
BPA and BPS Affect Connexin 37 in Bovine Cumulus Cells

 , ,
, , 
Abstract
1. Introduction
2. Materials and Methods
2.1. Oocyte Collection and Maturation
2.2. In Vitro Fertilization (IVF)
2.3. Cell Culture and Treatment
2.4. RNA Extraction
2.5. Quantitative Real-Time Polymerase Chain Reaction (qPCR)
2.6. Western Blotting
2.7. Confocal Microscopy
2.8. Statistical Analysis
3. Results
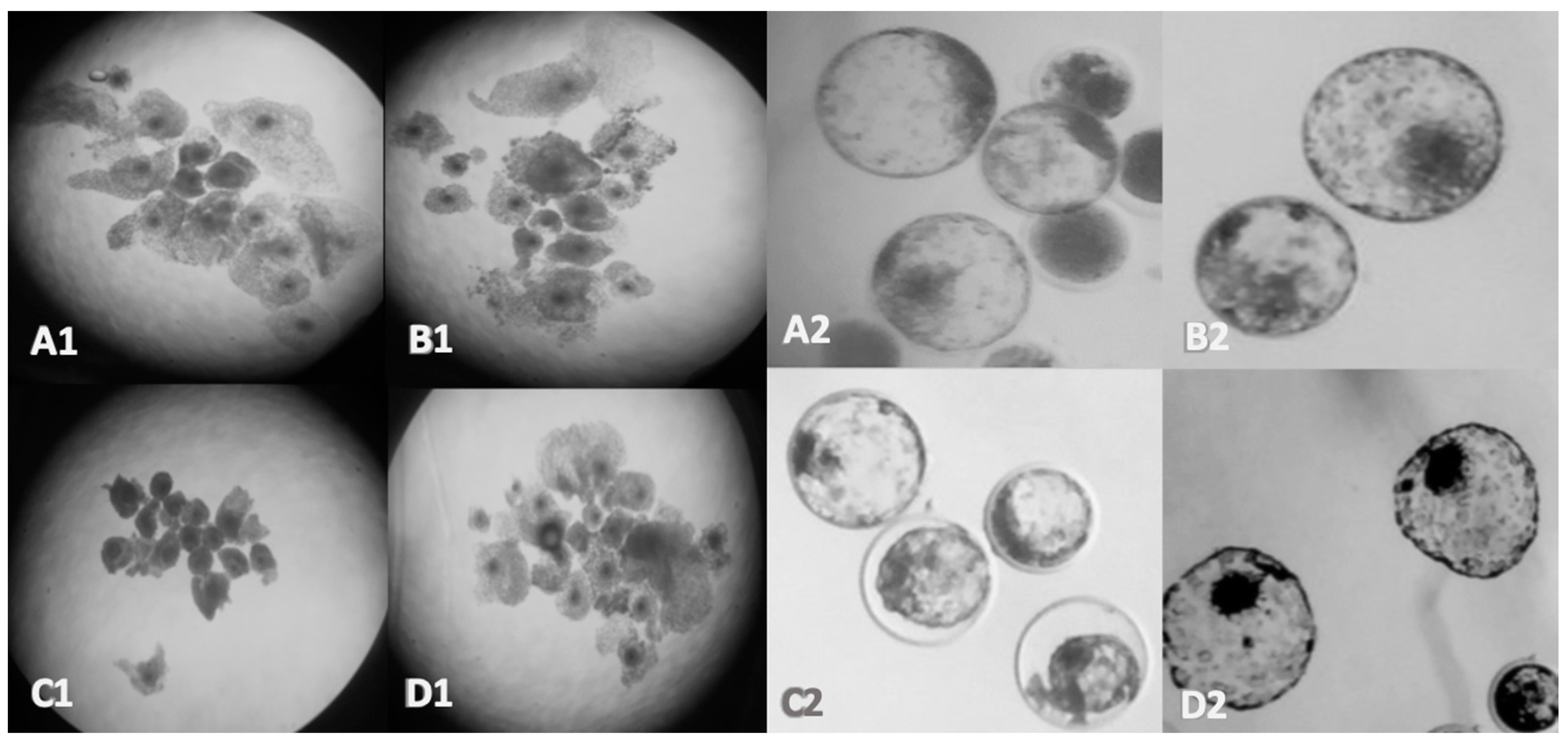
3.1. Oocyte and Embryo Morphology
3.2. Cell Culture Treatments
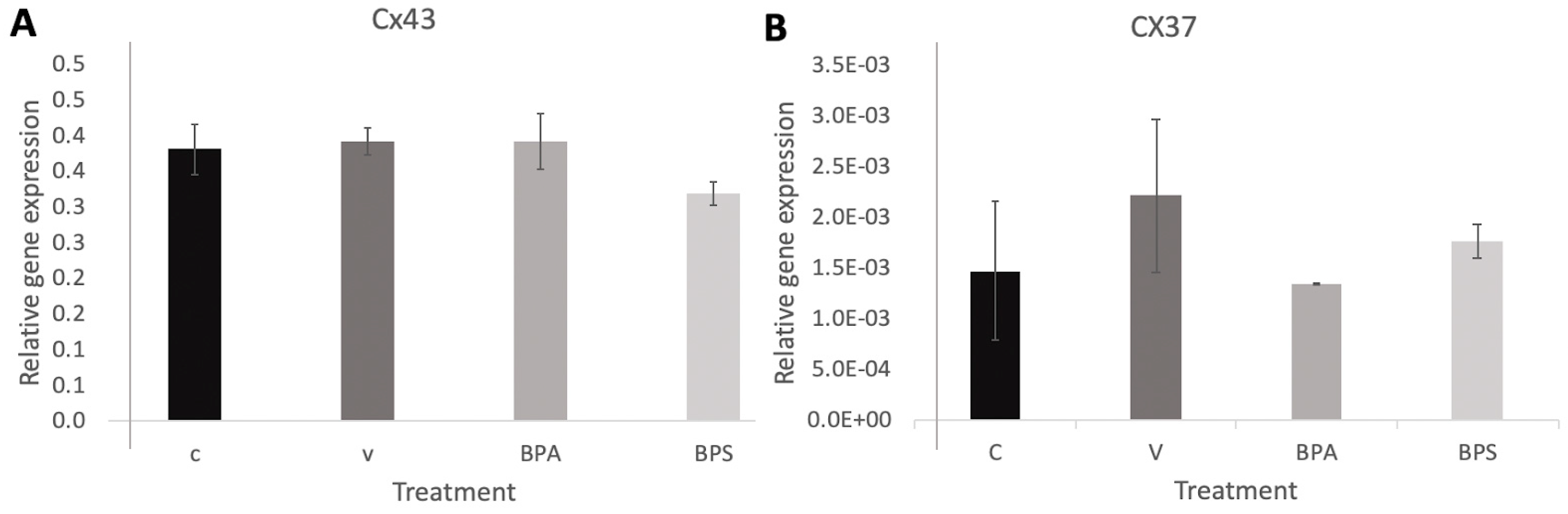
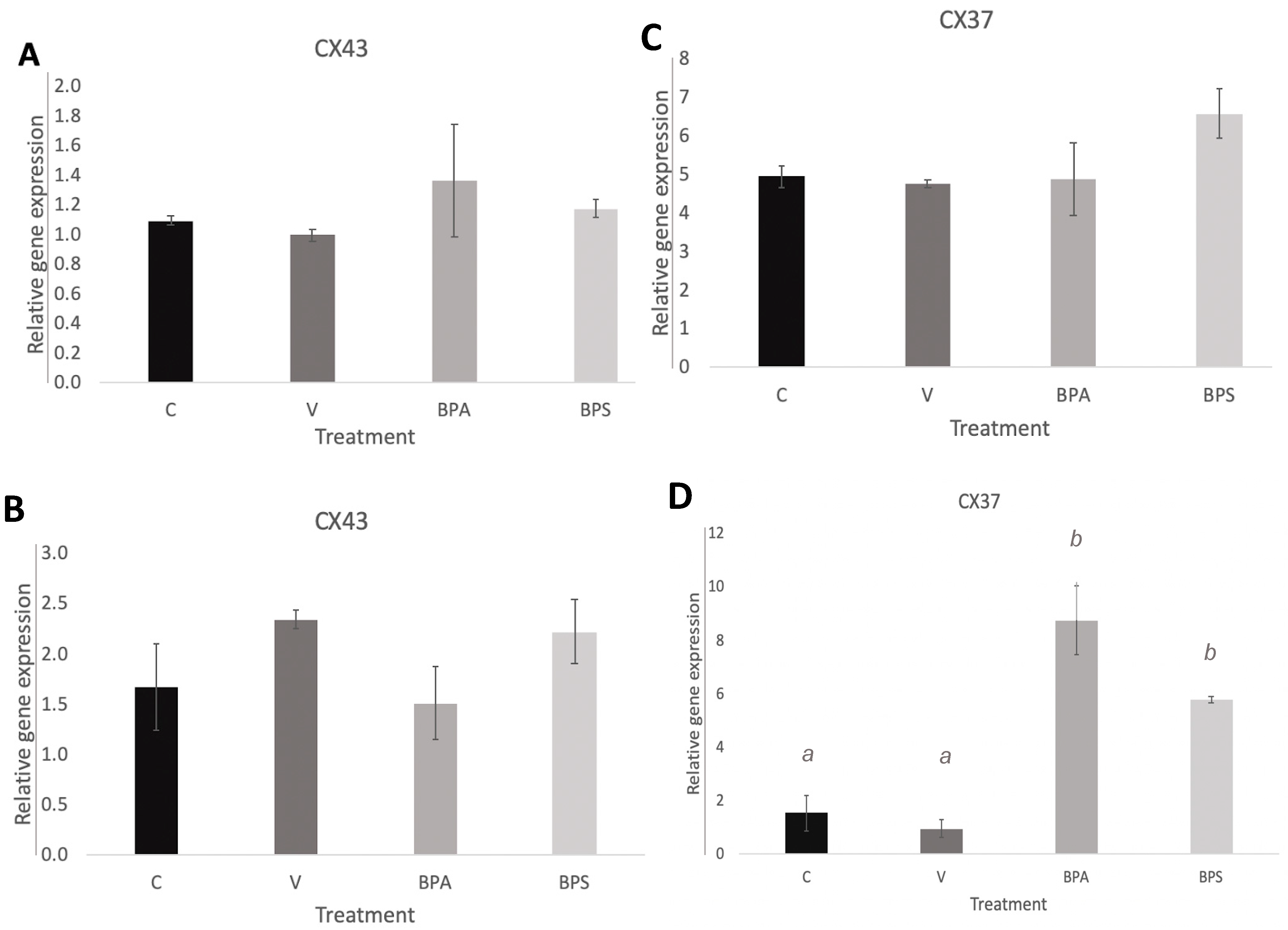
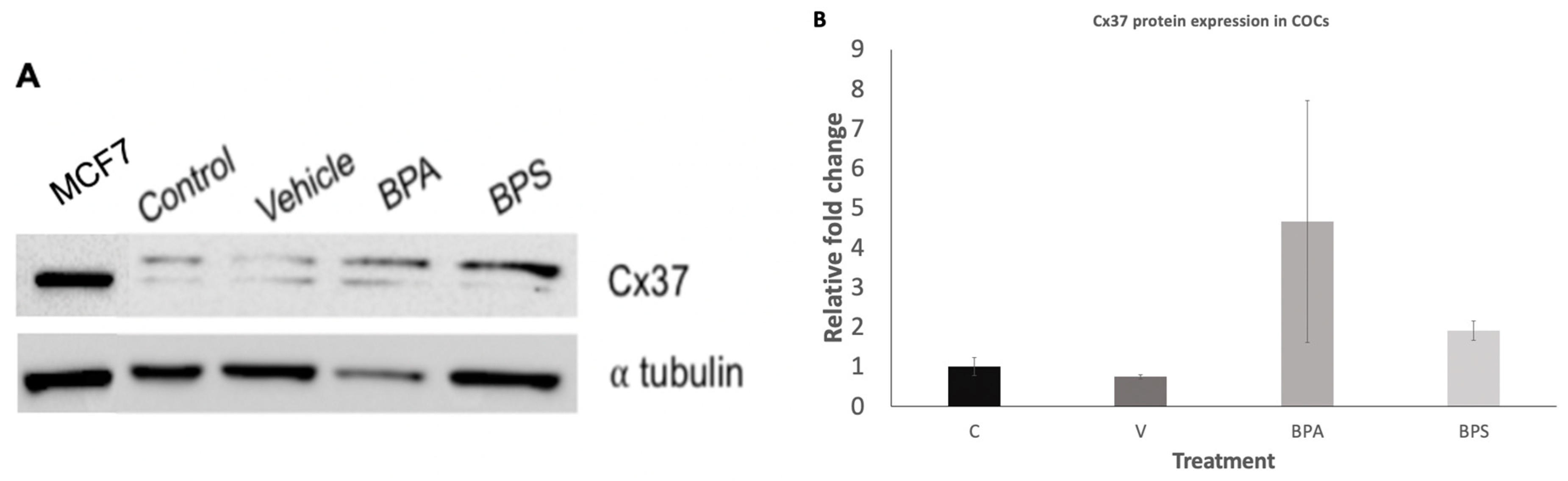
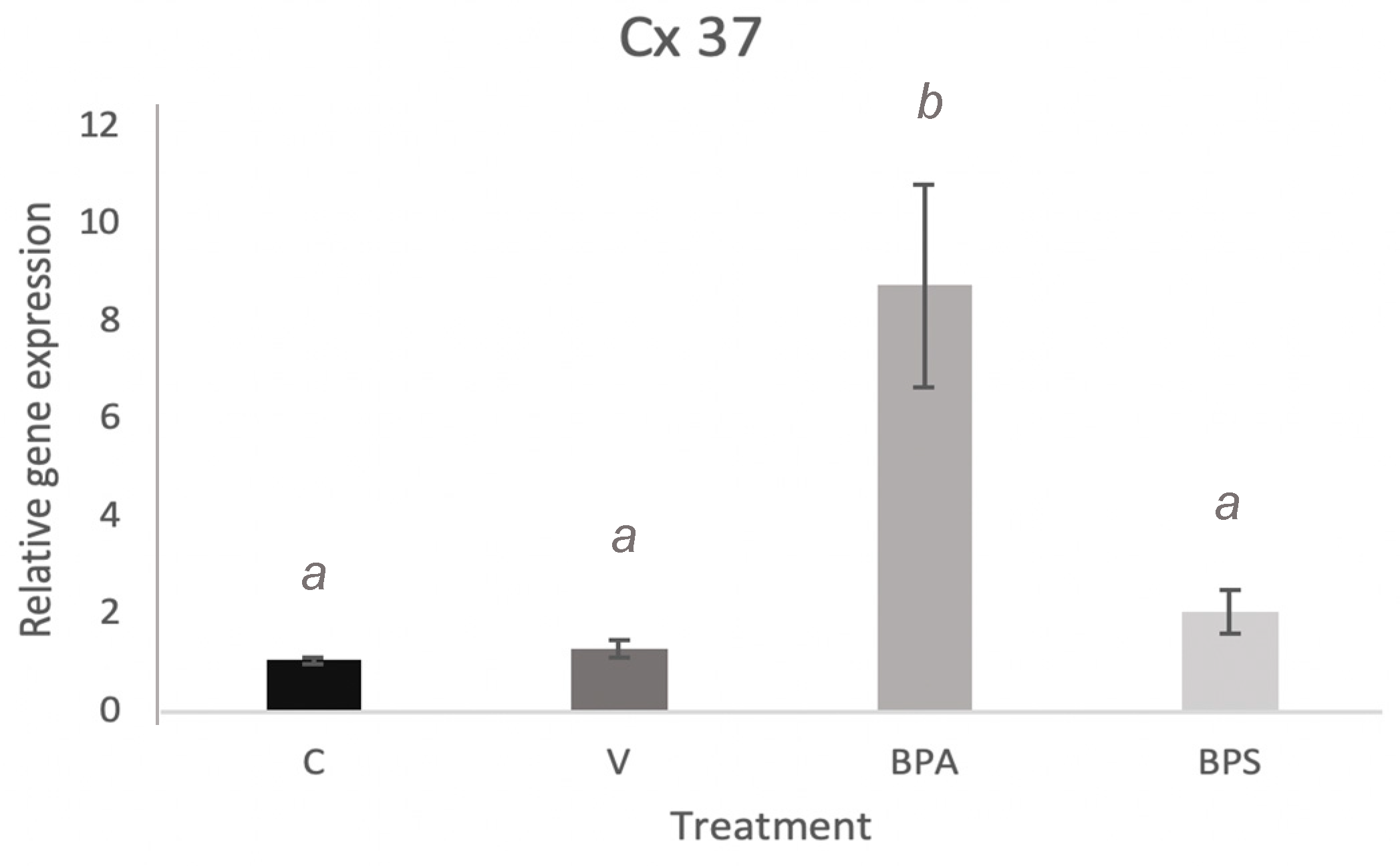
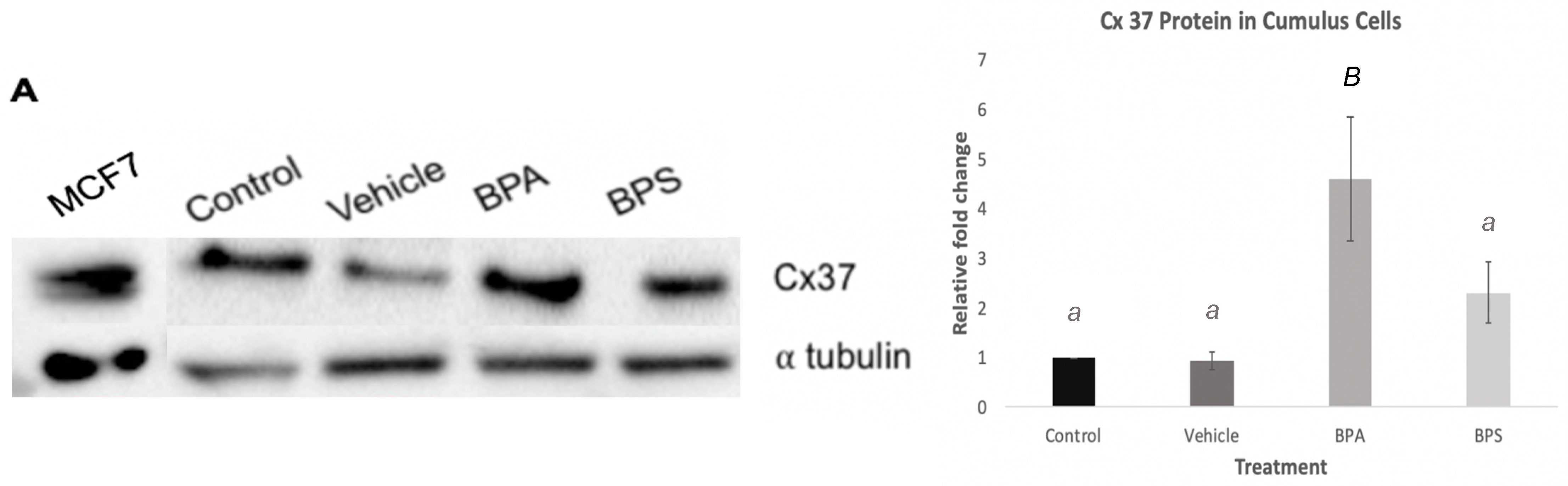
3.3. Cx43 and Cx37 Expression in COCs, Oocytes, and Cumulus Cells
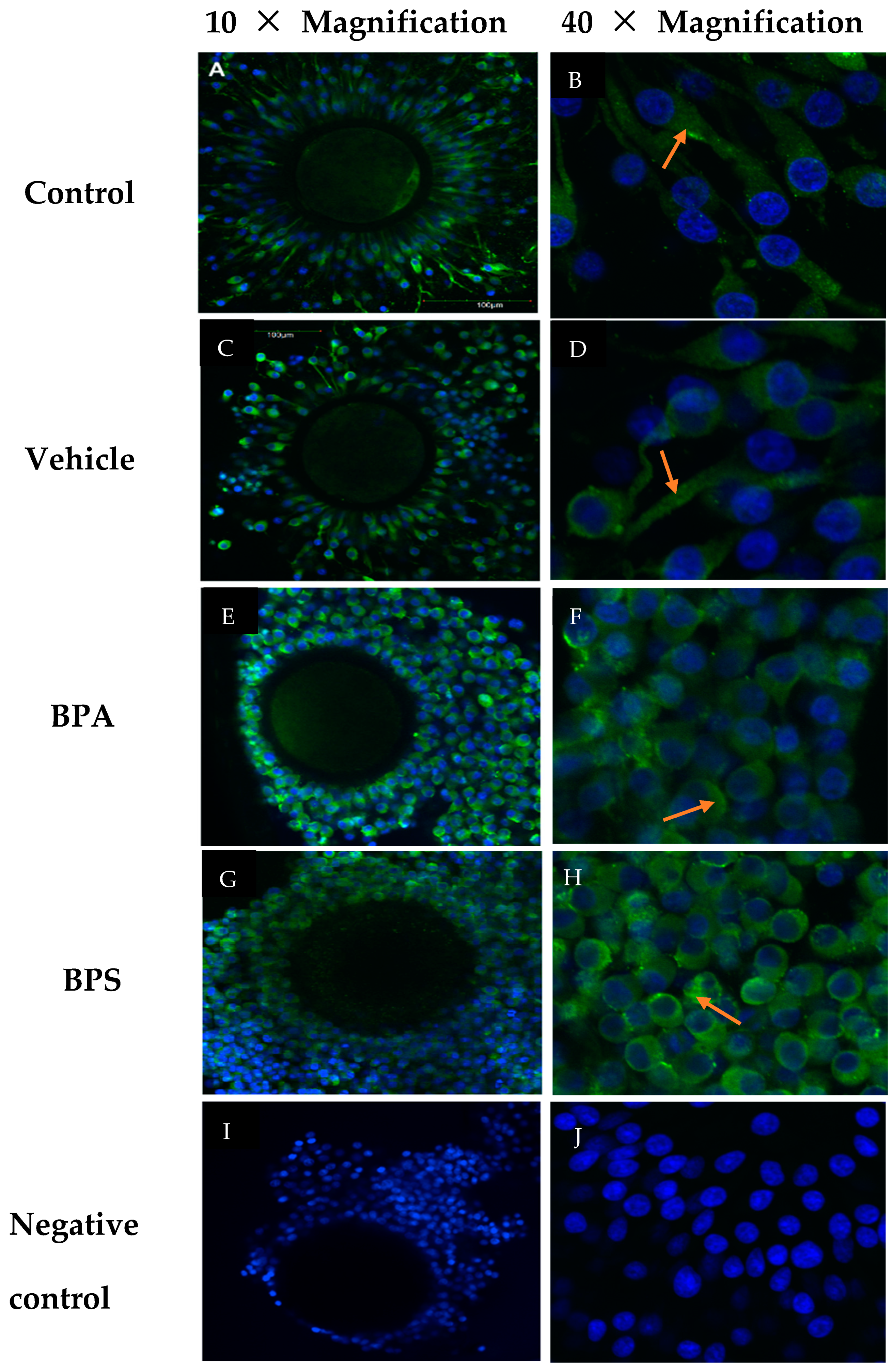
3.4. Cx37 Protein Localization in COCs, Oocytes, and Cumulus Cells
3.5. Cx37 Expression in In Vitro Cultured Cumulus Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Eichenlaub-Ritter, U.; Pacchierotti, F. Bisphenol A Effects on Mammalian Oogenesis and Epigenetic Integrity of Oocytes: A Case Study Exploring Risks of Endocrine Disrupting Chemicals. BioMed Res. Int. 2015, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rubin, B.S. Bisphenol A: An endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011, 127, 27–34. [Google Scholar] [CrossRef] [PubMed]
- Murata, M.; Kang, J. Bisphenol A (BPA) and cell signaling pathways. Biotechnol. Adv. 2018, 36, 311–327. [Google Scholar] [CrossRef] [PubMed]
- Acuña-Hernández, D.G.; Arreola-Mendoza, L.; Santacruz-Márquez, R.; García-Zepeda, S.P.; Parra-Forero, L.Y.; Olivares-Reyes, J.; Hernández-Ochoa, I. Bisphenol A alters oocyte maturation by prematurely closing gap junctions in the cumulus cell-oocyte complex. Toxicol. Appl. Pharmacol. 2018, 344, 13–22. [Google Scholar] [CrossRef]
