safaralizadeh, R.; Kazemzadeh-Bavili, M. Mechanisms of MiRNA-Mediated Gene Regulation from Common Downregulation to MRNA-Specific Upregulation. Int. J. Genom. 2014, 2014, 970607. [Google Scholar] [CrossRef]
- Lewis, B.P.; Burge, C.B.; Bartel, D.P. Conserved Seed Pairing, Often Flanked by Adenosines, Indicates That Thousands of Human Genes Are MicroRNA Targets. Cell 2005, 120, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Mattick, J.S.; Makunin, I.V. Small Regulatory RNAs in Mammals. Hum. Mol. Genet. 2005, 14 (Suppl. S1), R121–R132. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Wang, Y.-L. Editorial: MicroRNAs as New Players in Endocrinology. Front. Endocrinol. 2018, 9, 459. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Patócs, A. MicroRNAs in Endocrine Tumors. EJIFCC 2019, 30, 146–164. [Google Scholar] [PubMed]
- Seitz, H. A Large Imprinted MicroRNA Gene Cluster at the Mouse Dlk1-Gtl2 Domain. Genome Res. 2004, 14, 1741–1748. [Google Scholar] [CrossRef]
- Cavaillé, J.; Seitz, H.; Paulsen, M.; Ferguson-Smith, A.C.; Bachellerie, J.-P. Identification of Tandemly-Repeated C/D SnoRNA Genes at the Imprinted Human 14q32 Domain Reminiscent of Those at the Prader-Willi/Angelman Syndrome Region. Hum. Mol. Genet. 2002, 11, 1527–1538. [Google Scholar] [CrossRef]
- Royo, H.; Cavaillé, J. Non-Coding RNAs in Imprinted Gene Clusters. Biol. Cell 2008, 100, 149–166. [Google Scholar] [CrossRef]
- Goossens, E.A.C.; de Vries, M.R.; Simons, K.H.; Putter, H.; Quax, P.H.A.; Nossent, A.Y. MiRMap: Profiling 14q32 MicroRNA Expression and DNA Methylation throughout the Human Vasculature. Front. Cardiovasc. Med. 2019, 6. [Google Scholar] [CrossRef]
- Valbuena, G.N.; Apostolidou, S.; Roberts, R.; Barnes, J.; Alderton, W.; Harper, L.; Jacobs, I.; Menon, U.; Keun, H.C. The 14q32 Maternally Imprinted Locus Is a Major Source of Longitudinally Stable Circulating MicroRNAs as Measured by Small RNA Sequencing. Sci. Rep. 2019, 9, 15787. [Google Scholar] [CrossRef]
- Glazov, E.A.; McWilliam, S.; Barris, W.C.; Dalrymple, B.P. Origin, Evolution, and Biological Role of MiRNA Cluster in DLK-DIO3 Genomic Region in Placental Mammals. Mol. Biol. Evol. 2008, 25, 939–948. [Google Scholar] [CrossRef] [PubMed]
- Benetatos, L.; Voulgaris, E.; Vartholomatos, G. DLK1-MEG3 Imprinted Domain MicroRNAs in Cancer Biology. Crit. Rev. Eukaryot. Gene Expr. 2012, 22, 1–15. [Google Scholar] [CrossRef]
- Benetatos, L.; Vartholomatos, G.; Hatzimichael, E. MEG3 Imprinted Gene Contribution in Tumorigenesis. Int. J. Cancer 2011, 129, 773–779. [Google Scholar] [CrossRef] [PubMed]
- Wienholds, E. MicroRNA Expression in Zebrafish Embryonic Development. Science 2005, 309, 310–311. [Google Scholar] [CrossRef] [PubMed]
- Swarbrick, A.; Woods, S.L.; Shaw, A.; Balakrishnan, A.; Phua, Y.; Nguyen, A.; Chanthery, Y.; Lim, L.; Ashton, L.J.; Judson, R.L.; et al. MiR-380-5p Represses P53 to Control Cellular Survival and Is Associated with Poor Outcome in MYCN Amplified Neuroblastoma. Nat. Med. 2010, 16, 1134–1140. [Google Scholar] [CrossRef]
- Holder, J.L.; Lotze, T.E.; Bacino, C.; Cheung, S.-W. A Child with an Inherited 0.31 Mb Microdeletion of Chromosome 14q32.33: Further Delineation of a Critical Region for the 14q32 Deletion Syndrome. Am. J. Med. Genet. A 2012, 158A, 1962–1966. [Google Scholar] [CrossRef]
- Briggs, T.A.; Lokulo-Sodipe, K.; Chandler, K.E.; Mackay, D.J.G.; Temple, I.K. Temple Syndrome as a Result of Isolated Hypomethylation of the 14q32 Imprinted DLK1/MEG3 Region. Am. J. Med. Genet. A 2016, 170A, 170–175. [Google Scholar] [CrossRef]
- Bens, S.; Kolarova, J.; Gillessen-Kaesbach, G.; Buiting, K.; Beygo, J.; Caliebe, A.; Ammerpohl, O.; Siebert, R. The Differentially Methylated Region of MEG8 Is Hypermethylated in Patients with Temple Syndrome. Epigenomics 2015, 7, 1089–1097. [Google Scholar] [CrossRef]
- Kagami, M.; O’Sullivan, M.J.; Green, A.J.; Watabe, Y.; Arisaka, O.; Masawa, N.; Matsuoka, K.; Fukami, M.; Matsubara, K.; Kato, F.; et al. The IG-DMR and the MEG3-DMR at Human Chromosome 14q32.2: Hierarchical Interaction and Distinct Functional Properties as Imprinting Control Centers. PLoS Genet. 2010, 6, e1000992. [Google Scholar] [CrossRef] [PubMed]
- Kagami, M.; Sekita, Y.; Nishimura, G.; Irie, M.; Kato, F.; Okada, M.; Yamamori, S.; Kishimoto, H.; Nakayama, M.; Tanaka, Y.; et al. Deletions and Epimutations Affecting the Human 14q32.2 Imprinted Region in Individuals with Paternal and Maternal Upd(14)-like Phenotypes. Nat. Genet. 2008, 40, 237–242. [Google Scholar] [CrossRef]
- Uppal, A.; Wightman, S.C.; Mallon, S.; Oshima, G.; Pitroda, S.P.; Zhang, Q.; Huang, X.; Darga, T.E.; Huang, L.; Andrade, J.; et al. 14q32-Encoded MicroRNAs Mediate an Oligometastatic Phenotype. Oncotarget 2015, 6, 3540–3552. [Google Scholar] [CrossRef]
- Rosenfeld, J.A.; Fox, J.E.; Descartes, M.; Brewer, F.; Stroud, T.; Gorski, J.L.; Upton, S.J.; Moeschler, J.B.; Monteleone, B.; Neill, N.J.; et al. Clinical Features Associated with Copy Number Variations of the 14q32 Imprinted Gene Cluster. Am. J. Med. Genet. Part A 2015, 167, 345–353. [Google Scholar] [CrossRef] [PubMed]
- Ogata, T.; Kagami, M. Kagami–Ogata Syndrome: A Clinically Recognizable Upd(14)Pat and Related Disorder Affecting the Chromosome 14q32.2 Imprinted Region. J. Hum. Genet. 2016, 61, 87–94. [Google Scholar] [CrossRef]
- Sun, M.; Xia, R.; Jin, F.; Xu, T.; Liu, Z.; De, W.; Liu, X. Downregulated Long Noncoding RNA MEG3 Is Associated with Poor Prognosis and Promotes Cell Proliferation in Gastric Cancer. Tumour Biol. 2014, 35, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Béna, F.; Gimelli, S.; Migliavacca, E.; Brun-Druc, N.; Buiting, K.; Antonarakis, S.E.; Sharp, A.J. A Recurrent 14q32.2 Microdeletion Mediated by Expanded TGG Repeats. Hum. Mol. Genet. 2010, 19, 1967–1973. [Google Scholar] [CrossRef] [PubMed]
- Kawakami, T.; Chano, T.; Minami, K.; Okabe, H.; Okada, Y.; Okamoto, K. Imprinted DLK1 Is a Putative Tumor Suppressor Gene and Inactivated by Epimutation at the Region Upstream of GTL2 in Human Renal Cell Carcinoma. Hum. Mol. Genet. 2006, 15, 821–830. [Google Scholar] [CrossRef]
- Sanlaville, D.; Aubry, M.C.; Dumez, Y.; Nolen, M.C.; Amiel, J.; Pinson, M.P.; Lyonnet, S.; Munnich, A.; Vekemans, M.; Morichon-Delvallez, N. Maternal Uniparental Heterodisomy of Chromosome 14: Chromosomal Mechanism and Clinical Follow Up. J. Med. Genet. 2000, 37, 525–528. [Google Scholar] [CrossRef]
- Temple, I.K.; Shrubb, V.; Lever, M.; Bullman, H.; Mackay, D.J.G. Isolated Imprinting Mutation of the DLK1/GTL2 Locus Associated with a Clinical Presentation of Maternal Uniparental Disomy of Chromosome 14. J. Med. Genet. 2007, 44, 637–640. [Google Scholar] [CrossRef]
- Da Rocha, S.T.; Edwards, C.A.; Ito, M.; Ogata, T.; Ferguson-Smith, A.C. Genomic Imprinting at the Mammalian Dlk1-Dio3 Domain. Trends Genet. 2008, 24, 306–316. [Google Scholar] [CrossRef] [PubMed]
- Hagan, J.P.; O’Neill, B.L.; Stewart, C.L.; Kozlov, S.V.; Croce, C.M. At Least Ten Genes Define the Imprinted Dlk1-Dio3 Cluster on Mouse Chromosome 12qF1. PLoS ONE 2009, 4, e4352. [Google Scholar] [CrossRef] [PubMed]
- Wylie, A.A.; Murphy, S.K.; Orton, T.C.; Jirtle, R.L. Novel Imprinted DLK1/GTL2 Domain on Human Chromosome 14 Contains Motifs That Mimic Those Implicated in IGF2/H19 Regulation. Genome Res. 2000, 10, 1711–1718. [Google Scholar] [CrossRef] [PubMed]
- Takada, S.; Paulsen, M.; Tevendale, M.; Tsai, C.-E.; Kelsey, G.; Cattanach, B.M.; Ferguson-Smith, A.C. Epigenetic Analysis of the Dlk1-Gtl2 Imprinted Domain on Mouse Chromosome 12: Implications for Imprinting Control from Comparison with Igf2-H19. Hum. Mol. Genet. 2002, 11, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, J.V.; Matteson, P.G.; Jones, B.K.; Guan, X.J.; Tilghman, S.M. The Dlk1 and Gtl2 Genes Are Linked and Reciprocally Imprinted. Genes Dev. 2000, 14, 1997–2002. [Google Scholar] [PubMed]
