
Autochthonous Austrian Varieties of Prunus avium L. Represent a Regional Gene Pool, Assessed Using SSR and AFLP Markers

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction
2.3. SSR Analysis
2.4. AFLP-Analysis
2.5. Data Analysis and Statistics:
3. Results
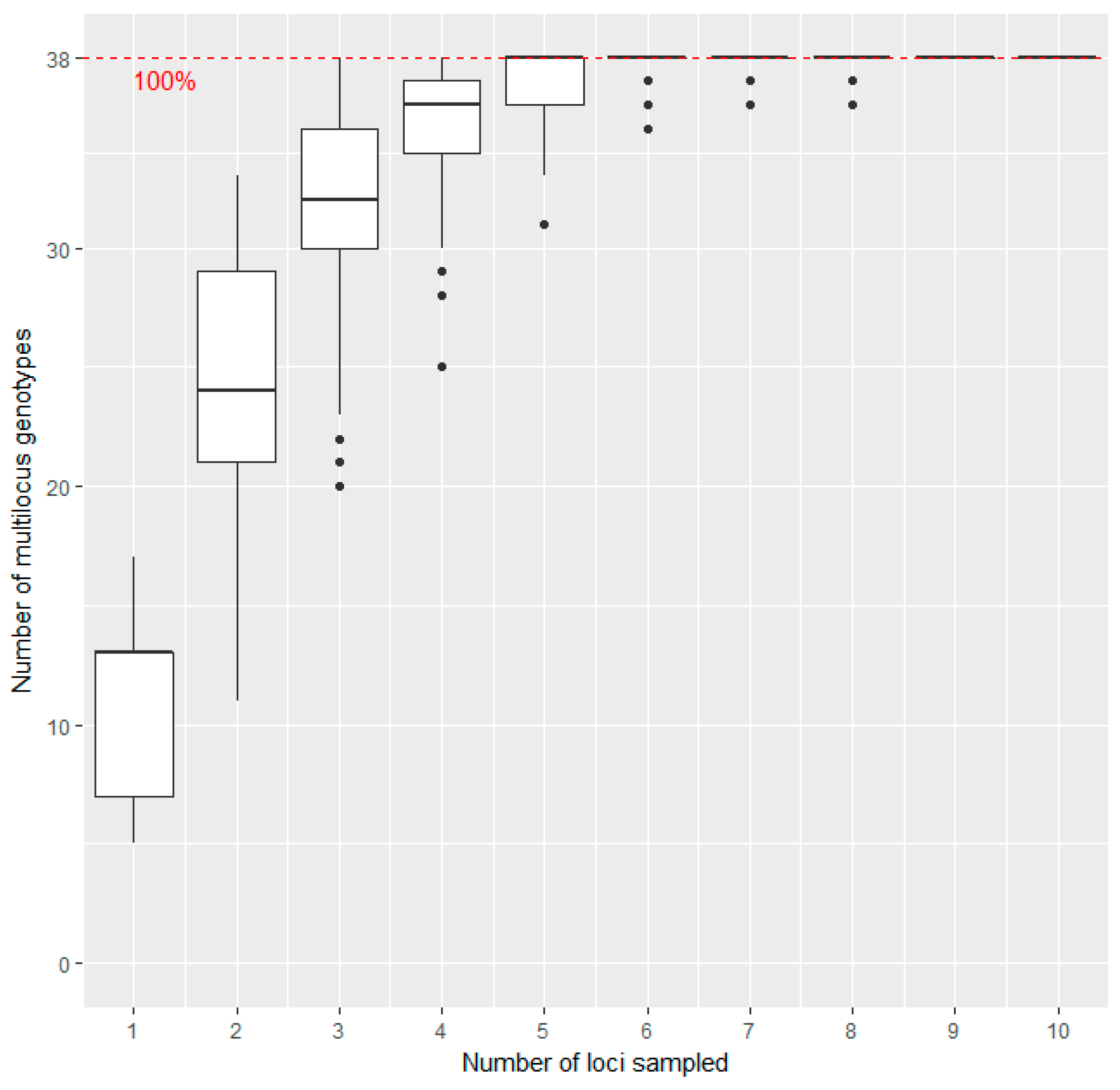
3.1. Method Evaluation of SSR-Analysis
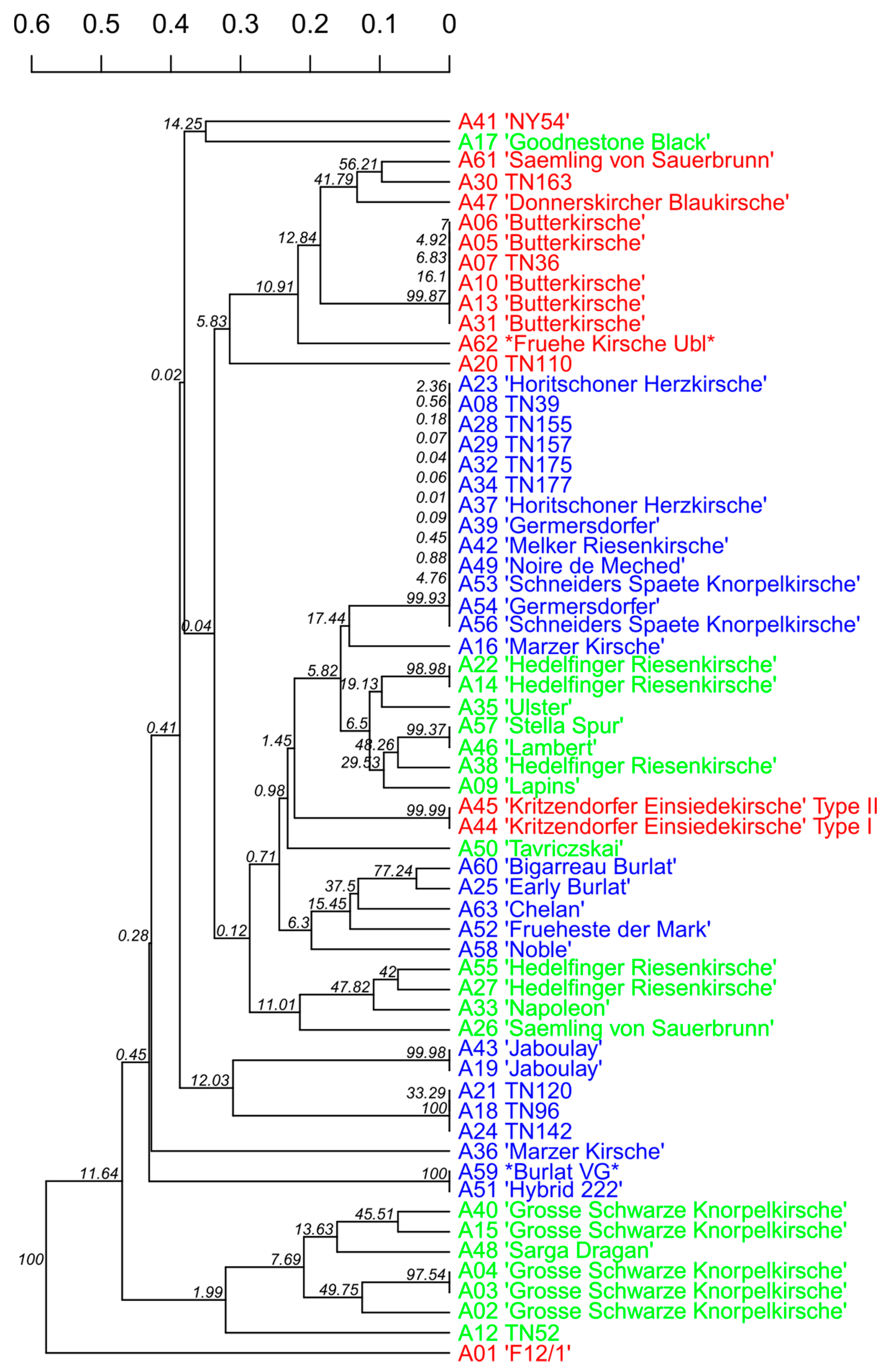
3.2. Variety Identification and Verification of Trueness to Type
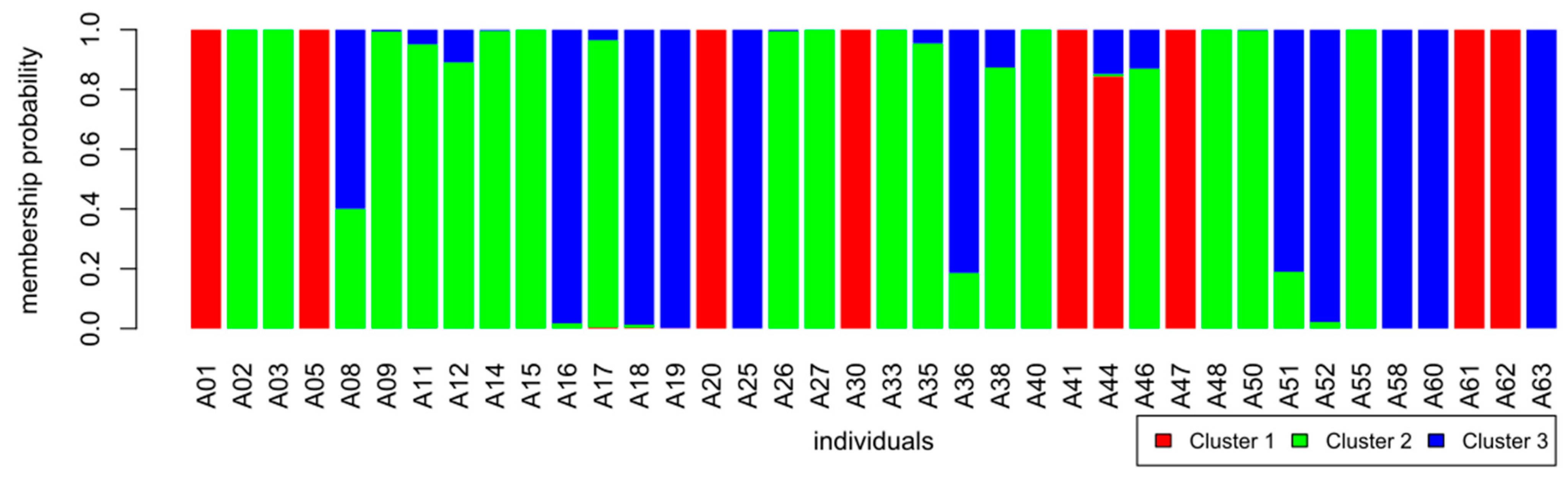
3.3. Genetic Diversity
3.4. Results from AFLP-Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gecer, M.K.; Kan, T.; Gundogdu, M.; Ercisli, S.; Ilhan, G.; Sagbas, H.I. Physicochemical characteristics of wild and cultivated apricots (Prunus armeniaca L.) from Aras valley in Turkey. Genet. Resour. Crop Evol. 2020, 67, 935–945. [Google Scholar] [CrossRef]
- Mariette, S.; Tavaud, M.; Arunyawat, U.; Capdeville, G.; Millan, M.; Salin, F. Population structure and genetic bottleneck in sweet cherry estimated with SSRs and the gametophytic self-incompatibility locus. Genetics 2010, 11, 1471–2156. [Google Scholar] [CrossRef]
- Kocsisné, G.M.; Bolla, D.; Anhalt-Brüderl, U.C.M.; Forneck, A.; Taller, J.; Kocsis, L. Genetic diversity and similarity of pear (Pyrus communis L.) cultivars in Central Europe revealed by SSR markers. Genet. Resour. Crop Evol. 2020, 67, 1755–1763. [Google Scholar] [CrossRef]
- Mihaljevic, M.Z.; Anhalt, U.C.M.; Rühl, E.; Mugosa, M.T.; Maras, V.; Forneck, A.; Zdunic, G.; Preiner, D.; Pejic, I. Cultivar Identity, Intravarietal Variation, and Health Status of Native Grapevine Varieties in Croatia and Montenegro. Am. J. Enol. Vitic. 2015, 66, 531–541. [Google Scholar] [CrossRef]
- Roversi, A.; Monteforte, A.; Panelli, D.; Folini, L.; Fajt, N. Observations on the occurrence of sweet cherry double-fruits in Italy and Slovenia. Acta Hortic. ISHS 2008, 849–854. [Google Scholar] [CrossRef]
- Sekse, L. Fruit Cracking in Sweet Cherries—Some Recent Advances. Acta Hortic. 2008, 615–624. [Google Scholar] [CrossRef]
- Clarke, J.B.; Tobutt, K.R. A Standard Set of Accessions, Microsatellites and Genotypes for Harmonising the Fingerprinting of Cherry Collections for the ECPGR. Acta Hortic. ISHS 2009, 814, 615–618. [Google Scholar] [CrossRef]
- Bodo, F. Burgenlands Kirschensorten; Victor Horáth: Neusiedl am See, Austria, 1936. [Google Scholar]
- BMNT. Grüner Bericht 2018. Bericht Über die Situation der Österreichischen Land- und Forstwirtschaft im Jahr 2017. 2018. Available online: www.gruenerbericht.at (accessed on 23 February 2021).