- Petro, E.M.; Leroy, J.L.; Covaci, A.; Fransen, E.; De Neubourg, D.; Dirtu, A.C.; Bols, P.E. Endocrine-disrupting chemicals in human follicular fluid impair in vitro oocyte developmental competence. Hum. Reprod. 2012, 27, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Rhind, S.M. Endocrine disrupting compounds and farm animals: Their properties, actions and routes of exposure. Domest. Anim. Endocrinol. 2002, 23, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Allard, P. Chapter 50—Bisphenol A. In Reproductive and Developmental Toxicology; Elsevier: Amsterdam, The Netherlands, 2011; pp. 673–686. [Google Scholar] [CrossRef]
- Moore-Ambriz, T.; Acuña-Hernández, D.; Ramos-Robles, B.; Sánchez-Gutiérrez, M.; Santacruz-Márquez, R.; Sierra-Santoyo, A. Exposure to bisphenol A in young adult mice does not alter ovulation but does alter the fertilization ability of oocytes. Toxicol. Appl. Pharmacol. 2015, 289, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Ferris, J.; Favetta, L.A.; King, W.A. Bisphenol A Exposure during Oocyte Maturation in vitro Results in Spindle Abnormalities and Chromosome Misalignment in Bos taurus. Cytogenet. Genome Res. 2015, 145, 50–58. [Google Scholar] [CrossRef]
- Ferris, J.; Mahboubi, K.; MacLusky, N.; King, W.A.; Favetta, L.A. BPA exposure during in vitro oocyte maturation results in dose-dependent alterations to embryo development rates, apoptosis rate, sex ratio and gene expression. Reprod. Toxicol. 2016, 59, 128–138. [Google Scholar] [CrossRef]
- Saleh, A.; Favetta, L. 159 Effect of bisphenol A and bisphenol S on AMH and AMHR mRNA expression during in vitro bovine oocyte maturation and early embryo development. Reprod. Fertil. Dev. 2018, 31, 204–205. [Google Scholar] [CrossRef]