- Liu, L.; Luo, G.-Z.; Yang, W.; Zhao, X.; Zheng, Q.; Lv, Z.; Li, W.; Wu, H.-J.; Wang, L.; Wang, X.-J.; et al. Activation of the Imprinted Dlk1-Dio3 Region Correlates with Pluripotency Levels of Mouse Stem Cells. J. Biol. Chem. 2010, 285, 19483–19490. [Google Scholar] [CrossRef] [PubMed]
- Kameswaran, V.; Bramswig, N.C.; McKenna, L.B.; Penn, M.; Schug, J.; Hand, N.J.; Chen, Y.; Choi, I.; Vourekas, A.; Won, K.-J.; et al. Epigenetic Regulation of the DLK1-MEG3 MicroRNA Cluster in Human Type 2 Diabetic Islets. Cell Metab. 2014, 19, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Guo, W.; Dong, Z.; Liu, S.; Qiao, Y.; Kuang, G.; Guo, Y.; Shen, S.; Liang, J. Promoter Hypermethylation-Mediated Downregulation of MiR-770 and Its Host Gene MEG3, a Long Non-Coding RNA, in the Development of Gastric Cardia Adenocarcinoma. Mol. Carcinog. 2017, 56, 1924–1934. [Google Scholar] [CrossRef]
- González-Vallinas, M.; Rodríguez-Paredes, M.; Albrecht, M.; Sticht, C.; Stichel, D.; Gutekunst, J.; Pitea, A.; Sass, S.; Sánchez-Rivera, F.J.; Lorenzo-Bermejo, J.; et al. Epigenetically Regulated Chromosome 14q32 MiRNA Cluster Induces Metastasis and Predicts Poor Prognosis in Lung Adenocarcinoma Patients. Mol. Cancer Res. 2018, 16, 390–402. [Google Scholar] [CrossRef]
- Moradi, S.; Sharifi-Zarchi, A.; Ahmadi, A.; Mollamohammadi, S.; Stubenvoll, A.; Günther, S.; Salekdeh, G.H.; Asgari, S.; Braun, T.; Baharvand, H. Small RNA Sequencing Reveals Dlk1-Dio3 Locus-Embedded MicroRNAs as Major Drivers of Ground-State Pluripotency. Stem Cell Rep. 2017, 9, 2081–2096. [Google Scholar] [CrossRef]
- Xi, S.; Xu, H.; Shan, J.; Tao, Y.; Hong, J.A.; Inchauste, S.; Zhang, M.; Kunst, T.F.; Mercedes, L.; Schrump, D.S. Cigarette Smoke Mediates Epigenetic Repression of MiR-487b during Pulmonary Carcinogenesis. J. Clin. Investig. 2013, 123, 1241–1261. [Google Scholar] [CrossRef]
- Oshima, G.; Poli, E.C.; Bolt, M.J.; Chlenski, A.; Forde, M.; Jutzy, J.M.S.; Biyani, N.; Posner, M.C.; Pitroda, S.P.; Weichselbaum, R.R.; et al. DNA Methylation Controls Metastasis-Suppressive 14q32-Encoded MiRNAs. Cancer Res. 2019, 79, 650–662. [Google Scholar] [CrossRef]
- Kelly, L.; Bryan, K.; Kim, S.Y.; Janeway, K.A.; Killian, J.K.; Schildhaus, H.-U.; Miettinen, M.; Helman, L.; Meltzer, P.S.; van de Rijn, M.; et al. Post-Transcriptional Dysregulation by MiRNAs Is Implicated in the Pathogenesis of Gastrointestinal Stromal Tumor [GIST]. PLoS ONE 2013, 8, e64102. [Google Scholar] [CrossRef]
- Anaya-Ruiz, M.; Bandala, C.; Perez-Santos, J.L.M. MiR-485 Acts as a Tumor Suppressor by Inhibiting Cell Growth and Migration in Breast Carcinoma T47D Cells. Asian Pac. J. Cancer Prev. 2013, 14, 3757–3760. [Google Scholar] [CrossRef]
- Qin, R.; Chen, Z.; Ding, Y.; Hao, J.; Hu, J.; Guo, F. Long Non-Coding RNA MEG3 Inhibits the Proliferation of Cervical Carcinoma Cells through the Induction of Cell Cycle Arrest and Apoptosis. Neoplasma 2013, 60, 486–492. [Google Scholar] [CrossRef]
- Ying, L.; Huang, Y.; Chen, H.; Wang, Y.; Xia, L.; Chen, Y.; Liu, Y.; Qiu, F. Downregulated MEG3 Activates Autophagy and Increases Cell Proliferation in Bladder Cancer. Mol. Biosyst. 2013, 9, 407–411. [Google Scholar] [CrossRef]
- Formosa, A.; Markert, E.K.; Lena, A.M.; Italiano, D.; Finazzi-Agrò, E.; Levine, A.J.; Bernardini, S.; Garabadgiu, A.V.; Melino, G.; Candi, E. MicroRNAs, MiR-154, MiR-299-5p, MiR-376a, MiR-376c, MiR-377, MiR-381, MiR-487b, MiR-485-3p, MiR-495 and MiR-654-3p, Mapped to the 14q32.31 Locus, Regulate Proliferation, Apoptosis, Migration and Invasion in Metastatic Prostate Cancer Cells. Oncogene 2014, 33, 5173–5182. [Google Scholar] [CrossRef]
- Sarver, A.L.; Thayanithy, V.; Scott, M.C.; Cleton-Jansen, A.-M.; Hogendoorn, P.C.; Modiano, J.F.; Subramanian, S. MicroRNAs at the Human 14q32 Locus Have Prognostic Significance in Osteosarcoma. Orphanet J. Rare Dis. 2013, 8, 7. [Google Scholar] [CrossRef] [PubMed]
- Zehavi, L.; Avraham, R.; Barzilai, A.; Bar-Ilan, D.; Navon, R.; Sidi, Y.; Avni, D.; Leibowitz-Amit, R. Silencing of a Large MicroRNA Cluster on Human Chromosome 14q32 in Melanoma: Biological Effects of Mir-376a and Mir-376c on Insulin Growth Factor 1 Receptor. Mol. Cancer 2012, 11, 44. [Google Scholar] [CrossRef] [PubMed]
- Niu, C.S.; Yang, Y.; Cheng, C.-D. MiR-134 Regulates the Proliferation and Invasion of Glioblastoma Cells by Reducing Nanog Expression. Int. J. Oncol. 2013, 42, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Zhang, J.; Feng, Y.; Li, R.; Sun, X.; Du, W.; Piao, X.; Wang, H.; Yang, D.; Sun, Y.; et al. MiR-410 Regulates MET to Influence the Proliferation and Invasion of Glioma. Int. J. Biochem. Cell Biol. 2012, 44, 1711–1717. [Google Scholar] [CrossRef] [PubMed]
- Lim, L.; Balakrishnan, A.; Huskey, N.; Jones, K.D.; Jodari, M.; Ng, R.; Song, G.; Riordan, J.; Anderton, B.; Cheung, S.-T.; et al. MicroRNA-494 within an Oncogenic MicroRNA Megacluster Regulates G1/S Transition in Liver Tumorigenesis through Suppression of Mutated in Colorectal Cancer. Hepatology 2014, 59, 202–215. [Google Scholar] [CrossRef]
- Chien, W.W.; Domenech, C.; Catallo, R.; Kaddar, T.; Magaud, J.-P.; Salles, G.; Ffrench, M. Cyclin-Dependent Kinase 1 Expression Is Inhibited by P16 INK4a at the Post-Transcriptional Level through the MicroRNA Pathway. Oncogene 2011, 30, 1880–1891. [Google Scholar] [CrossRef][Green Version]
- Dixon-McIver, A.; East, P.; Mein, C.A.; Cazier, J.-B.; Molloy, G.; Chaplin, T.; Andrew Lister, T.; Young, B.D.; Debernardi, S. Distinctive Patterns of MicroRNA Expression Associated with Karyotype in Acute Myeloid Leukaemia. PLoS ONE 2008, 3, e2141. [Google Scholar] [CrossRef] [PubMed]
- Chabre, O.; Libé, R.; Assie, G.; Barreau, O.; Bertherat, J.; Bertagna, X.; Feige, J.-J.; Cherradi, N. Serum MiR-483-5p and MiR-195 Are Predictive of Recurrence Risk in Adrenocortical Cancer Patients. Endocr. Relat. Cancer 2013, 20, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Lussier, Y.A.; Xing, H.R.; Salama, J.K.; Khodarev, N.N.; Huang, Y.; Zhang, Q.; Khan, S.A.; Yang, X.; Hasselle, M.D.; Darga, T.E.; et al. MicroRNA Expression Characterizes Oligometastasis(Es). PLoS ONE 2011, 6, e28650. [Google Scholar] [CrossRef] [PubMed]
- Lussier, Y.A.; Khodarev, N.N.; Regan, K.; Corbin, K.; Li, H.; Ganai, S.; Khan, S.A.; Gnerlich, J.L.; Gnerlich, J.; Darga, T.E.; et al. Oligo- and Polymetastatic Progression in Lung Metastasis(Es) Patients Is Associated with Specific MicroRNAs. PLoS ONE 2012, 7, e50141. [Google Scholar] [CrossRef]
- Yoshimoto, T.; Motoi, N.; Yamamoto, N.; Nagano, H.; Ushijima, M.; Matsuura, M.; Okumura, S.; Yamaguchi, T.; Fukayama, M.; Ishikawa, Y. Pulmonary Carcinoids and Low-Grade Gastrointestinal Neuroendocrine Tumors Show Common MicroRNA Expression Profiles, Different from Adenocarcinomas and Small Cell Carcinomas. Neuroendocrinology 2018, 106, 47–57. [Google Scholar] [CrossRef] [PubMed]
- Mairinger, F.D.; Ting, S.; Werner, R.; Walter, R.F.H.; Hager, T.; Vollbrecht, C.; Christoph, D.; Worm, K.; Mairinger, T.; Sheu-Grabellus, S.-Y.; et al. Different Micro-RNA Expression Profiles Distinguish Subtypes of Neuroendocrine Tumors of the Lung: Results of a Profiling Study. Mod. Pathol. 2014, 27, 1632–1640. [Google Scholar] [CrossRef]
- Deng, B.; Molina, J.; Aubry, M.C.; Sun, Z.; Wang, L.; Eckloff, B.W.; Vasmatzis, G.; You, M.; Wieben, E.D.; Jen, J.; et al. Clinical Biomarkers of Pulmonary Carcinoid Tumors in Never Smokers via Profiling MiRNA and Target MRNA. Cell Biosci. 2014, 4, 35. [Google Scholar] [CrossRef] [PubMed]