- Leifer, H. Kartierung und Beschreibung von Kirschbäumen und alten Kirschensorten in Pöttsching (Burgenland). Diplomar-Beit; University of Natural Resources and Life Sciences: Vienna, Austria, 2002. [Google Scholar]
- Spörr, T.; Schüller, E.; Keppel, H.; Spornberger, A. Mapping of regionally typical old cherry cultivars in the gourmet region ‘Genussregion Leithaberger Edelkirsche’. Mitteilungen Klosterneuburg 2014, 64, 82–95. [Google Scholar]
- Schüller, E.; Pilz, V.; Holler, C.; Keppel, H.; Spornberger, A. Mapping and description of old cherry trees and regional cherry cultivars in Stoob, Mittelburgenland. Mitteilungen Klosterneuburg 2016, 66, 113–126. [Google Scholar]
- Putz, S. Survey and Identification of Old Local Sweet Cherry (Prunus avium) cultivars in Scharten, Upper Austria. Master’s Thesis, University of Natural Resources and Life Sciences, Vienna, Austria, 2014. [Google Scholar]
- Schoedl, K.; Denk, A.; Hummelbrunner, S.; Modl, P.; Forneck, A. No improvement in fruit quality through chemical flower thinning in sweet cherry (Prunus avium L.). J. Sci. Food Agric. 2009, 89, 1236–1240. [Google Scholar] [CrossRef]
- Wünsch, A.; Hormaza, J.I. Cultivar identification and genetic fingerprinting of temperate fruit tree species using DNA markers. Euphytica 2002, 125, 59–67. [Google Scholar] [CrossRef]
- Turkoglu, Z.; Bilgener, S.; Ercisli, S.; Bakir, M.; Koc, A.; Akbulut, M.; Gercekcioglu, R.; Gunes, M.; Esitken, A. Simple sequence repeat-based assessment of genetic relationships among Prunus rootstocks. Genet. Mol. Res. 2010, 9, 2156–2165. [Google Scholar] [CrossRef] [PubMed]
- Anhalt, U.C.; Martini, K.; Ruehl, E.-H.; Forneck, A. Tracing Heterozygosity in the Vvlexp1 Locus in Grapevine by Sequenc-ing and High-resolution Melt Analysis. J. Am. Soc. Hort. Sci. 2013, 138, 120–124. [Google Scholar] [CrossRef]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–221. [Google Scholar] [CrossRef]
- Struss, D.; Ahmad, R.; Southwick, S.M.; Boritzki, M. Analysis of sweet cherry (Prunus avium L.) cultivars using SSR and AFLP markers. J. Am. Soc. Hortic. Sci. 2003, 128, 904–909. [Google Scholar] [CrossRef]
- Peace, C.; Bassil, N.; Main, D.; Ficklin, S.; Rosyara, U.R.; Stegmeir, T.; Sebolt, A.; Gilmore, B.; Lawley, C.; Mockler, T.C.; et al. Development and evaluation of a genome-wide 6K SNP array for diploid sweet cherry and tetraploid sour cherry. PLoS ONE 2012, 7, e48305. [Google Scholar] [CrossRef]