- Téteau, O.; Jaubert, M.; Desmarchais, A.; Papillier, P.; Binet, A.; Maillard, V.; Elis, S. Bisphenol A and S impaired ovine granulosa cell steroidogenesis. Reproduction 2020, 159, 571–583. [Google Scholar] [CrossRef] [PubMed]
- Vandenberg, L.N.; Maffini, M.V.; Sonnenschein, C.; Rubin, B.S.; Soto, A.M. Bisphenol-A and the Great Divide: A Review of Controversies in the Field of Endocrine Disruption. Endocr. Rev. 2009, 30, 75–95. [Google Scholar] [CrossRef] [PubMed]
- Rochester, J.R.; Bolden, A.L. Bisphenol S and F: A Systematic Review and Comparison of the Hormonal Activity of Bisphenol A Substitutes. Environ. Health Perspect. 2015, 123, 643–650. [Google Scholar] [CrossRef] [PubMed]
- Liao, C.; Liu, F.; Alomirah, H.; Loi, V.; Mohd, M.; Moon, H. Bisphenol S in Urine from the United States and Seven Asian Countries: Occurrence and Human Exposures. Environ. Sci. Technol. 2012, 46, 6860–6866. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Jahan, S.; Ain, Q.; Shaheen, G.; Ahsan, N. Effect of bisphenol S exposure on male reproductive system of rats: A histological and biochemical study. Chemosphere 2016, 152, 383–391. [Google Scholar] [CrossRef]
- Grignard, E.; Lapenna, S.; Bremer, S. Weak estrogenic transcriptional activities of Bisphenol A and Bisphenol S. Toxicol. In Vitro 2012, 26, 727–731. [Google Scholar] [CrossRef]
- Qiu, W.; Yang, M.; Liu, S.; Lei, P.; Hu, L.; Chen, B.; Wang, K. Toxic Effects of Bisphenol S Showing Immunomodulation in Fish Macrophages. Environ. Sci. Technol. 2018, 52, 831–838. [Google Scholar] [CrossRef]
- Zhang, T.; Xue, J.; Gao, C.Z.; Qiu, R.L.; Li, Y.X.; Li, X.; Kannan, K. Urinary concentrations of bisphenols and their association with biomarkers of oxidative stress in people living near E-waste recycling facilities in china. Environ. Sci. Technol. 2016, 50, 4045–4053. [Google Scholar] [CrossRef]