- Rapa, I.; Votta, A.; Felice, B.; Righi, L.; Giorcelli, J.; Scarpa, A.; Speel, E.-J.M.; Scagliotti, G.V.; Papotti, M.; Volante, M. Identification of MicroRNAs Differentially Expressed in Lung Carcinoid Subtypes and Progression. Neuroendocrinology 2015, 101, 246–255. [Google Scholar] [CrossRef]
- Wong, J.J.M.; Ginter, P.S.; Tyryshkin, K.; Yang, X.; Nanayakkara, J.; Zhou, Z.; Tuschl, T.; Chen, Y.-T.; Renwick, N. Classifying Lung Neuroendocrine Neoplasms through MicroRNA Sequence Data Mining. Cancers 2020, 12, 2653. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, N.; Knief, J.; Kacprowski, T.; Lazar-Karsten, P.; Keck, T.; Billmann, F.; Schmid, S.; Luley, K.; Lehnert, H.; Brabant, G.; et al. MicroRNA Analysis of Gastroenteropancreatic Neuroendocrine Tumors and Metastases. Oncotarget 2018, 9, 28379–28390. [Google Scholar] [CrossRef] [PubMed]
- Roldo, C.; Missiaglia, E.; Hagan, J.P.; Falconi, M.; Capelli, P.; Bersani, S.; Calin, G.A.; Volinia, S.; Liu, C.-G.; Scarpa, A.; et al. MicroRNA Expression Abnormalities in Pancreatic Endocrine and Acinar Tumors Are Associated with Distinctive Pathologic Features and Clinical Behavior. J. Clin. Oncol. 2006, 24, 4677–4684. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Shan, A.; Su, Y.; Cheng, Y.; Gu, W.; Wang, W.; Ning, G.; Cao, Y. MiR-144/451 Promote Cell Proliferation via Targeting PTEN/AKT Pathway in Insulinomas. Endocrinology 2015, 156, 2429–2439. [Google Scholar] [CrossRef]
- Zhou, H.-Q.; Chen, Q.-C.; Qiu, Z.-T.; Tan, W.-L.; Mo, C.-Q.; Gao, S.-W. Integrative MicroRNA-MRNA and Protein-Protein Interaction Analysis in Pancreatic Neuroendocrine Tumors. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 2842–2852. [Google Scholar] [PubMed]
- Lee, Y.S.; Kim, H.; Kim, H.W.; Lee, J.-C.; Paik, K.-H.; Kang, J.; Kim, J.; Yoon, Y.-S.; Han, H.-S.; Sohn, I.; et al. High Expression of MicroRNA-196a Indicates Poor Prognosis in Resected Pancreatic Neuroendocrine Tumor. Medicine 2015, 94, e2224. [Google Scholar] [CrossRef]
- Arvidsson, Y.; Rehammar, A.; Bergström, A.; Andersson, E.; Altiparmak, G.; Swärd, C.; Wängberg, B.; Kristiansson, E.; Nilsson, O. MiRNA Profiling of Small Intestinal Neuroendocrine Tumors Defines Novel Molecular Subtypes and Identifies MiR-375 as a Biomarker of Patient Survival. Mod. Pathol. 2018, 31, 1302–1317. [Google Scholar] [CrossRef]
- Li, S.-C.; Essaghir, A.; Martijn, C.; Lloyd, R.V.; Demoulin, J.-B.; Oberg, K.; Giandomenico, V. Global MicroRNA Profiling of Well-Differentiated Small Intestinal Neuroendocrine Tumors. Mod. Pathol. 2013, 26, 685–696. [Google Scholar] [CrossRef]
- Castro-Vega, L.J.; Letouzé, E.; Burnichon, N.; Buffet, A.; Disderot, P.-H.; Khalifa, E.; Loriot, C.; Elarouci, N.; Morin, A.; Menara, M.; et al. Multi-Omics Analysis Defines Core Genomic Alterations in Pheochromocytomas and Paragangliomas. Nat. Commun. 2015, 6, 6044. [Google Scholar] [CrossRef]
- Tömböl, Z.; Eder, K.; Kovács, A.; Szabó, P.M.; Kulka, J.; Likó, I.; Zalatnai, A.; Rácz, G.; Tóth, M.; Patócs, A.; et al. MicroRNA Expression Profiling in Benign (Sporadic and Hereditary) and Recurring Adrenal Pheochromocytomas. Mod. Pathol. 2010, 23, 1583–1595. [Google Scholar] [CrossRef] [PubMed]
- De Cubas, A.A.; Leandro-García, L.J.; Schiavi, F.; Mancikova, V.; Comino-Méndez, I.; Inglada-Pérez, L.; Perez-Martinez, M.; Ibarz, N.; Ximénez-Embún, P.; López-Jiménez, E.; et al. Integrative Analysis of MiRNA and MRNA Expression Profiles in Pheochromocytoma and Paraganglioma Identifies Genotype-Specific Markers and Potentially Regulated Pathways. Endocr. Relat. Cancer 2013, 20, 477–493. [Google Scholar] [CrossRef]
- Meyer-Rochow, G.Y.; Jackson, N.E.; Conaglen, J.V.; Whittle, D.E.; Kunnimalaiyaan, M.; Chen, H.; Westin, G.; Sandgren, J.; Stålberg, P.; Khanafshar, E.; et al. MicroRNA Profiling of Benign and Malignant Pheochromocytomas Identifies Novel Diagnostic and Therapeutic Targets. Endocr. Relat. Cancer 2010, 17, 835–846. [Google Scholar] [CrossRef]
- Calsina, B.; Castro-Vega, L.J.; Torres-Pérez, R.; Inglada-Pérez, L.; Currás-Freixes, M.; Roldán-Romero, J.M.; Mancikova, V.; Letón, R.; Remacha, L.; Santos, M.; et al. Integrative Multi-Omics Analysis Identifies a Prognostic MiRNA Signature and a Targetable MiR-21-3p/TSC2/MTOR Axis in Metastatic Pheochromocytoma/Paraganglioma. Theranostics 2019, 9, 4946–4958. [Google Scholar] [CrossRef] [PubMed]
- Tömböl, Z.; Szabó, P.M.; Molnár, V.; Wiener, Z.; Tölgyesi, G.; Horányi, J.; Riesz, P.; Reismann, P.; Patócs, A.; Likó, I.; et al. Integrative Molecular Bioinformatics Study of Human Adrenocortical Tumors: MicroRNA, Tissue-Specific Target Prediction, and Pathway Analysis. Endocr. Relat. Cancer 2009, 16, 895–906. [Google Scholar] [CrossRef] [PubMed]
- Özata, D.M.; Caramuta, S.; Velázquez-Fernández, D.; Akçakaya, P.; Xie, H.; Höög, A.; Zedenius, J.; Bäckdahl, M.; Larsson, C.; Lui, W.-O. The Role of MicroRNA Deregulation in the Pathogenesis of Adrenocortical Carcinoma. Endocr. Relat. Cancer 2011, 18, 643–655. [Google Scholar] [CrossRef]
- Assié, G.; Letouzé, E.; Fassnacht, M.; Jouinot, A.; Luscap, W.; Barreau, O.; Omeiri, H.; Rodriguez, S.; Perlemoine, K.; René-Corail, F.; et al. Integrated Genomic Characterization of Adrenocortical Carcinoma. Nat. Genet. 2014, 46, 607–612. [Google Scholar] [CrossRef] [PubMed]
- Mao, Z.-G.; He, D.-S.; Zhou, J.; Yao, B.; Xiao, W.-W.; Chen, C.-H.; Zhu, Y.-H.; Wang, H.-J. Differential Expression of MicroRNAs in GH-Secreting Pituitary Adenomas. Diagn. Pathol. 2010, 5, 79. [Google Scholar] [CrossRef]
- D’Angelo, D.; Palmieri, D.; Mussnich, P.; Roche, M.; Wierinckx, A.; Raverot, G.; Fedele, M.; Croce, C.M.; Trouillas, J.; Fusco, A. Altered MicroRNA Expression Profile in Human Pituitary GH Adenomas: Down-Regulation of MiRNA Targeting HMGA1, HMGA2, and E2F1. J. Clin. Endocrinol. Metab. 2012, 97, E1128–E1138. [Google Scholar] [CrossRef]
- Bottoni, A.; Zatelli, M.C.; Ferracin, M.; Tagliati, F.; Piccin, D.; Vignali, C.; Calin, G.A.; Negrini, M.; Croce, C.M.; Degli Uberti, E.C. Identification of Differentially Expressed MicroRNAs by Microarray: A Possible Role for MicroRNA Genes in Pituitary Adenomas. J. Cell. Physiol. 2007, 210, 370–377. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Liko, I.; Boyle, B.; Czirjak, S.; Igaz, P.; Patocs, A.; Racz, K. MicroRNA Expression in Human Sporadic Pituitary Adenomas. Endocr. Abstr. 2009, 20, 566. [Google Scholar]
- Cheunsuchon, P.; Zhou, Y.; Zhang, X.; Lee, H.; Chen, W.; Nakayama, Y.; Rice, K.A.; Tessa Hedley-Whyte, E.; Swearingen, B.; Klibanski, A. Silencing of the Imprinted DLK1-MEG3 Locus in Human Clinically Nonfunctioning Pituitary Adenomas. Am. J. Pathol. 2011, 179, 2120–2130. [Google Scholar] [CrossRef]
- He, Z.; Chen, L.; Hu, X.; Tang, J.; He, L.; Hu, J.; Fei, F.; Wang, Q. Next-Generation Sequencing of MicroRNAs Reveals a Unique Expression Pattern in Different Types of Pituitary Adenomas. Endocr. J. 2019, 66, 709–722. [Google Scholar] [CrossRef]
- Gentilin, E.; Tagliati, F.; Filieri, C.; Molè, D.; Minoia, M.; Rosaria Ambrosio, M.; Degli Uberti, E.C.; Zatelli, M.C. MiR-26a Plays an Important Role in Cell Cycle Regulation in ACTH-Secreting Pituitary Adenomas by Modulating Protein Kinase Cδ. Endocrinology 2013, 154, 1690–1700. [Google Scholar] [CrossRef]
- Amaral, F.C.; Torres, N.; Saggioro, F.; Neder, L.; Machado, H.R.; Silva, W.A.; Moreira, A.C.; Castro, M. MicroRNAs Differentially Expressed in ACTH-Secreting Pituitary Tumors. J. Clin. Endocrinol. Metab. 2009, 94, 320–323. [Google Scholar] [CrossRef] [PubMed]