- Vaughan, S.P.; Bošković, R.I.; Gisbert-Climent, A.; Russell, K.; Tobutt, K.R. Characterisation of novel S-alleles from cherry (Prunus avium L.). Tree Genet. Genomes 2008, 4, 531–541. [Google Scholar] [CrossRef]
- Marchese, A.; Giovannini, D.; Leone, A.; Mafrica, R.; Palasciano, M.; Cantini, C.; Di Vaio, C.; de Salvador, F.R.; Giacalone, G.; Caruso, T.; et al. S-genotype identification, genetic diversity and structure analysis of Italian sweet cherry germplasm. Tree Genet. Genomes 2017, 13, 1–20. [Google Scholar] [CrossRef]
- Ercisli, S.; Agar, G.; Yildirim, N.; Karlidag, H.; Duralija, B.; Vokurka, A. Genetic diversity in wild sweet cherries (Prunus avium) in Turkey revealed by SSR markers. Genet. Mol. Res. 2011, 10, 1211–1219. [Google Scholar] [CrossRef]
- Xuan, H.; Wang, R.; Büchele, M.; Möller, O.; Hartmann, W. Microsatellite Markers (SSR) as a Tool to Assist in Identification of Sweet (Prunus avium) and Sour Cherry (Prunus cerasus). Acta Hortic. 2009. [Google Scholar] [CrossRef]
- Fernández i Martí, A.; Athanson, B.; Koepke, T.; Font I Forcada, C.; Dhingra, A.; Oraguzie, N. Genetic diversity and relat-edness of sweet cherry (Prunus avium L.) cultivars based on single nucleotide polymorphic markers. Front. Plant Sci. 2012, 3, 116. [Google Scholar] [CrossRef]
- Wünsch, A.; Hormaza, J.I. Molecular characterisation of sweet cherry (Prunus avium L.) genotypes using peach (Prunus per-sica (L.) Batsch) SSR sequences. Heredity 2002, 89, 56–63. [Google Scholar]
- Clausen, S.K.; Andersen, S.B.; Henriksen, K.; Toldam-Andersen, T.B.; Grout, B. Assessment of genetic diversity within sour cherry clones. Sci. Hortic. 2013, 164, 556–562. [Google Scholar] [CrossRef]
- Janes, J.K.; Steane, D.A.; Vaillancourt, R.E. What does population structure analysis reveal about the Pterostylis longifolia complex (Orchidaceae)? Ecol. Evol. 2012, 2, 2631–2644. [Google Scholar] [CrossRef]
- Struss, D.; Boritzki, M.; Glozer, K.; Southwick, S.M. Detection of genetic diversity among populations of sweet cherry (Prunus avium L.) by AFLPs. J. Hortic. Sci. Biotechnol. 2001, 76, 362–367. [Google Scholar] [CrossRef]
- Zhou, L.; Kappel, F.; Hampson, C.; Wiersma, P.A.; Bakkeren, G. Genetic Analysis and Discrimination of Sweet Cherry Cu-litvars and Selections Using Amplified Fragment Length Polymorphism Fingerprints. J. Am. Soc. Horti Cult. Sci. 2002, 127, 786–793. [Google Scholar] [CrossRef]