- Žalmanová, T.; Hošková, K.; Nevoral, J.; Adámková, K.; Kott, T.; Šulc, M.; Kotíková, Z.; Prokešová, Š.; Jílek, F.; Králíčková, M.; et al. Bisphenol S negatively affects the meiotic maturation of pig oocytes. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Desmarchais, A.; Téteau, O.; Papillier, P.; Jaubert, M.; Druart, X.; Binet, A.; Maillard, V.; Elis, S. Bisphenol S Impaired In Vitro Ovine Early Developmental Oocyte Competence. Int. J. Mol. Sci. 2020, 21, 1238. [Google Scholar] [CrossRef]
- Amar, S.; Binet, A.; Téteau, O.; Desmarchais, A.; Papillier, P.; Lacroix, M.Z.; Maillard, V.; Guérif, F.; Elis, S. Bisphenol S Impaired Human Granulosa Cell Steroidogenesis in Vitro. Int. J. Mol. Sci. 2020, 21, 1821. [Google Scholar] [CrossRef]
- Ben-Jonathan, N.; Hugo, E.R. Bisphenols Come in Different Flavors: Is “S” Better Than “A”? Endocrinology 2016, 157, 1321–1323. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Almeida, S.; Raposo, A.; Almeida-González, M.; Carrascosa, C. Bisphenol A: Food Exposure and Impact on Human Health. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1503–1517. [Google Scholar] [CrossRef]
- Apau, J.; Acheampong, A.; Adua, E. Exposure to bisphenol A, bisphenol F, and bisphenol S can result in obesity in human body. Cogent Chem. 2018, 4, 1506601. [Google Scholar] [CrossRef]
- Ghayda, R.A.; Williams, P.L.; Chavarro, J.E.; Ford, J.B.; Souter, I.; Calafat, A.M.; Hauser, R.; Mínguez-Alarcón, L. Urinary bisphenol S concentrations: Potential predictors of and associations with semen quality parameters among men attending a fertility center. Environ. Int. 2019, 131, 105050. [Google Scholar] [CrossRef] [PubMed]
- Uyar, A.; Torrealday, S.; Seli, E. Cumulus and granulosa cell markers of oocyte and embryo quality. Fertil. Steril. 2013, 99, 979–997. [Google Scholar] [CrossRef] [PubMed]