- Stilling, G.; Sun, Z.; Zhang, S.; Jin, L.; Righi, A.; Kovācs, G.; Korbonits, M.; Scheithauer, B.W.; Kovacs, K.; Lloyd, R.V. MicroRNA Expression in ACTH-Producing Pituitary Tumors: Up-Regulation of MicroRNA-122 and -493 in Pituitary Carcinomas. Endocrine 2010, 38, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Müssnich, P.; Raverot, G.; Jaffrain-Rea, M.-L.; Fraggetta, F.; Wierinckx, A.; Trouillas, J.; Fusco, A.; D’Angelo, D. Downregulation of MiR-410 Targeting the Cyclin B1 Gene Plays a Role in Pituitary Gonadotroph Tumors. Cell Cycle 2015, 14, 2590–2597. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Li, Q.; Wang, C.; Su, Z.; Li, W.; Chen, X.; Wu, Z. Differential expression analysis of prolactinoma-related microRNAs. Zhonghua Yi Xue Za Zhi 2012, 92, 320–323. [Google Scholar] [PubMed]
- Darvasi, O.; Szabo, P.M.; Nemeth, K.; Szabo, K.; Spisak, S.; Liko, I.; Czirjak, S.; Racz, K.; Igaz, P.; Patocs, A.; et al. Limitations of High Throughput Methods for MiRNA Expression Profiles in Non-Functioning Pituitary Adenomas. Pathol. Oncol. Res. 2019, 25, 169–182. [Google Scholar] [CrossRef] [PubMed]
- Butz, H.; Likó, I.; Czirják, S.; Igaz, P.; Korbonits, M.; Rácz, K.; Patócs, A. MicroRNA Profile Indicates Downregulation of the TGFβ Pathway in Sporadic Non-Functioning Pituitary Adenomas. Pituitary 2011, 14, 112–124. [Google Scholar] [CrossRef]
- Liang, H.; Wang, R.; Diao, C.; Li, J.; Su, J.; Zhang, S. The PTTG1-Targeting MiRNAs MiR-329, MiR-300, MiR-381, and MiR-655 Inhibit Pituitary Tumor Cell Tumorigenesis and Are Involved in a P53/PTTG1 Regulation Feedback Loop. Oncotarget 2015, 6, 29413–29427. [Google Scholar] [CrossRef]
- Krokker, L.; Nyírő, G.; Reiniger, L.; Darvasi, O.; Szücs, N.; Czirják, S.; Tóth, M.; Igaz, P.; Patócs, A.; Butz, H. Differentially Expressed MiRNAs Influence Metabolic Processes in Pituitary Oncocytoma. Neurochem. Res. 2019. [Google Scholar] [CrossRef]
- Lassalle, S.; Zangari, J.; Popa, A.; Ilie, M.; Hofman, V.; Long, E.; Patey, M.; Tissier, F.; Belléannée, G.; Trouette, H.; et al. MicroRNA-375/SEC23A as Biomarkers of the in Vitro Efficacy of Vandetanib. Oncotarget 2016, 7, 30461–30478. [Google Scholar] [CrossRef] [PubMed]
- Hudson, J.; Duncavage, E.; Tamburrino, A.; Salerno, P.; Xi, L.; Raffeld, M.; Moley, J.; Chernock, R.D. Over Expression of MiR-10a and MiR-375 and down Regulation of YAP1 in Medullary Thyroid Carcinoma. Exp. Mol. Pathol. 2013, 95, 62–67. [Google Scholar] [CrossRef] [PubMed]
- Nikiforova, M.N.; Tseng, G.C.; Steward, D.; Diorio, D.; Nikiforov, Y.E. MicroRNA Expression Profiling of Thyroid Tumors: Biological Significance and Diagnostic Utility. J. Clin. Endocrinol. Metab. 2008, 93, 1600–1608. [Google Scholar] [CrossRef]
- Geraldo, M.V.; Nakaya, H.I.; Kimura, E.T. Down-Regulation of 14q32-Encoded MiRNAs and Tumor Suppressor Role for MiR-654-3p in Papillary Thyroid Cancer. Oncotarget 2017, 8, 9597–9607. [Google Scholar] [CrossRef]
- Rosignolo, F.; Sponziello, M.; Giacomelli, L.; Russo, D.; Pecce, V.; Biffoni, M.; Bellantone, R.; Lombardi, C.P.; Lamartina, L.; Grani, G.; et al. Identification of Thyroid-Associated Serum MicroRNA Profiles and Their Potential Use in Thyroid Cancer Follow-Up. J. Endocr. Soc. 2017, 1, 3–13. [Google Scholar] [CrossRef]
- Tetzlaff, M.T.; Liu, A.; Xu, X.; Master, S.R.; Baldwin, D.A.; Tobias, J.W.; Livolsi, V.A.; Baloch, Z.W. Differential Expression of MiRNAs in Papillary Thyroid Carcinoma Compared to Multinodular Goiter Using Formalin Fixed Paraffin Embedded Tissues. Endocr. Pathol. 2007, 18, 163–173. [Google Scholar] [CrossRef]
- Linwah, Y.; Kelly, L.; Yongli, S.; Armstrong, M.J.; Nikiforov, Y.E.; Carty, S.E.; Nikiforova, M.N. MicroRNA Signature Distinguishes the Degree of Aggressiveness of Papillary Thyroid Carcinoma. Ann. Surg. Oncol. 2011, 18, 2035–2041. [Google Scholar] [CrossRef]
- Jacques, C.; Guillotin, D.; Fontaine, J.-F.; Franc, B.; Mirebeau-Prunier, D.; Fleury, A.; Malthiery, Y.; Savagner, F. DNA Microarray and MiRNA Analyses Reinforce the Classification of Follicular Thyroid Tumors. J. Clin. Endocrinol. Metab. 2013, 98, E981–E989. [Google Scholar] [CrossRef] [PubMed]
- Lassalle, S.; Hofman, V.; Ilie, M.; Bonnetaud, C.; Puisségur, M.-P.; Brest, P.; Loubatier, C.; Guevara, N.; Bordone, O.; Cardinaud, B.; et al. Can the MicroRNA Signature Distinguish between Thyroid Tumors of Uncertain Malignant Potential and Other Well-Differentiated Tumors of the Thyroid Gland? Endocr. Relat. Cancer 2011, 18, 579–594. [Google Scholar] [CrossRef]
- Mancikova, V.; Castelblanco, E.; Pineiro-Yanez, E.; Perales-Paton, J.; de Cubas, A.A.; Inglada-Perez, L.; Matias-Guiu, X.; Capel, I.; Bella, M.; Lerma, E.; et al. MicroRNA Deep-Sequencing Reveals Master Regulators of Follicular and Papillary Thyroid Tumors. Mod. Pathol. 2015, 28, 748–757. [Google Scholar] [CrossRef]
- Peng, Y.; Li, C.; Luo, D.-C.; Ding, J.-W.; Zhang, W.; Pan, G. Expression Profile and Clinical Significance of MicroRNAs in Papillary Thyroid Carcinoma. Molecules 2014, 19, 11586–11599. [Google Scholar] [CrossRef] [PubMed]
- Riesco-Eizaguirre, G.; Wert-Lamas, L.; Perales-Patón, J.; Sastre-Perona, A.; Fernández, L.P.; Santisteban, P. The MiR-146b-3p/PAX8/NIS Regulatory Circuit Modulates the Differentiation Phenotype and Function of Thyroid Cells during Carcinogenesis. Cancer Res. 2015, 75, 4119–4130. [Google Scholar] [CrossRef]
- Saiselet, M.; Gacquer, D.; Spinette, A.; Craciun, L.; Decaussin-Petrucci, M.; Andry, G.; Detours, V.; Maenhaut, C. New Global Analysis of the MicroRNA Transcriptome of Primary Tumors and Lymph Node Metastases of Papillary Thyroid Cancer. BMC Genom. 2015, 16. [Google Scholar] [CrossRef] [PubMed]
- Swierniak, M.; Wojcicka, A.; Czetwertynska, M.; Stachlewska, E.; Maciag, M.; Wiechno, W.; Gornicka, B.; Bogdanska, M.; Koperski, L.; de la Chapelle, A.; et al. In-Depth Characterization of the MicroRNA Transcriptome in Normal Thyroid and Papillary Thyroid Carcinoma. J. Clin. Endocrinol. Metab. 2013, 98, E1401–E1409. [Google Scholar] [CrossRef] [PubMed]
- Rossing, M.; Borup, R.; Henao, R.; Winther, O.; Vikesaa, J.; Niazi, O.; Godballe, C.; Krogdahl, A.; Glud, M.; Hjort-Sørensen, C.; et al. Down-Regulation of MicroRNAs Controlling Tumourigenic Factors in Follicular Thyroid Carcinoma. J. Mol. Endocrinol. 2012, 48, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Dettmer, M.; Vogetseder, A.; Durso, M.B.; Moch, H.; Komminoth, P.; Perren, A.; Nikiforov, Y.E.; Nikiforova, M.N. MicroRNA Expression Array Identifies Novel Diagnostic Markers for Conventional and Oncocytic Follicular Thyroid Carcinomas. J. Clin. Endocrinol. Metab. 2013, 98, E1–E7. [Google Scholar] [CrossRef]
- Wojtas, B.; Ferraz, C.; Stokowy, T.; Hauptmann, S.; Lange, D.; Dralle, H.; Musholt, T.; Jarzab, B.; Paschke, R.; Eszlinger, M. Differential MiRNA Expression Defines Migration and Reduced Apoptosis in Follicular Thyroid Carcinomas. Mol. Cell. Endocrinol. 2014, 388, 1–9. [Google Scholar] [CrossRef]
- Hébrant, A.; Floor, S.; Saiselet, M.; Antoniou, A.; Desbuleux, A.; Snyers, B.; La, C.; de Saint Aubain, N.; Leteurtre, E.; Andry, G.; et al. MiRNA Expression in Anaplastic Thyroid Carcinomas. PLoS ONE 2014, 9. [Google Scholar] [CrossRef]
- Visone, R.; Pallante, P.; Vecchione, A.; Cirombella, R.; Ferracin, M.; Ferraro, A.; Volinia, S.; Coluzzi, S.; Leone, V.; Borbone, E.; et al. Specific MicroRNAs Are Downregulated in Human Thyroid Anaplastic Carcinomas. Oncogene 2007, 26, 7590–7595. [Google Scholar] [CrossRef]
- Boufraqech, M.; Nilubol, N.; Zhang, L.; Gara, S.K.; Sadowski, S.M.; Mehta, A.; He, M.; Davis, S.; Dreiling, J.; Copland, J.A.; et al. MiR30a Inhibits LOX Expression and Anaplastic Thyroid Cancer Progression. Cancer Res. 2015, 75, 367–377. [Google Scholar] [CrossRef]