- Vanderzande, S.; Zheng, P.; Cai, L.; Barac, G.; Gasic, K.; Main, D.; Iezzoni, A.; Peace, C. The cherry 6+9K SNP array: A cost-effective improvement to the cherry 6K SNP array for genetic studies. Sci. Rep. 2020, 10, 7613. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S.; Rafalski, A. The comparison of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breed. 1996, 2, 225–238. [Google Scholar] [CrossRef]
- Xu, Q.; Wen, X.; Deng, X. A simple protocol for isolating genomic DNA from chestnut rose (Rosa roxburghii tratt) for RFLP and PCR analyses. Plant Mol. Biol. Rep. 2004, 22, 301–302. [Google Scholar] [CrossRef]
- Clarke, J.B.; Tobutt, K.R. Development and characterization of polymorphic microsatellites from Prunus avium ‘Napoleon’. Mol. Ecol. Notes 2003, 3, 578–580. [Google Scholar] [CrossRef]
- Vaughan, S.P.; Russell, K. Characterization of novel microsatellites and development of multiplex PCR for large-scale popu-lation studies in wild cherry, Prunus avium. Mol. Ecol. Notes 2004, 4, 429–431. [Google Scholar] [CrossRef]
- Testolin, R.; Marrazzo, T.; Cipriani, G.; Quarta, R.; Verde, I.; Dettori, M.T.; Pancaldi, M.; Sansavini, S. Microsatellite DNA in peach [Prunus persica (L.) Batsch] and its use in fingerprinting and testing the genetic origin of cultivars. Genome 2000, 43, 512–520. [Google Scholar] [CrossRef]
- Aranzana, M.J.; Garcia-Mas, J.; Carbo, J.; Arus, P. Development and variability of microsatellite markers in peach. Plant Breed. 2002, 121, 87–92. [Google Scholar] [CrossRef]
- Mnejja, M.; Garcia-Mas, J.; Howad, W.; Badenes, M.L.; Arus, P. Simple-sequence repeat (SSR) markers of Japanese plum (Prunus salicina Lindl.) are highly polymorphic and transferable to peach and almond. Mol. Ecol. Notes 2004, 4, 163–166. [Google Scholar] [CrossRef]
- Dirlewanger, E.; Cosson, P.; Tavaud, M.; Aranzana, M.J.; Poizat, C.; Zanetto, A.e.a. Development of microsatellite markers in peach [Prunus persica (L.) Batsch] and their use in genetic diversity analysis in peach and sweet cherry (Prunus avium L.). Theor. Appl. Genet. 2002, 105, 127–138. [Google Scholar] [CrossRef]
- Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M.; et al. AFLP: A new technique for DNA fingerprinting. Nucleic Acids Res. 1995, 23, 4407–4414. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mo-Lecular Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef]
- Hildebrand, C.E.; Torney, D.C.; Wagner, R.P. Informativeness of Polymorphic DNA Markers. Los Alamos Sci. 1992, 20, 100–102. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://r-project.org (accessed on 23 February 2021).