- Davidson, A. The Effect of Bisphenol A Exposure during Bovine Oocyte Maturation on Connexin 37 and Connexin 43 Gene Expression. Ph.D. Thesis, The University of Guelph, Guelph, ON, Canada, 2016. [Google Scholar]
- Thomas, R.E.; Armstrong, D.T.; Gilchrist, R.B. Bovine Cumulus Cell-Oocyte Gap Junctional Communication During In Vitro Maturation in Response to Manipulation of Cell-Specific Cyclic Adenosine 3′,5′-Monophosophate Levels1. Biol. Reprod. 2004, 70, 548–556. [Google Scholar] [CrossRef] [PubMed]
- Veitch, G.I.; Gittens, J.E.I.; Shao, Q.; Laird, D.W.; Kidder, G.M. Selective assembly of connexin37 into heterocellular gap junctions at the oocyte/granulosa cell interface. J. Cell Sci. 2004, 117, 2699–2707. [Google Scholar] [CrossRef]
- Houghton, F.D. Role of gap junctions during early embryo development. Reproduction 2005, 129, 129–135. [Google Scholar] [CrossRef]
- Gittens, J.E.I.; Kidder, G.M. Differential contributions of connexin37 and connexin43 to oogenesis revealed in chimeric reaggregated mouse ovaries. J. Cell Sci. 2005, 118, 5071–5078. [Google Scholar] [CrossRef]
- Silva, A.M.; Bruno, J.B.; Lima, L.F.; Sá, N.A.; Lunardi, F.O.; Ferreira, A.C.; Rodrigues, A.P. Connexin 37 and 43 gene and protein expression and developmental competence of isolated ovine secondary follicles cultured in vitro after vitrification of ovarian tissue. Theriogenology 2016, 85, 1457–1467. [Google Scholar] [CrossRef] [PubMed]
- Ke, Q.; Li, L.; Cai, B.; Liu, C.; Yang, Y.; Gao, Y.; Peng Xiang, A. Connexin 43 is involved in the generation of human-induced pluripotent stem cells. Hum. Mol. Genet. 2013, 22, 2221–2223. [Google Scholar] [CrossRef] [PubMed]
- Kidder, G.M.; Vanderhyden, B.C. Bidirectional communication between oocytes and follicle cells: Ensuring oocyte developmental competence. Can. J. Physiol. Pharmacol. 2010, 88, 399–413. [Google Scholar] [CrossRef] [PubMed]