- Braun, J.; Hoang-Vu, C.; Dralle, H.; Hüttelmaier, S. Downregulation of MicroRNAs Directs the EMT and Invasive Potential of Anaplastic Thyroid Carcinomas. Oncogene 2010, 29, 4237–4244. [Google Scholar] [CrossRef]
- Yao, J.C.; Hassan, M.; Phan, A.; Dagohoy, C.; Leary, C.; Mares, J.E.; Abdalla, E.K.; Fleming, J.B.; Vauthey, J.-N.; Rashid, A.; et al. One Hundred Years after “Carcinoid”: Epidemiology of and Prognostic Factors for Neuroendocrine Tumors in 35,825 Cases in the United States. J. Clin. Oncol. 2008, 26, 3063–3072. [Google Scholar] [CrossRef]
- Hendifar, A.E.; Marchevsky, A.M.; Tuli, R. Neuroendocrine Tumors of the Lung: Current Challenges and Advances in the Diagnosis and Management of Well-Differentiated Disease. J. Thorac. Oncol. 2017, 12, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Malczewska, A.; Kidd, M.; Matar, S.; Kos-Kudla, B.; Modlin, I.M. A Comprehensive Assessment of the Role of MiRNAs as Biomarkers in Gastroenteropancreatic Neuroendocrine Tumors. Neuroendocrinology 2018, 107, 73–90. [Google Scholar] [CrossRef]
- Zatelli, M.C.; Grossrubatscher, E.M.; Guadagno, E.; Sciammarella, C.; Faggiano, A.; Colao, A. Circulating Tumor Cells and MiRNAs as Prognostic Markers in Neuroendocrine Neoplasms. Endocr. Relat. Cancer 2017, 24, R223–R237. [Google Scholar] [CrossRef] [PubMed]
- Abuhatzira, L.; Xu, H.; Tahhan, G.; Boulougoura, A.; Schäffer, A.A.; Notkins, A.L. Multiple microRNAs within the 14q32 cluster target the mRNAs of major type 1 diabetes autoantigens IA-2, IA-2β, and GAD65. FASEB J. 2015, 29, 4374–4383. [Google Scholar] [CrossRef] [PubMed]
- Miller, H.C.; Frampton, A.E.; Malczewska, A.; Ottaviani, S.; Stronach, E.A.; Flora, R.; Kaemmerer, D.; Schwach, G.; Pfragner, R.; Faiz, O.; et al. MicroRNAs Associated with Small Bowel Neuroendocrine Tumours and Their Metastases. Endocr. Relat. Cancer 2016, 23, 711–726. [Google Scholar] [CrossRef] [PubMed]
- Ruebel, K.; Leontovich, A.A.; Stilling, G.A.; Zhang, S.; Righi, A.; Jin, L.; Lloyd, R.V. MicroRNA Expression in Ileal Carcinoid Tumors: Downregulation of MicroRNA-133a with Tumor Progression. Mod. Pathol. 2010, 23, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Heverhagen, A.E.; Legrand, N.; Wagner, V.; Fendrich, V.; Bartsch, D.K.; Slater, E.P. Overexpression of MicroRNA MiR-7-5p Is a Potential Biomarker in Neuroendocrine Neoplasms of the Small Intestine. Neuroendocrinology 2018, 106, 312–317. [Google Scholar] [CrossRef] [PubMed]
- Hamfjord, J.; Stangeland, A.M.; Hughes, T.; Skrede, M.L.; Tveit, K.M.; Ikdahl, T.; Kure, E.H. Differential Expression of MiRNAs in Colorectal Cancer: Comparison of Paired Tumor Tissue and Adjacent Normal Mucosa Using High-Throughput Sequencing. PLoS ONE 2012, 7, e34150. [Google Scholar] [CrossRef] [PubMed]
- Castro-Vega, L.J.; Lepoutre-Lussey, C.; Gimenez-Roqueplo, A.-P.; Favier, J. Rethinking Pheochromocytomas and Paragangliomas from a Genomic Perspective. Oncogene 2016, 35, 1080–1089. [Google Scholar] [CrossRef] [PubMed]
- Pereira, B.D.; Luiz, H.V.; Ferreira, A.G.; Portugal, J. Genetics of Pheochromocytoma and Paraganglioma. In Paraganglioma: A Multidisciplinary Approach; Mariani-Costantini, R., Ed.; Codon Publications: Brisbane, Austrilia, 2019; ISBN 978-0-9944381-7-1. [Google Scholar]
- Patterson, E.; Webb, R.; Weisbrod, A.; Bian, B.; He, M.; Zhang, L.; Holloway, A.K.; Krishna, R.; Nilubol, N.; Pacak, K.; et al. The MicroRNA Expression Changes Associated with Malignancy and SDHB Mutation in Pheochromocytoma. Endocr. Relat. Cancer 2012, 19, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Igaz, P.; Igaz, I.; Nagy, Z.; Nyírő, G.; Szabó, P.M.; Falus, A.; Patócs, A.; Rácz, K. MicroRNAs in Adrenal Tumors: Relevance for Pathogenesis, Diagnosis, and Therapy. Cell. Mol. Life Sci. 2015, 72, 417–428. [Google Scholar] [CrossRef] [PubMed]
- Robertson, S.; MacKenzie, S.M.; Alvarez-Madrazo, S.; Diver, L.A.; Lin, J.; Stewart, P.M.; Fraser, R.; Connell, J.M.; Davies, E. MicroRNA-24 Is a Novel Regulator of Aldosterone and Cortisol Production in the Human Adrenal Cortex. Hypertension 2013, 62, 572–578. [Google Scholar] [CrossRef]
- MacKenzie, S.M.; van Kralingen, J.; Martin, H.; Davies, E. MicroRNAs in Aldosterone Production and Action. Aldosterone Miner. Recept. Cell Biol. Transl. Med. 2019. [Google Scholar] [CrossRef]
- He, J.; Cao, Y.; Su, T.; Jiang, Y.; Jiang, L.; Zhou, W.; Zhang, C.; Wang, W.; Ning, G. Downregulation of MiR-375 in Aldosterone-Producing Adenomas Promotes Tumour Cell Growth via MTDH. Clin. Endocrinol. 2015, 83, 581–589. [Google Scholar] [CrossRef]
- Peng, K.-Y.; Chang, H.-M.; Lin, Y.-F.; Chan, C.-K.; Chang, C.-H.; Chueh, S.-C.J.; Yang, S.-Y.; Huang, K.-H.; Lin, Y.-H.; Wu, V.-C.; et al. MiRNA-203 Modulates Aldosterone Levels and Cell Proliferation by Targeting Wnt5a in Aldosterone-Producing Adenomas. J. Clin. Endocrinol. Metab. 2018, 103, 3737–3747. [Google Scholar] [CrossRef]
- Nakano, Y.; Yoshimoto, T.; Watanabe, R.; Murakami, M.; Fukuda, T.; Saito, K.; Fujii, Y.; Akashi, T.; Tanaka, T.; Yamada, T.; et al. MiRNA299 Involvement in CYP11B2 Expression in Aldosterone-Producing Adenoma. Eur. J. Endocrinol. 2019, 181, 69–78. [Google Scholar] [CrossRef]
- Patterson, E.E.; Holloway, A.K.; Weng, J.; Fojo, T.; Kebebew, E. MicroRNA Profiling of Adrenocortical Tumors Reveals MiR-483 as a Marker of Malignancy. Cancer 2011, 117, 1630–1639. [Google Scholar] [CrossRef]
- Kwok, G.T.Y.; Zhao, J.T.; Glover, A.R.; Gill, A.J.; Clifton-Bligh, R.; Robinson, B.G.; Ip, J.C.Y.; Sidhu, S.B. MicroRNA-431 as a Chemosensitizer and Potentiator of Drug Activity in Adrenocortical Carcinoma. Oncologist 2019, 24, e241–e250. [Google Scholar] [CrossRef] [PubMed]
- Turányi, E.; Dezso, K.; Paku, S.; Nagy, P. DLK Is a Novel Immunohistochemical Marker for Adrenal Gland Tumors. Virchows Arch. 2009, 455, 295–299. [Google Scholar] [CrossRef] [PubMed]
- Aflorei, E.D.; Korbonits, M. Epidemiology and Etiopathogenesis of Pituitary Adenomas. J. Neurooncol. 2014, 117, 379–394. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Mao, Z.-G.; Wang, X.; Du, Q.; Jian, M.; Zhu, D.; Xiao, Z.; Wang, H.-J.; Zhu, Y.-H. MicroRNAs and Target Genes in Pituitary Adenomas. Horm. Metab. Res. 2018, 50, 179–192. [Google Scholar] [CrossRef]
- Wierinckx, A.; Roche, M.; Legras-Lachuer, C.; Trouillas, J.; Raverot, G.; Lachuer, J. MicroRNAs in Pituitary Tumors. Mol. Cell. Endocrinol. 2017, 456, 51–61. [Google Scholar] [CrossRef]
- Vicchio, T.M.; Aliquò, F.; Ruggeri, R.M.; Ragonese, M.; Giuffrida, G.; Cotta, O.R.; Spagnolo, F.; Torre, M.L.; Alibrandi, A.; Asmundo, A.; et al. MicroRNAs Expression in Pituitary Tumors: Differences Related to Functional Status, Pathological Features, and Clinical Behavior. J. Endocrinol. Investig. 2020, 43, 947–958. [Google Scholar] [CrossRef] [PubMed]
- Palumbo, T.; Faucz, F.R.; Azevedo, M.; Xekouki, P.; Iliopoulos, D.; Stratakis, C.A. Functional Screen Analysis Reveals MiR-26b and MiR-128 as Central Regulators of Pituitary Somatomammotrophic Tumor Growth through Activation of the PTEN–AKT Pathway. Oncogene 2013, 32, 1651–1659. [Google Scholar] [CrossRef]
- Beylerli, O.; Beeraka, N.M.; Gareev, I.; Pavlov, V.; Yang, G.; Liang, Y.; Aliev, G. MiRNAs as Noninvasive Biomarkers and Therapeutic Agents of Pituitary Adenomas. Int. J. Mol. Sci. 2020, 21, 7287. [Google Scholar] [CrossRef]