- Kamvar, Z.N.; Tabima, J.F.; Grünwald, N.J. Poppr: An R package for genetic analysis of populations with clonal, partially clonal, and/or sexual reproduction. PeerJ 2014, 2, e281. [Google Scholar] [CrossRef]
- Kamvar, Z.N.; Brooks, J.C.; Grünwald, N.J. Novel R tools for analysis of genome-wide population genetic data with empha-sis on clonality. Front. Genet. 2015, 6, 208. [Google Scholar] [CrossRef]
- Jombart, T. adegenet: A R package for the multivariate analysis of genetic markers. Bioinformatics 2008, 24, 1403–1405. [Google Scholar] [CrossRef]
- Jombart, T.; Ahmed, I. adegenet 1.3-1: New tools for the analysis of genome-wide SNP data. Bioinformatics 2011, 27, 3070–3071. [Google Scholar] [CrossRef] [PubMed]
- Jombart, T.; Collins, C. A Tutorial for Discriminant Analysis of Principal Components (DAPC) Using Adegenet 2.0. 0; Imperial College London, MRC Centre for Outbreak Analysis and Modelling: London, UK, 2015; pp. 1–43. [Google Scholar]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genet-ically structured populations. BMC Genet. 2010, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Wünsch, A.; Hormaza, J.I. Molecular evaluation of genetic diversity and S-allele composition of local Spanish sweet cherry (Prunus avium L.) cultivars. Genet. Resour. Crop Evol. 2004, 51, 635–641. [Google Scholar]
- Braun-Lüllemann, A.; Bannier, H.-J. Obstsortenwerk Alte Süßkirschsorten. Erstellt im Rahmen eines Modell- und Demonstrationsvorhabens im Bereich der Biologischen Vielfalt, Gefördert Durch die BLE. 2010. [Google Scholar]
- Modl, P.; Spornberger, A. Kirschen für den Naturnahen Hausgarten; Österreichischer Agrarverlag: Wien, Austria, 2009; ISBN 9783704023308. [Google Scholar]
- Barth, S.; Forneck, A.; Verzeletti, F.; Blaich, R.; Schumann, F. Genotypes and phenotypes of an ex situ Vitis vinifera ssp. syl-vestris (Gmel.) Beger germplasm collection from the Upper Rhine Valley. Genet. Resour. Crop Evol. 2009, 56, 1171–1181. [Google Scholar] [CrossRef]
- McKey, D.; Elias, M.; Pujol, B.; Duputie, A. The evolutionary ecology of clonally propagated domesticated plants. New Phytol. 2010, 186, 318–332. [Google Scholar] [CrossRef]
- Forneck, A. Plant Breeding: Clonality—A Concept for Stability and Variability During Vegetative Propagation. In Progress in Botany; Esser, K., Lüttge, U., Beyschlag, W., Murata, J., Eds.; Springer: Berlin/Heidelberg, Germany, 2005; pp. 164–183. ISBN 978-3-540-27043-0. [Google Scholar]
- Zhang, G.; Sebolt, A.M.; Sooriyapathirana, S.S.; Wang, D.; Bink, M.C.; Olmstead, J.W.; Iezzoni, A.F. Fruit size QTL analysis of an F1 population derived from a cross between a domesticated sweet cherry cultivar and a wild forest sweet cherry. Tree Genet. Genomes 2010, 6, 25–36. [Google Scholar] [CrossRef]
- Schüller, E.; Halbwirth, H.; Mikulic-Petkovsek, M.; Slatnar, A.; Veberic, R.; Forneck, A.; Stich, K.; Spornberger, A. High con-centrations of anthocyanins in genuine cherry-juice of old local Austrian Prunus avium varieties. Food Chem. 2015, 173, 935–942. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Variety Name or TN | TN in Collection | Sample Origin | Variety Origin | No of Trees of this Variety Name | Sample Set | Interesting Traits of Austrian Varieties | Reference |
|---|---|---|---|---|---|---|---|---|
| A60 | “Bigarreau Burlat” | 26113BSch | AT | FR | 2 | BOK | ||
| A59 | “Burlat VG“ | 26112BVG | AT | FR | 1 | BOK | ||