- Winterhager, E.; Kidder, G.M. Gap junction connexins in female reproductive organs: Implications for women’s reproductive health. Hum. Reprod. Update 2015, 21, 340–352. [Google Scholar] [CrossRef]
- Peretz, J.; Craig, Z.R.; Flaws, J.A. Bisphenol A Inhibits Follicle Growth and Induces Atresia in Cultured Mouse Antral Follicles Independently of the Genomic Estrogenic Pathway. Biol. Reprod. 2012, 87, 63. [Google Scholar] [CrossRef]
- Sabry, R.; Saleh, A.C.; Stalker, L.; Lamarre, J.; Favetta, L.A. Effects of bisphenol A and bisphenol S on microRNA expression during bovine (Bos taurus) oocyte maturation and early embryo development. Reprod. Toxicol. 2020, 99, 96–108. [Google Scholar] [CrossRef]
- Hamilton, C.K.; Révay, T.; Domander, R.; Favetta, L.A.; King, W.A. A Large Expansion of the HSFY Gene Family in Cattle Shows Dispersion across Yq and Testis-Specific Expression. PLoS ONE 2011, 6, e17790. [Google Scholar] [CrossRef] [PubMed]
- Tesfaye, D.; Lonergan, P.; Hoelker, M.; Rings, F.; Nganvongpanit, K.; Havlicek, V.; Besenfelder, U.; Jennen, D.; Tholen, E.; Schellander, K. Suppression of connexin 43 and E-cadherin transcripts in in vitro derived bovine embryos following culture in vitro or in vivo in the homologous bovine oviduct. Mol. Reprod. Dev. 2007, 74, 978–988. [Google Scholar] [CrossRef] [PubMed]
- Bo, G.A.; Mapletoft, R.J. Evaluation and classification of bovine embryos. Anim. Reprod. 2013, 10, 344–348. Available online: http://www.animal-reproduction.org/journal/animreprod/article/5b5a604cf7783717068b46a2 (accessed on 11 January 2021).
- Lourenço, B.; Sousa, A.P.; Almeida-Santos, T.; Ramalho-Santos, J. Relation of Cumulus Cell Status with Single Oocyte Maturity, Fertilization Capability and Patient Age. J. Reprod. Infertil. 2014, 15, 15–21. Available online: https://pubmed.ncbi.nlm.nih.gov/24696155 (accessed on 7 January 2021).