- Gejman, R.; Batista, D.L.; Zhong, Y.; Zhou, Y.; Zhang, X.; Swearingen, B.; Stratakis, C.A.; Hedley-Whyte, E.T.; Klibanski, A. Selective Loss of MEG3 Expression and Intergenic Differentially Methylated Region Hypermethylation in the MEG3/DLK1 Locus in Human Clinically Nonfunctioning Pituitary Adenomas. J. Clin. Endocrinol. Metab. 2008, 93, 4119–4125. [Google Scholar] [CrossRef]
- Zhao, J.; Dahle, D.; Zhou, Y.; Zhang, X.; Klibanski, A. Hypermethylation of the Promoter Region Is Associated with the Loss of MEG3 Gene Expression in Human Pituitary Tumors. J. Clin. Endocrinol. Metab. 2005, 90, 2179–2186. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Riss, D.; Ruebel, K.; Kajita, S.; Scheithauer, B.W.; Horvath, E.; Kovacs, K.; Lloyd, R.V. Galectin-3 Expression in Functioning and Silent ACTH-Producing Adenomas. Endocr. Pathol. 2005, 16, 107–114. [Google Scholar] [CrossRef] [PubMed]
- Riss, D.; Jin, L.; Qian, X.; Bayliss, J.; Scheithauer, B.W.; Young, W.F.; Vidal, S.; Kovacs, K.; Raz, A.; Lloyd, R.V. Differential Expression of Galectin-3 in Pituitary Tumors. Cancer Res. 2003, 63, 2251–2255. [Google Scholar]
- Ruebel, K.H.; Jin, L.; Qian, X.; Scheithauer, B.W.; Kovacs, K.; Nakamura, N.; Zhang, H.; Raz, A.; Lloyd, R.V. Effects of DNA Methylation on Galectin-3 Expression in Pituitary Tumors. Cancer Res. 2005, 65, 1136–1140. [Google Scholar] [CrossRef]
- Zhang, H.-Y.; Jin, L.; Stilling, G.A.; Ruebel, K.H.; Coonse, K.; Tanizaki, Y.; Raz, A.; Lloyd, R.V. RUNX1 and RUNX2 Upregulate Galectin-3 Expression in Human Pituitary Tumors. Endocrine 2009, 35, 101–111. [Google Scholar] [CrossRef]
- Raue, F.; Frank-Raue, K. Epidemiology and Clinical Presentation of Medullary Thyroid Carcinoma. Recent Results Cancer Res. 2015, 204, 61–90. [Google Scholar] [CrossRef] [PubMed]
- Malek, A.; Sleptsov, I.E.; Cheburkin, Y.V.; Roman, S.; Kolesnikov, N. MiRNA as Potential Tool for Thyroid Cancer Diagnostics and Follow up: Practical Considerations. JSM Thyroid Disord. Manag. 2017, 2, 1007. [Google Scholar]
- Romeo, P.; Colombo, C.; Granata, R.; Calareso, G.; Gualeni, A.V.; Dugo, M.; De Cecco, L.; Rizzetti, M.G.; Zanframundo, A.; Aiello, A.; et al. Circulating MiR-375 as a Novel Prognostic Marker for Metastatic Medullary Thyroid Cancer Patients. Endocr. Relat. Cancer 2018, 25, 217–231. [Google Scholar] [CrossRef]
- Saiselet, M.; Pita, J.M.; Augenlicht, A.; Dom, G.; Tarabichi, M.; Fimereli, D.; Dumont, J.E.; Detours, V.; Maenhaut, C. MiRNA Expression and Function in Thyroid Carcinomas: A Comparative and Critical Analysis and a Model for Other Cancers. Oncotarget 2016, 7, 52475–52492. [Google Scholar] [CrossRef]
- Boufraqech, M.; Klubo-Gwiezdzinska, J.; Kebebew, E. MicroRNAs in the Thyroid. Best Pract. Res. Clin. Endocrinol. Metab. 2016, 30, 603–619. [Google Scholar] [CrossRef]
- Fiore, R.; Khudayberdiev, S.; Christensen, M.; Siegel, G.; Flavell, S.W.; Kim, T.-K.; Greenberg, M.E.; Schratt, G. Mef2-Mediated Transcription of the MiR379-410 Cluster Regulates Activity-Dependent Dendritogenesis by Fine-Tuning Pumilio2 Protein Levels. EMBO J. 2009, 28, 697–710. [Google Scholar] [CrossRef] [PubMed]
- Aavik, E.; Lumivuori, H.; Leppänen, O.; Wirth, T.; Häkkinen, S.-K.; Bräsen, J.-H.; Beschorner, U.; Zeller, T.; Braspenning, M.; van Criekinge, W.; et al. Global DNA Methylation Analysis of Human Atherosclerotic Plaques Reveals Extensive Genomic Hypomethylation and Reactivation at Imprinted Locus 14q32 Involving Induction of a MiRNA Cluster. Eur. Heart J. 2015, 36, 993–1000. [Google Scholar] [CrossRef] [PubMed]
- Romitti, M.; Wajner, S.M.; Zennig, N.; Goemann, I.M.; Bueno, A.L.; Meyer, E.L.S.; Maia, A.L. Increased Type 3 Deiodinase Expression in Papillary Thyroid Carcinoma. Thyroid 2012, 22, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Greife, A.; Knievel, J.; Ribarska, T.; Niegisch, G.; Schulz, W.A. Concomitant Downregulation of the Imprinted Genes DLK1 and MEG3 at 14q32.2 by Epigenetic Mechanisms in Urothelial Carcinoma. Clin. Epigenetics 2014, 6, 29. [Google Scholar] [CrossRef]
- Jishnu, P.V.; Jayaram, P.; Shukla, V.; Varghese, V.K.; Pandey, D.; Sharan, K.; Chakrabarty, S.; Satyamoorthy, K.; Kabekkodu, S.P. Prognostic Role of 14q32.31 MiRNA Cluster in Various Carcinomas: A Systematic Review and Meta-Analysis. Clin. Exp. Metastasis 2020, 37, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Shu, J.; Li, L.; Sarver, A.E.; Pope, E.A.; Varshney, J.; Thayanithy, V.; Spector, L.; Largaespada, D.A.; Steer, C.J.; Subramanian, S. Imprinting Defects at Human 14q32 Locus Alters Gene Expression and Is Associated with the Pathobiology of Osteosarcoma. Oncotarget 2016, 7, 21298–21314. [Google Scholar] [CrossRef] [PubMed]
- Manodoro, F.; Marzec, J.; Chaplin, T.; Miraki-Moud, F.; Moravcsik, E.; Jovanovic, J.V.; Wang, J.; Iqbal, S.; Taussig, D.; Grimwade, D.; et al. Loss of Imprinting at the 14q32 Domain Is Associated with MicroRNA Overexpression in Acute Promyelocytic Leukemia. Blood 2014, 123, 2066–2074. [Google Scholar] [CrossRef]
- Romitti, M.; Wajner, S.M.; Ceolin, L.; Ferreira, C.V.; Ribeiro, R.V.P.; Rohenkohl, H.C.; Weber, S.D.S.; Lopez, P.L.D.C.; Fuziwara, C.S.; Kimura, E.T.; et al. MAPK and SHH Pathways Modulate Type 3 Deiodinase Expression in Papillary Thyroid Carcinoma. Endocr. Relat. Cancer 2016, 23, 135–146. [Google Scholar] [CrossRef]
- Gardiner, E.; Beveridge, N.J.; Wu, J.Q.; Carr, V.; Scott, R.J.; Tooney, P.A.; Cairns, M.J. Imprinted DLK1-DIO3 Region of 14q32 Defines a Schizophrenia-Associated MiRNA Signature in Peripheral Blood Mononuclear Cells. Mol. Psychiatry 2012, 17, 827–840. [Google Scholar] [CrossRef]
- Stark, K.L.; Xu, B.; Bagchi, A.; Lai, W.-S.; Liu, H.; Hsu, R.; Wan, X.; Pavlidis, P.; Mills, A.A.; Karayiorgou, M.; et al. Altered Brain MicroRNA Biogenesis Contributes to Phenotypic Deficits in a 22q11-Deletion Mouse Model. Nat. Genet. 2008, 40, 751–760. [Google Scholar] [CrossRef]
- Schaefer, A.; Im, H.-I.; Venø, M.T.; Fowler, C.D.; Min, A.; Intrator, A.; Kjems, J.; Kenny, P.J.; O’Carroll, D.; Greengard, P. Argonaute 2 in Dopamine 2 Receptor–Expressing Neurons Regulates Cocaine Addiction. J. Exp. Med. 2010, 207, 1843–1851. [Google Scholar] [CrossRef]
- Diederichs, S.; Haber, D.A. Dual Role for Argonautes in MicroRNA Processing and Posttranscriptional Regulation of MicroRNA Expression. Cell 2007, 131, 1097–1108. [Google Scholar] [CrossRef] [PubMed]
- O’Carroll, D.; Mecklenbrauker, I.; Das, P.P.; Santana, A.; Koenig, U.; Enright, A.J.; Miska, E.A.; Tarakhovsky, A. A Slicer-Independent Role for Argonaute 2 in Hematopoiesis and the MicroRNA Pathway. Genes Dev. 2007, 21, 1999–2004. [Google Scholar] [CrossRef]
- Welten, S.M.J.; de Vries, M.R.; Peters, E.A.B.; Agrawal, S.; Quax, P.H.A.; Nossent, A.Y. Inhibition of Mef2a Enhances Neovascularization via Post-Transcriptional Regulation of 14q32 MicroRNAs MiR-329 and MiR-494. Mol. Ther. Nucleic Acids 2017, 7, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Downie Ruiz Velasco, A.; Welten, S.M.J.; Goossens, E.A.C.; Quax, P.H.A.; Rappsilber, J.; Michlewski, G.; Nossent, A.Y. Posttranscriptional Regulation of 14q32 MicroRNAs by the CIRBP and HADHB during Vascular Regeneration after Ischemia. Mol. Ther. Nucleic Acids 2019, 14, 329–338. [Google Scholar] [CrossRef] [PubMed]
- Rosa, A.L.; Wu, Y.-Q.; Kwabi-Addo, B.; Coveler, K.J.; Reid Sutton, V.; Shaffer, L.G. Allele-Specific Methylation of a Functional CTCF Binding Site Upstream of MEG3 in the Human Imprinted Domain of 14q32. Chromosome Res. 2005, 13, 809–818. [Google Scholar] [CrossRef] [PubMed]
- Bell, A.C.; West, A.G.; Felsenfeld, G. The Protein CTCF Is Required for the Enhancer Blocking Activity of Vertebrate Insulators. Cell 1999, 98, 387–396. [Google Scholar] [CrossRef]