| A05 | “Butterkirsche” | TN34 | AT | AT | 5 | STB | taste, blushed type, early ripening | [12] |
| A06 | “Butterkirsche” | TN35 | AT | AT | 5 | STB | taste, blushed type, early ripening | [12] |
| A10 | “Butterkirsche” | TN44 | AT | AT | 5 | STB | taste, blushed type, early ripening | [12] |
| A13 | “Butterkirsche” | TN58 | AT | AT | 5 | STB | taste, blushed type, early ripening | [12] |
| A31 | “Butterkirsche” | TN165 | AT | AT | 5 | STB | taste, blushed type, early ripening | [12] |
| A63 | “Chelan” | US | CA | 1 | IS | |||
| A47 | “Donnerskircher Blaukirsche” | 2472Do | AT | AT | 1 | BOK | rich juice color, processed products | |
| A25 | “Early Burlat” | US | FR | 1 | IS | |||
| A01 | “F12/1” | GB | GB | 1 | IS | |||
| A62 | “Fruehe Kirsche Ubl” | 26325FKU | AT | AT | 1 | BOK | ||
| A52 | “Frueheste der Mark” | 10816FdM | AT | FR | 1 | BOK | early ripening | [8] |
| A39 | “Germersdorfer” | 2428Ge | AT | DE | 2 | BOK | ||
| A54 | “Germersdorfer” | 2426T1 | AT | DE | 2 | BOK | fruit size, fruit firmness | |
| A17 | “Goodnestone Black” | GB | GB | 1 | IS | |||
| A02 | “Grosse Schwarze Knorpelkirsche” | TN09 | AT | DE | 5 | STB | ||
| A03 | “Grosse Schwarze Knorpelkirsche” | TN29 | AT | DE | 5 | STB | ||
| A04 | “Grosse Schwarze Knorpelkirsche” | TN30 | AT | DE | 5 | STB | ||
| A15 | “Grosse Schwarze Knorpelkirsche” | TN87 | AT | DE | 5 | STB | ||
| A40 | “Grosse Schwarze Knorpelkirsche” | 2424GSK | AT | DE | 5 | BOK | ||
| A14 | “Hedelfinger Riesenkirsche” | TN60 | AT | DE | 5 | STB | ||
| A22 | “Hedelfinger Riesenkirsche” | TN128 | AT | DE | 5 | STB | ||
| A27 | “Hedelfinger Riesenkirsche” | TN146 | AT | DE | 5 | STB | ||
| A38 | “Hedelfinger Riesenkirsche” | 2444HR | AT | DE | 5 | BOK | ||
| A55 | “Hedelfinger Riesenkirsche” | 2437T2 | AT | DE | 5 | BOK | ||
| A23 | “Horitschoner Herzkirsche” | TN133 | AT | AT | 1 | STB | fruit size, fruit firmness | |
| A37 | “Horitschoner Herzkirsche” | 2452HH | AT | AT | 1 | BOK | fruit size, fruit firmness | |
| A51 | “Hybrid 222” | 10812Hy | AT | FR | 1 | BOK | ||
| A19 | “Jaboulay” | TN109 | AT | FR | 2 | STB | early ripening | |
| A43 | “Jaboulay” | 26120Sch | AT | FR | 2 | BOK | early ripening | |
| A44 | “Kritzendorfer Einsiedekirsche” Type I | 26225KrD1 | AT | AT | 1 | BOK | rich juice color, processed products | |
| A45 | “Kritzendorfer Einsiedekirsche” Type II | 2641KrD2 | AT | AT | 1 | BOK | rich juice color, processed products | |
| A46 | “Lambert” | 2456La | AT | US | 1 | BOK | ||
| A09 | “Lapins” | US | CA | 1 | IS | |||
| A16 | “Marzer Kirsche” | TN95 | AT | AT | 2 | STB | high fruit acidity, high ratio fruit weight/stone weight, early ripening | [8] |
| A36 | “Marzer Kirsche” | 2468Ma | AT | AT | 2 | BOK | high fruit acidity, high ratio fruit weight/stone weight, early ripening | [8] |
| A42 | “Melker Riesenkirsche” | 2431MRK | AT | AT | 1 | BOK | fruit size, fruit firmness | [8] |
| A33 | “Napoleon” | GB | DE | 1 | IS | |||
| A58 | “Noble” | GB | GB | 1 | IS | |||
| A49 | “Noire de Meched” | GB | IR | 1 | IS | |||
| A41 | “NY54” | US | US | 1 | IS | |||
| A26 | “Saemling von Sauerbrunn” | TN143 | AT | AT | 1 | STB | tolerance to fruit cracking | [8] |
| A61 | “Saemling von Sauerbrunn” | 26122LaSt | AT | AT | 2 | BOK | tolerance to fruit cracking | [8] |
| A48 | “Sarga Dragan” | 1071SD | AT | HU | 1 | BOK | ||
| A56 | “Schneiders Spaete Knorpelkirsche” | US | DE | 2 | IS | |||