- Gottschall, P.E.; Katsuura, G.; Arimura, A. Interleukin-1 suppresses follicle-stimulating hormone-induced estradiol secretion from cultured ovarian granulosa cells. J. Reprod. Immunol. 1989, 15, 281–290. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, W. Methods for Evaluation of Ovarian Granulosa Cells with Exposure to Nanoparticles. Methods Mol. Biol. 2019, 1894, 73–81. [Google Scholar] [CrossRef]
- Chermuła, B.; Kranc, W.; Jopek, K.; Budna-Tukan, J.; Hutchings, G.; Dompe, C.; Moncrieff, L.; Janowicz, K.; Józkowiak, M.; Jeseta, M.; et al. Human Cumulus Cells in Long-Term In Vitro Culture Reflect Differential Expression Profile of Genes Responsible for Planned Cell Death and Aging-A Study of New Molecular Markers. Cells 2020, 9, 1265. [Google Scholar] [CrossRef]
- Macaulay, A.D.; Gilbert, I.; Scantland, S.; Fournier, E.; Ashkar, F.; Bastien, A.; Saadi, H.A.; Gagné, D.; Sirard, M.A.; Khandjian, É.W.; et al. Cumulus Cell Transcripts Transit to the Bovine Oocyte in Preparation for Maturation. Biol. Reprod. 2016, 94, 16. [Google Scholar] [CrossRef]
- Ciesiółka, S.; Budna, J.; Jopek, K.; Bryja, A.; Kranc, W.; Borys, S.; Jeseta, M.; Chachuła, A.; Ziółkowska, A.; Antosik, P.; et al. Time- and Dose-Dependent Effects of 17 Beta-Estradiol on Short-Term, Real-Time Proliferation and Gene Expression in Porcine Granulosa Cells. BioMed Res. Int. 2017, 2017, 9738640. [Google Scholar] [CrossRef] [PubMed]
- Hatzirodos, N.; Glister, C.; Hummitzsch, K.; Irving-Rodgers, H.F.; Knight, P.G.; Rodgers, R.J. Transcriptomal profiling of bovine ovarian granulosa and theca interna cells in primary culture in comparison with their in vivo counterparts. PLoS ONE 2017, 12, e0173391. [Google Scholar] [CrossRef]
- Kempisty, B.; Ziółkowska, A.; Ciesiółka, S.; Piotrowska, H.; Antosik, P.; Bukowska, D.; Nowicki, M.; Brüssow, K.P.; Zabel, M. Study on connexin gene and protein expression and cellular distribution in relation to real-time proliferation of porcine granulosa cells. J. Biol. Regul. Homeost. Agents 2014, 28, 625–635. [Google Scholar] [PubMed]
- Mohammed, B.T.; Donadeu, F.X. Bovine Granulosa Cell Culture. Methods Mol. Biol. 2018, 1817, 79–87. [Google Scholar] [CrossRef]
- Leoni, G.G.; Naitana, S. Ovine Granulosa Cells Isolation and Culture to Improve Oocyte Quality. Methods Mol. Biol. 2018, 1817, 95–106. [Google Scholar] [CrossRef]
- Mansur, A.; Adir, M.; Racowsky, C.; Combelles, C.M.; Landa, N.; Machtinger, R. Susceptibility of human cumulus cells to bisphenol a In vitro. Reprod. Toxicol. 2017, 74, 189–194. [Google Scholar] [CrossRef] [PubMed]
- Saleh, A.C. The Effects of BPA and BPS on anti-Mullerian Hormone and Its Receptor during Early Embryonic Development. Master’s Thesis, Retrieved from The Atrium. University of Guelph, Guelph, ON, Canada, 2020. [Google Scholar]
- Li, T.Y.; Colley, D.; Barr, K.J.; Yee, S.-P.; Kidder, G.M. Rescue of oogenesis in Cx37-null mutant mice by oocyte-specific replacement with Cx43. J. Cell Sci. 2007, 120, 4117–4125. [Google Scholar] [CrossRef]
- Davis, H.M.; Pacheco-Costa, R.; Atkinson, E.G.; Brun, L.R.; Gortazar, A.R.; Harris, J.; Hiasa, M.; Bolarinwa, S.A.; Yoneda, T.; Ivan, M.; et al. Disruption of the Cx43/miR21 pathway leads to osteocyte apoptosis and increased osteoclastogenesis with aging. Aging Cell 2017, 16, 551–563. [Google Scholar] [CrossRef] [PubMed]
- Plotkin, L.I.; Pacheco-Costa, R.; Davis, H.M. MicroRNAs and Connexins in Bone: Interaction and Mechanisms of Delivery. Curr. Mol. Biol. Rep. 2017, 3, 63–70. [Google Scholar] [CrossRef]
- Umrani, M.R.; Joglekar, M.V.; Glover, E.S.; Wong, W.; Hardikar, A.A. Connexins and microRNAs: Interlinked players in regulating islet function? Islets 2017, 9, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Calderón, J.F.; Retamal, M.A. Regulation of Connexins Expression Levels by MicroRNAs, an Update. Front. Physiol. 2016, 7, 558. [Google Scholar] [CrossRef] [PubMed]
- Pan, B.; Toms, D.; Shen, W.; Li, J. MicroRNA-378 regulates oocyte maturation via the suppression of aromatase in porcine cumulus cells. Am. J. Physiol. Metab. 2015, 308, E525–E534. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-K.; Rhee, S.-K. Inhibitory effect of bisphenol A on gap junctional intercellular communication in an epithelial cell line of rat mammary tissue. Arch. Pharmacal Res. 2007, 30, 337–343. [Google Scholar] [CrossRef]
- Oh, S. Bisphenol A and 4-tert-Octylphenol Inhibit Cx46 Hemichannel Currents. Korean J. Physiol. Pharmacol. 2015, 19, 73–79. [Google Scholar] [CrossRef][Green Version]
- Musil, L.S.; Le, A.; Vanslyke, J.K.; Roberts, L.M. Regulation of Connexin Degradation as a Mechanism to Increase Gap Junction Assembly and Function. J. Biol. Chem. 2000, 275, 25207–25215. [Google Scholar] [CrossRef]