- Szabó, P.E.; Tang, S.-H.E.; Rentsendorj, A.; Pfeifer, G.P.; Mann, J.R. Maternal-Specific Footprints at Putative CTCF Sites in the H19 Imprinting Control Region Give Evidence for Insulator Function. Curr. Biol. 2000, 10, 607–610. [Google Scholar] [CrossRef]
- Welten, S.M.J.; Bastiaansen, A.J.N.M.; de Jong, R.C.M.; de Vries, M.R.; Peters, E.A.B.; Boonstra, M.C.; Sheikh, S.P.; La Monica, N.; Kandimalla, E.R.; Quax, P.H.A.; et al. Inhibition of 14q32 MicroRNAs MiR-329, MiR-487b, MiR-494, and MiR-495 Increases Neovascularization and Blood Flow Recovery after Ischemia. Circ. Res. 2014, 115, 696–708. [Google Scholar] [CrossRef]
- Gupta, G.P.; Massagué, J. Cancer Metastasis: Building a Framework. Cell 2006, 127, 679–695. [Google Scholar] [CrossRef]
- Uppal, A.; Ferguson, M.K.; Posner, M.C.; Hellman, S.; Khodarev, N.N.; Weichselbaum, R.R. Towards a Molecular Basis of Oligometastatic Disease: Potential Role of Micro-RNAs. Clin. Exp. Metastasis 2014, 31, 735–748. [Google Scholar] [CrossRef]
- Qian, P.; He, X.C.; Paulson, A.; Li, Z.; Tao, F.; Perry, J.M.; Guo, F.; Zhao, M.; Zhi, L.; Venkatraman, A.; et al. The Dlk1-Gtl2 Locus Preserves LT-HSC Function by Inhibiting the PI3K-MTOR Pathway to Restrict Mitochondrial Metabolism. Cell Stem Cell 2016, 18, 214–228. [Google Scholar] [CrossRef] [PubMed]
- Thayanithy, V.; Sarver, A.L.; Kartha, R.V.; Li, L.; Angstadt, A.Y.; Breen, M.; Steer, C.J.; Modiano, J.F.; Subramanian, S. Perturbation of 14q32 MiRNAs-CMYC Gene Network in Osteosarcoma. Bone 2012, 50, 171–181. [Google Scholar] [CrossRef] [PubMed]
- Feldmann, G.; Mishra, A.; Hong, S.-M.; Bisht, S.; Strock, C.J.; Ball, D.W.; Goggins, M.; Maitra, A.; Nelkin, B.D. Inhibiting the Cyclin-Dependent Kinase CDK5 Blocks Pancreatic Cancer Formation and Progression through the Suppression of Ras-Ral Signaling. Cancer Res. 2010, 70, 4460–4469. [Google Scholar] [CrossRef]
- Reis, E.M.; Nakaya, H.I.; Louro, R.; Canavez, F.C.; Flatschart, A.V.F.; Almeida, G.T.; Egidio, C.M.; Paquola, A.C.; Machado, A.A.; Festa, F.; et al. Antisense Intronic Non-Coding RNA Levels Correlate to the Degree of Tumor Differentiation in Prostate Cancer. Oncogene 2004, 23, 6684–6692. [Google Scholar] [CrossRef] [PubMed]
- Konoplev, S.N.; Fritsche, H.A.; O’Brien, S.; Wierda, W.G.; Keating, M.J.; Gornet, T.G.; St Romain, S.; Wang, X.; Inamdar, K.; Johnson, M.R.; et al. High Serum Thymidine Kinase 1 Level Predicts Poorer Survival in Patients with Chronic Lymphocytic Leukemia. Am. J. Clin. Pathol. 2010, 134, 472–477. [Google Scholar] [CrossRef]
- Xu, Y.; Shi, Q.-L.; Ma, H.; Zhou, H.; Lu, Z.; Yu, B.; Zhou, X.; Eriksson, S.; He, E.; Skog, S. High Thymidine Kinase 1 (TK1) Expression Is a Predictor of Poor Survival in Patients with PT1 of Lung Adenocarcinoma. Tumour Biol. 2012, 33, 475–483. [Google Scholar] [CrossRef]
- He, E.; Xu, X.H.; Guan, H.; Chen, Y.; Chen, Z.H.; Pan, Z.L.; Tang, L.L.; Hu, G.Z.; Li, Y.; Zhang, M.; et al. Thymidine Kinase 1 Is a Potential Marker for Prognosis and Monitoring the Response to Treatment of Patients with Breast, Lung, and Esophageal Cancer and Non-Hodgkin’s Lymphoma. Nucleosides Nucleotides Nucleic Acids 2010, 29, 352–358. [Google Scholar] [CrossRef]
- Pathak, S.; Meng, W.-J.; Nandy, S.K.; Ping, J.; Bisgin, A.; Helmfors, L.; Waldmann, P.; Sun, X.-F. Radiation and SN38 Treatments Modulate the Expression of MicroRNAs, Cytokines and Chemokines in Colon Cancer Cells in a P53-Directed Manner. Oncotarget 2015, 6, 44758–44780. [Google Scholar] [CrossRef]
- Liu, D.; Zhang, X.; Yan, C.; Li, Y.; Tian, X.; Zhu, N.; Rong, J.; Peng, C.; Han, Y. MicroRNA-495 Regulates the Proliferation and Apoptosis of Human Umbilical Vein Endothelial Cells by Targeting Chemokine CCL2. Thromb. Res. 2015, 135, 146–154. [Google Scholar] [CrossRef]
- Wilhelm, K.; Happel, K.; Eelen, G.; Schoors, S.; Oellerich, M.F.; Lim, R.; Zimmermann, B.; Aspalter, I.M.; Franco, C.A.; Boettger, T.; et al. FOXO1 Couples Metabolic Activity and Growth State in the Vascular Endothelium. Nature 2016, 529, 216–220. [Google Scholar] [CrossRef]
- Yao, G.-D.; Zhang, Y.-F.; Chen, P.; Ren, X.-B. MicroRNA-544 Promotes Colorectal Cancer Progression by Targeting Forkhead Box O1. Oncol. Lett. 2018, 15, 991–997. [Google Scholar] [CrossRef]
- Jeon, H.H.; Yu, Q.; Lu, Y.; Spencer, E.; Lu, C.; Milovanova, T.; Yang, Y.; Zhang, C.; Stepanchenko, O.; Vafa, R.P.; et al. FOXO1 Regulates VEGFA Expression and Promotes Angiogenesis in Healing Wounds. J. Pathol. 2018, 245, 258–264. [Google Scholar] [CrossRef]
- Haig, D.; Mainieri, A. The Evolution of Imprinted MicroRNAs and Their RNA Targets. Genes 2020, 11, 1038. [Google Scholar] [CrossRef] [PubMed]
- Haig, D. Kin Conflict in Seed Development: An Interdependent but Fractious Collective. Annu. Rev. Cell Dev. Biol. 2013, 29, 189–211. [Google Scholar] [CrossRef] [PubMed]
- Haig, D. Imprinted Green Beards: A Little Less than Kin and More than Kind. Biol. Lett. 2013, 9. [Google Scholar] [CrossRef]
- Patten, M.M.; Ross, L.; Curley, J.P.; Queller, D.C.; Bonduriansky, R.; Wolf, J.B. The Evolution of Genomic Imprinting: Theories, Predictions and Empirical Tests. Heredity 2014, 113, 119–128. [Google Scholar] [CrossRef]
- Pallante, P.; Sepe, R.; Puca, F.; Fusco, A. High Mobility Group A Proteins as Tumor Markers. Front. Med. 2015, 2. [Google Scholar] [CrossRef]
- Fusco, A.; Fedele, M. Roles of HMGA Proteins in Cancer. Nat. Rev. Cancer 2007, 7, 899–910. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Tumor Type | Study | miRNA Profiling Platform | |
|---|---|---|---|
| NET | Lung | Yoshimoto et al., 2018 [57] | microarray |
| Mairinger et al., 2014 [58] | TaqMan array | ||
| Deng et al., 2014 [59] | microarray | ||
| Rapa et al., 2015 [60] | PCR array | ||
| Wong et al., 2020 [61] | NGS | ||
| Pancreas | Zimmermann et al., 2018 [62] | TaqMan array | |
| Roldo et al., 2006 [63] | microarray | ||
| Jiang et al., 2015 [64] | PCR array | ||
| Zhou et al., 2016 [65] | microarray (GSE43796) reanalysis | ||
| Lee et al., 2015 [66] | Nanostring nCounter | ||
| small intestinal | Yoshimoto et al., 2018 [57] | microarray | |
| Arvidsson et al., 2018 [67] | microarray | ||
| Li et al., 2013 [68] | microarray | ||
| Miller et al., 2016 | Nanostring nCounter | ||
| PPGL | Castro-Vega et al., 2015 [69] | NGS | |
| Tömböl et al., 2010 [70] | TaqMan array | ||
| de Cubas et al., 2013 [71] | microarray | ||
| Meyer-Rochow et al., 2010 [72] | microarray | ||
| Calsina et al., 2019 [73] | individual qPCR | ||
| ACC | Tömböl et al., 2009 [74] | TaqMan array | |
| Chabre et al., 2012 [54] | microarray | ||
| Özata et al., 2011 [75] | microarray | ||
| Assié et al., 2014 [76] | NGS | ||
| Pituitary | GH | Mao et al., 2010 [77] | microarray |
| D’Angelo et al., 2012 [78] | microarray | ||
| Bottoni et al., 2007 [79] | microarray | ||
| Butz et al., 2009 [80] | TaqMan array | ||
| Cheunsuchon et al., 2011 [81] | individual qPCR | ||
| He et al., 2019 [82] | NGS | ||
| ACTH | Gentilin et al., 2013 [83] | individual TaqMan assay | |
| Amaral et al., 2009 [84] | individual TaqMan assay | ||
| Stilling et al., 2010 [85] | microarray | ||
| Cheunsuchon et al., 2011 [81] | individual qPCR | ||
| PRL | He et al., 2019 [82] | NGS | |
| Müssnich et al., 2015 [86] | microarray | ||
| Chen et al., 2012 [87] | NGS | ||
| Cheunsuchon et al., 2011 [81] | individual qPCR | ||
| NFPA | He et al., 2019 [82] | NGS | |
| Darvasi et al., 2019 [88] | NGS, TaqMan array and microarray | ||
| Butz et al., 2011 [89] | TaqMan array | ||
| Liang et al., 2015 [90] | individual qPCR | ||
| Cheunsuchon et al., 2011 [81] | individual qPCR | ||
| Müssnich et al., 2015 [86] | microarray | ||
| Bottoni et al., 2007 [79] | microarray | ||
| OC | Krokker et al., 2019 [91] | NGS | |