| A53 | “Schneiders Spaete Knorpelkirsche” | 2422SSK | AT | DE | 2 | BOK | ||
| A57 | “Stella Spur” | 2453StP | AT | CA | 1 | BOK | ||
| A50 | “Tavriczskai” | 1082Ta | AT | HU | 1 | BOK | ||
| A20 | TN110 | TN110 | AT | AT | 1 | STB | early ripening, mechanical harvest | [12] |
| A21 | TN120 | TN120 | AT | AT | 1 | STB | early ripening | [12] |
| A24 | TN142 | TN142 | AT | AT | 1 | STB | early ripening, mechanical harvest | [12] |
| A28 | TN155 | TN155 | AT | AT | 1 | STB | ||
| A29 | TN157 | TN157 | AT | AT | 1 | STB | ||
| A30 | TN163 | TN163 | AT | AT | 1 | STB | mechanical harvest | [12] |
| A32 | TN175 | TN175 | AT | AT | 1 | STB | fruit size, fruit firmness | |
| A34 | TN177 | TN177 | AT | AT | 1 | STB | ||
| A07 | TN36 | TN36 | AT | AT | 1 | STB | ||
| A08 | TN39 | TN39 | AT | AT | 1 | STB | ||
| A11 | TN46 | TN46 | AT | AT | 1 | STB | mechanical harvest | [12] |
| A12 | TN52 | TN52 | AT | AT | 1 | STB | early ripening | [12] |
| A18 | TN96 | TN96 | AT | AT | 1 | STB | taste, early ripening | [12] |
| A35 | “Ulster” | US | US | 1 | IS |
| Code, TN | Variety Name or TN | Sample Origin | Variety Origin | Sample Set | SSR-Analysis (Table 1) |
|---|---|---|---|---|---|
| 2451F12_1 | “F12/1” | GB | GB | IS | Yes |
| 2425T1 | “Germersdorfer” | AT | DE | BOK | No |
| 2428Ge | “Germersdorfer” | AT | DE | BOK | Yes |
| Ge_alt_VG | “Germersdorfer” | AT | DE | BOK | No |
| B04 | “Germersdorfer” | AT | DE | Leitha | No |
| P10 | “Germersdorfer” | AT | DE | Leitha | No |
| S20 | “Germersdorfer” | AT | DE | Scharten | No |
| S26 | “Germersdorfer” | AT | DE | Scharten | No |
| 2452HH | “Horitschoner Herzkirsche” | AT | AT | BOK | Yes |
| D10 | “Horitschoner Herzkirsche” | AT | AT | Leitha | No |
| P12 | “Horitschoner Herzkirsche” | AT | AT | Leitha | No |
| 2431MRK | “Melker Riesenkirsche” | AT | AT | BOK | Yes |
| P09 | “Melker Riesenkirsche” | AT | AT | Leitha | No |
| Noble 2013, Noble 2015 | “Noble” | GB | GB | IS | Yes |
| N.d.M. 2013, N.d.M.2015 | “Noire de Meched” | GB | IR | IS | Yes |
| K02 | “Rainkirsche” | AT | AT | Scharten | No |
| 2422SSK | “Schneiders Spaete Knorpelkirsche” | AT | DE | BOK | Yes |
| SSK_alt_VG | “Schneiders Spaete Knorpelkirsche” | AT | DE | BOK | No |
| SSKB | “Schneiders Spaete Knorpelkirsche” | AT | DE | Scharten | No |
| Multiplex | Primer | nmol in 13 µL PCR Reaction | Dye | Linkage Group | Reference |
|---|---|---|---|---|---|
| MP1 | EMPa002 | 2.50 | PET | LG1 | [34] |
| EMPaS12 | 1.75 | FAM | LG6 | [35] | |
| EMPaS02 | 3.75 | NED | LG3 | [35] | |
| UDP98-412 | 1.25 | VIC | LG6 | [36] | |
| CPPCT006 | 2.50 | PET | LG8 | [37] | |
| EMPaS01 | 5.00 | FAM | LG6 | [35] | |
| EMPa017 | 5.00 | NED | LG2 | [34] | |
| CPPCT022 | 2.50 | VIC | LG7 | [37] | |
| MP2 | CPSCT038 | 2.50 | NED | LG2 | [38] |
| BPPCT034 | 3.00 | PET | LG2 | [39] | |
| BPPCT037 | 1.75 | FAM | LG5 | [39] | |
| EMPaS10 | 1.75 | VIC | LG4 | [35] |
| Locus | N | Ho | He | PIC | PA | PA-MLGs |
|---|---|---|---|---|---|---|
| BPPCT037 | 8 | 0.89 | 0.81 | 0.78 | 2 | 2 |
| CPPCT006 | 8 | 0.89 | 0.77 | 0.73 | 2 | 2 |
| CPPCT022 | 8 | 0.51 | 0.59 | 0.56 | 4 | 6 |
| CPSCT038 | 3 | 0.58 | 0.61 | 0.54 | ||
| EMPa002 | 3 | 0.75 | 0.54 | 0.45 | ||
| EMPa017 | 5 | 0.06 | 0.23 | 0.22 | 1 | 1 |
| EMPaS01 | 5 | 0.73 | 0.70 | 0.66 | ||
| EMPaS02 | 7 | 0.84 | 0.72 | 0.68 | 1 | 1 |
| EMPaS10 | 8 | 0.49 | 0.60 | 0.53 | 4 | 5 |
| EMPaS12 | 5 | 0.83 | 0.75 | 0.71 | ||