- Ge, L.-C.; Chen, Z.-J.; Liu, H.; Zhang, K.-S.; Su, Q.; Ma, X.-Y.; Huang, H.-B.; Zhao, Z.-D.; Wang, Y.-Y.; Giesy, J.P.; et al. Signaling related with biphasic effects of bisphenol A (BPA) on Sertoli cell proliferation: A comparative proteomic analysis. Biochim. Biophys. Acta (BBA) Gen. Subj. 2014, 1840, 2663–2673. [Google Scholar] [CrossRef]
- Grasselli, F.; Baratta, L.; Baioni, L.; Bussolati, S.; Ramoni, R.; Grolli, S.; Basini, G. Bisphenol A disrupts granulosa cell function. Domest. Anim. Endocrinol. 2010, 39, 34–39. [Google Scholar] [CrossRef] [PubMed]
- Campen, K.; Lavallee, M.; Combelles, C. The impact of bisphenol S on bovine granulosa and theca cells. Reprod. Domest. Anim. 2018, 53, 450–457. [Google Scholar] [CrossRef]
- Chen, Y.; Shu, L.; Qiu, Z.; Lee, N.Y.; Settle, S.J.; Hee, S.Q.; Telesca, D.; Yang, X.; Allard, P. Exposure to the BPA-Substitute Bisphenol S Causes Unique Alterations of Germline Function. PLoS Genet. 2016, 12, e1006223. [Google Scholar] [CrossRef] [PubMed]
- Lin, T.; Wang, K.; Chuang, K.; Kao, A.; Kuo, T. Down-regulation of gap junctional intercellular communication and connexin 43 expression by bisphenol A in human granulosa cells. Biotechnol. Appl. Biochem. 2020. [Google Scholar] [CrossRef]
- Zhang, H.; Wei, Q.; Gao, Z.; Ma, C.; Yang, Z.; Zhao, H.; Liu, C.; Liu, J.; Zhao, X.; Ma, B. G protein-coupled receptor 30 mediates meiosis resumption and gap junction communications downregulation in goat cumulus-oocyte complexes by 17β-estradiol. J. Steroid Biochem. Mol. Biol. 2019, 187, 58–67. [Google Scholar] [CrossRef]
- Wang, H.-X.; Gillio-Meina, C.; Chen, S.; Gong, X.-Q.; Li, T.Y.; Bai, D.; Kidder, G.M.; Saccary, L.; She, Y.-M.; Oko, R.; et al. The Canonical WNT2 Pathway and FSH Interact to Regulate Gap Junction Assembly in Mouse Granulosa Cells1. Biol. Reprod. 2013, 89, 39. [Google Scholar] [CrossRef]
- Park, H.-J.; Park, S.-Y.; Kim, J.-W.; Yang, S.-G.; Kim, M.-J.; Jegal, H.-G.; Kim, I.-S.; Choo, Y.-K.; Koo, D.-B. Melatonin Improves Oocyte Maturation and Mitochondrial Functions by Reducing Bisphenol A-Derived Superoxide in Porcine Oocytes In Vitro. Int. J. Mol. Sci. 2018, 19, 3422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Xu, Y.; Kuai, Y.; Wang, S.; Xue, Q.; Shang, J. Effect of testosterone on the Connexin37 of sexual mature mouse cumulus oocyte complex. J. Ovarian Res. 2016, 9, 82. [Google Scholar] [CrossRef][Green Version]
- Zama, A.M.; Uzumcu, M. Epigenetic effects of endocrine-disrupting chemicals on female reproduction: An ovarian perspective. Front. Neuroendocr. 2010, 31, 420–439. [Google Scholar] [CrossRef]
- Wojtanowicz-Markiewicz, K.; Kulus, M.; Knap, S.; Kocherova, I.; Jankowski, M.; Stefańska, K.; Jeseta, M.; Piotrowska-Kempisty, H.; Bukowska, D.; Zabel, M.; et al. Expression of Selected Connexin and Aquaporin Genes and Real-Time Proliferation of Porcine Endometrial Luminal Epithelial Cells in Primary Culture Model. BioMed Res. Int. 2020, 2020, 7120375. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | GenBank Accession # | Source | Primer Sequence (5′-3′) | IVM Primer Efficiency (%) | IVC Primer Efficiency (%) | Product Size (bp) |
|---|---|---|---|---|---|---|
| GAPDH | NM_001034034.1 | [39] | 5′-ttcctggtacgacaatgaatttg-3′ 5′-ggagatggggcaggactc-3′ | 102.3 | 100.8 | 153 |
| PPIA | NM_178320.2 | [10] | 5′-tcttgtccatggcaaatgctg-3′ 5′-tttcacctgccaaagtaccac-3′ | 98.8 | 99.0 | 111 |
| Cx37 | NM_001083738.1 | [28] | 5′-gactcatctccctggtgctc-3′ 5′-gttctgctcactggacgaca-3′ | 97.0 | 100.3 | 221 |
| Cx43 | NM_174068 | [40] | 5′-gtcttcgaggtggccttcttg-3′ 5′-agtccacctgatgtgggcag-3′ | 101.9 | NA | 104 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sabry, R.; Apps, C.; Reiter-Saunders, J.A.; Saleh, A.C.; Balachandran, S.; St. John, E.J.; Favetta, L.A. BPA and BPS Affect Connexin 37 in Bovine Cumulus Cells. Genes 2021, 12, 321. https://doi.org/10.3390/genes12020321
Sabry R, Apps C, Reiter-Saunders JA, Saleh AC, Balachandran S, St. John EJ, Favetta LA. BPA and BPS Affect Connexin 37 in Bovine Cumulus Cells. Genes. 2021; 12(2):321. https://doi.org/10.3390/genes12020321
Chicago/Turabian StyleSabry, Reem, Charlotte Apps, Jaqueline A. Reiter-Saunders, Angela C. Saleh, Sumetha Balachandran, Elizabeth J. St. John, and Laura A. Favetta. 2021. "BPA and BPS Affect Connexin 37 in Bovine Cumulus Cells" Genes 12, no. 2: 321. https://doi.org/10.3390/genes12020321
APA StyleSabry, R., Apps, C., Reiter-Saunders, J. A., Saleh, A. C., Balachandran, S., St. John, E. J., & Favetta, L. A. (2021). BPA and BPS Affect Connexin 37 in Bovine Cumulus Cells. Genes, 12(2), 321. https://doi.org/10.3390/genes12020321