| Thyroid | MTC | Lassalle et al., 2016 [92] | microarray |
| Hudson et al., 2013 [93] | Taqman array | ||
| Nikiforova et al., 2008 [94] | Taqman array | ||
| PTC | Geraldo et al., 2017 [95] | NGS (obtained from The Cancer Genome Atlas dataset) | |
| Rosignolo et al., 2017 [96] | Taqman array | ||
| Tetzlaff et al., 2007 [97] | microarray | ||
| Linwah et al., 2011 [98] | microarray | ||
| Jacques et al., 2013 [99] | microarray | ||
| Lassalle et al., 2011 [100] | microarray | ||
| Mancikova et al., 2015 [101] | NGS | ||
| Peng et al., 2014 [102] | microarray | ||
| Riesco-Eizaguirre et al., 2015 [103] | NGS | ||
| Saiselet et al., 2015 [104] | NGS | ||
| Swierniak et al., 2013 [105] | NGS | ||
| FTC | Nikiforova et al., 2008 [94] | TaqMan array | |
| Rossing et al., 2012 [106] | microarray | ||
| Dettmer et al., 2013 [107] | Taqman array | ||
| Jacques et al., 2013 [99] | microarray | ||
| Lassalle et al., 2011 [100] | microarray | ||
| Mancikova et al., 2015 [101] | NGS | ||
| Wojtas et al., 2014 [108] | microarray | ||
| ATC | Hébrant et al., 2014 [109] | microarray | |
| Visone et al., 2007 [110] | microarray | ||
| Boufraqech et al., 2015 [111] | microarray | ||
| Braun et al., 2010 [112] | microarray | ||
| NET | |
| miR-485-3p | increased in the metastatic tumors compared to the primary pNET |
| miR-494 | overexpressed in metastases compared to primary siNET |
| downregulated in carcinoid vs. adenocarcinoma/normal lung tissue | |
| miR-376a, miR-376b, miR-381, miR-409-3p, miR-409-5p, | upregulated in typical compared to atypical lung carcinoids |
| miR-127, miR-136, miR-154, miR-369, miR-370, miR-376a, miR-410, miR-432, miR-409, miR-487b, miR-494 | downregulated in lung carcinoid compared to adjacent normal tissue |
| miR-409-3p, miR-409-5p, miR-411, miR-431-5p, miR-485 and miR-539 | downregulated in metastatic carcinoids compared to non-metastatic lung NET |
| miR-127, miR-136, miR-154, miR-485, miR-770-5p | negative correlation with tumor biology of lung NET |
| PPGL | |
| miR-493-5p | commonly downregulated in all PCC molecular subtypes (based on germline mutation) |
| miR-127-3p, miR-136, miR-154-3p/5p, miR-323a-3p, miR-337-5p/-3p, miR-369-5p, miR-370, miR-376a-5p, miR-376c, miR-377, miR-382, miR-409-5p, miR-410, miR-485-3p és 5p, miR-487a, miR-495, miR-539, miR-543, miR-758, miR-889 | downregulation in MAX-related PPGLs and a subset of sporadic PCC |
| miR-154-3p, hsa-miR-369-5p, hsa-miR-485-5p, hsa-miR-487a, hsa-miR-495, hsa-miR-543, hsa-miR-656, hsa-miR-889 | overexpression in TMEM127-related PPGL cases |
| miR-541 | overexpressed in VHL-related PCC vs. sporadic PCC, decreased expression in recurrent tumors compared to primary tumors |
| miR-154, miR-337-3p | upregulated in a subset of metastatic PCC compared to non-metastatic cases |
| miR-409-3p, miR-369-3p | downregulation in a subset of metastatic PCC compared to benign PCC |
| miR-431 | upregulated in malignant tumors compared to benign |
| Adrenocortical Tumors | |
| miR-370 | overexpressed in APA compared to non-APA adrenal tumors |
| miR-299 | downregulated in KCNJ5 mutant APA vs. non-KCNJ5 mutant samples |
| 14q32 miRNA cluster | whole miRNA cluster downregulation in Mi1 subset of ACC |
| miR-136, miR-127-3p, miR-487b, miR-376c and miR-432 | overexpressed in ACC compared to normal adrenal cortex |
| miR-376a, miR-376b | overexpression in ACC vs. ACA |
| miR-376a | downregulated in ACC vs. NF adenoma, CPA and normal adrenal cortex |
| miR-299-5p, miR-485-5p | overexpressed in ACC vs. NF adenoma, CPA and normal adrenal cortex |
| miR-370, miR-376a, miR-376b, miR-376c, miR-377, miR-379, miR-382, miR-411, miR-487a, miR-494, miR-495 | downregulated in non-aggressive ACC as compared to aggressive ones |
| miRNA-665 | overexpressed in ACC as compared to benign adrenocortical tumors |
| miR-431 | implicated in adjuvant therapy response in ACC |
| PitNET | |
| miR-127-3p, miR-154, miR-329, miR-337, miR-369-5p, miR-376c, miR-432, miR-433 | downregulated in PRL adenoma vs. normal |
| miR-410 | overexpressed in PRL adenoma vs. normal |
| miR-411-3p | overexpressed in GH adenoma vs. normal |
| miR-381, miR-654-3p | downregulated in GH adenoma vs. normal |
| miR-127, miR-134, miR-136, miR-154, miR-323a, miR-337, miR-369, miR-370, miR-376a-1, miR-376a-2, miR-376b, miR-376c, miR-379, miR-380, miR-381, miR-382, miR-409, miR-410, miR-411, miR-431, miR-432, miR-433, miR-487b, miR-493, miR-494, miR-495, miR-539, miR-543, miR-544a, miR-654, miR-656, miR-770-5p, miR-889 | downregulated in NF adenoma vs. normal |
| miR-1185-1-3p | upregulated in NF adenoma vs. normal |
| miR-127-3p, miR-136, miR-154, miR-299-5p, miR-323-5p, miR-329, miR-369-3p, miR-369-5p, miR-376c, miR-377, miR-411-3p, miR-431-3p, miR-433, miR-493 | downregulated in corticitroph adenoma vs. normal |
| miR-431, miR-493 | overexpressed in corticotroph carcinoma vs. adenoma |
| miR-127, miR-136, miR-154, miR-299, miR-323a, miR-323b, miR-329-1, miR-329-2, miR-369, miR-370, miR-376a-1, miR-376a-2, miR-376b, miR-376c, miR-379, miR-381, miR-382, miR-409, miR-411, miR-431, miR-485, miR-487b, miR-494, miR-539, miR-654, miR-889 | downregulated in oncocytoma vs. normal |
| Thyroid Carcinoma | |
| miR-9, miR-127, miR-136, miR-154, miR-323, miR-376a,c, miR-370, miR-487b | upregulated in MTC vs. normal |
| miR-299 | downregulated in FTC |
| miR-134, miR-136, miR-409, miR-654 | overexpressed in PTC |
| miR-134, miR-300, miR-379, miR-382, miR-494-3p, miR-494-5p, miR-495 | downregulated in PTC |
| miR-654-3p | inverse correlation with PTC progression |
| KEGG Pathway | p-Value | # Genes | # miRNAs |
|---|---|---|---|
| Hippo signaling pathway (hsa04390) | 2.635 × 10−7 | 103 | 47 |
| Proteoglycans in cancer (hsa05205) | 2.507 × 10−6 | 132 | 48 |
| Pathways in cancer (hsa05200) | 3.424 × 10−6 | 255 | 48 |
| Adherens junction (hsa04520) | 1.345 × 10−5 | 57 | 41 |
| TGF-β signaling pathway (hsa04350) | 1.582 × 10−5 | 58 | 45 |
| Axon guidance (hsa04360) | 2.465 × 10−5 | 88 | 45 |
| Rap1 signaling pathway (hsa04015) | 3.946 × 10−5 | 141 | 48 |
| Glioma (hsa05214) | 4.825 × 10−5 | 47 | 43 |
| Ras signaling pathway (hsa04014) | 4.825 × 10−5 | 146 | 49 |
| Circadian rhythm (hsa04710) | 6.429 × 10−5 | 27 | 37 |
| Lysine degradation (hsa00310) | 9.643 × 10−5 | 33 | 43 |
| Signaling pathways regulating pluripotency of stem cells (hsa04550) | 0.0001 | 96 | 50 |
| FoxO signaling pathway (hsa04068) | 0.0001 | 92 | 46 |
| Thyroid hormone signaling pathway (hsa04919) | 0.0001 | 79 | 46 |
| Ubiquitin mediated proteolysis (hsa04120) | 0.0004 | 93 | 44 |
| Dorso-ventral axis formation (hsa04320) | 0.0006 | 24 | 36 |
| Prion diseases (hsa05020) | 0.0009 | 17 | 26 |
| ErbB signaling pathway (hsa04012) | 0.0011 | 63 | 45 |
| Renal cell carcinoma (hsa05211) | 0.0015 | 48 | 41 |
| Pancreatic cancer (hsa05212) | 0.0023 | 48 | 43 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krokker, L.; Patócs, A.; Butz, H. Essential Role of the 14q32 Encoded miRNAs in Endocrine Tumors. Genes 2021, 12, 698. https://doi.org/10.3390/genes12050698
Krokker L, Patócs A, Butz H. Essential Role of the 14q32 Encoded miRNAs in Endocrine Tumors. Genes. 2021; 12(5):698. https://doi.org/10.3390/genes12050698
Chicago/Turabian StyleKrokker, Lilla, Attila Patócs, and Henriett Butz. 2021. "Essential Role of the 14q32 Encoded miRNAs in Endocrine Tumors" Genes 12, no. 5: 698. https://doi.org/10.3390/genes12050698
APA StyleKrokker, L., Patócs, A., & Butz, H. (2021). Essential Role of the 14q32 Encoded miRNAs in Endocrine Tumors. Genes, 12(5), 698. https://doi.org/10.3390/genes12050698