| UDP98-412 | 9 | 0.71 | 0.70 | 0.67 | 1 | 1 |
| Total | 69 | |||||
| Average | 6.3 | 0.66 | 0.64 | 0.59 |
| Cluster | Variety | Sample Set | Accessions with Same Genotype | Code |
|---|---|---|---|---|
| 1 | “F12/1” | IS | A01 | |
| 1 | “Butterkirsche” | STB | A06, A07, A10, A13, A31 | A05 |
| 1 | TN110 | STB | A20 | |
| 1 | TN163 | STB | A30 | |
| 1 | “NY54” | IS | A41 | |
| 1 | “Kritzendorfer Einsiedekirsche” “Type I” | BOK | A45 | A44 |
| 1 | “Donnerskircher Blaukirsche” | BOK | A47 | |
| 1 | “Sämling von Sauerbrunn” | BOK | A61 | |
| 1 | “Frühe Kirsche Ubl” | BOK | A62 | |
| 2 | “Große Schwarze Knorpelkirsche” | STB | A02 | |
| 2 | “Große Schwarze Knorpelkirsche” | STB | A04 | A03 |
| 2 | “Lapins” | IS | A09 | |
| 2 | TN46 | STB | A11 | |
| 2 | TN52 | STB | A12 | |
| 2 | “Hedelfinger Riesenkirsche” | STB | A22 | A14 |
| 2 | “Große Schwarze Knorpelkirsche” | STB | A15 | |
| 2 | “Goodnestone Black” | IS | A17 | |
| 2 | “Sämling von Sauerbrunn” | STB | A26 | |
| 2 | “Hedelfinger Riesenkirsche” | STB | A27 | |
| 2 | “Napoleon” | IS | A33 | |
| 2 | “Ulster” | IS | A35 | |
| 2 | “Hedelfinger Riesenkirsche” | BOK | A38 | |
| 2 | “Große Schwarze Knorpelkirsche” | BOK | A40 | |
| 2 | “Lambert” | BOK | A57 | A46 |
| 2 | “Sarga Dragan” | BOK | A48 | |
| 2 | “Tavriczskai” | BOK | A50 | |
| 2 | “Tscholl 2” | BOK | A55 | |
| 3 | TN39 | STB | A23, A28, A29, A32, A34, A37, A39, A42, A49, A53, A54, A56 | A08 |
| 3 | “Marzer Kirsche” | STB | A16 | |
| 3 | TN96 | STB | A21, A24 | A18 |
| 3 | “Jaboulay” | STB | A43 | A19 |
| 3 | “Early Burlat” | IS | A25 | |
| 3 | “Marzer Kirsche” | BOK | A36 | |
| 3 | “Hybrid 222” | BOK | A59 | A51 |
| 3 | “Früheste der Mark” | BOK | A52 | |
| 3 | “Noble” | IS | A58 | |
| 3 | “Bigarreau Burlat” | BOK | A60 | |
| 3 | “Chelan” | IS | A63 |
| Pop | N | MLG | eMLG | SE | lambda | Hexp | Ia | rbarD |
|---|---|---|---|---|---|---|---|---|
| 1 | 9 | 9 | 9 | 0 | 0.889 | 0.630 | 1.035 | 0.1075 |
| 2 | 18 | 18 | 10 | 5.43 × 10−7 | 0.944 | 0.595 | 0.316 | 0.0331 |
| 3 | 11 | 11 | 10 | 0 | 0.909 | 0.614 | 0.634 | 0.0654 |
| Total | 38 | 38 | 10 | 1.72 × 10−6 | 0.974 | 0.665 | 0.653 | 0.0662 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Schüller, E.; Fernández, F.F.; Antanaviciute, L.; Anhalt-Brüderl, U.; Spornberger, A.; Forneck, A. Autochthonous Austrian Varieties of Prunus avium L. Represent a Regional Gene Pool, Assessed Using SSR and AFLP Markers. Genes 2021, 12, 322. https://doi.org/10.3390/genes12030322
Schüller E, Fernández FF, Antanaviciute L, Anhalt-Brüderl U, Spornberger A, Forneck A. Autochthonous Austrian Varieties of Prunus avium L. Represent a Regional Gene Pool, Assessed Using SSR and AFLP Markers. Genes. 2021; 12(3):322. https://doi.org/10.3390/genes12030322
Chicago/Turabian StyleSchüller, Elisabeth, Felicidad Fernández Fernández, Laima Antanaviciute, Ulrike Anhalt-Brüderl, Andreas Spornberger, and Astrid Forneck. 2021. "Autochthonous Austrian Varieties of Prunus avium L. Represent a Regional Gene Pool, Assessed Using SSR and AFLP Markers" Genes 12, no. 3: 322. https://doi.org/10.3390/genes12030322
APA StyleSchüller, E., Fernández, F. F., Antanaviciute, L., Anhalt-Brüderl, U., Spornberger, A., & Forneck, A. (2021). Autochthonous Austrian Varieties of Prunus avium L. Represent a Regional Gene Pool, Assessed Using SSR and AFLP Markers. Genes, 12(3), 322. https://doi.org/10.3390/genes12030322