
Functional Haplotype of LIPC Induces Triglyceride-Mediated Suppression of HDL-C Levels According to Genome-Wide Association Studies


Abstract
1. Introduction
2. Materials and Methods
2.1. Participants
2.2. GWAS Population
2.3. Genomic DNA Extraction and Genotyping
2.4. Clinical and Biochemical Analysis
2.5. Haplotype Frequency and Association Analyses
2.6. Mediation Analysis
2.7. Construct
2.8. Culture of HepG2 Cell Lines
2.9. Transfection and Dual Luciferase Assay
2.10. Statistical Analysis
3. Results
3.1. Baseline Characteristics of the TWB Population
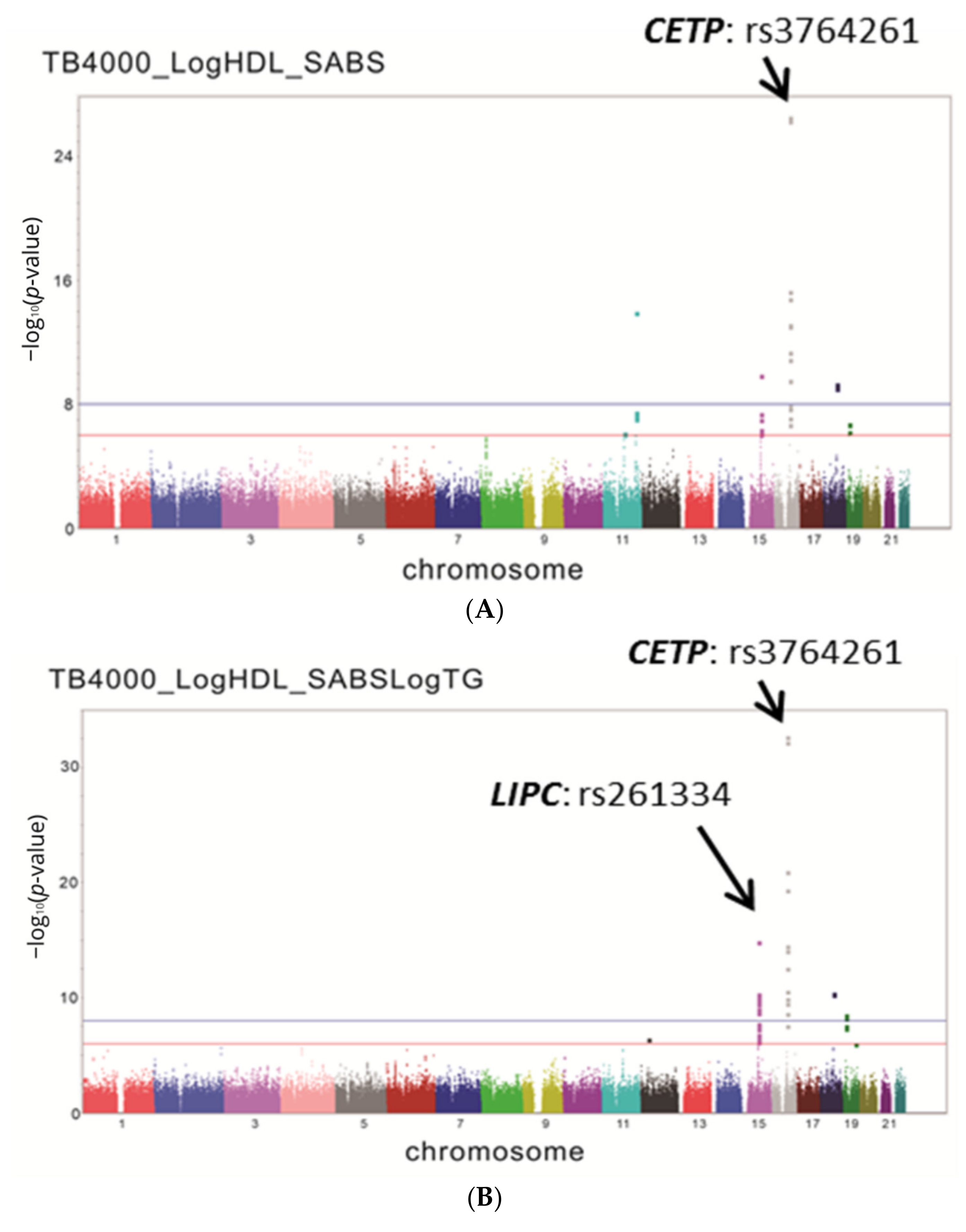
3.2. GWAS for HDL-C in the TWB Population
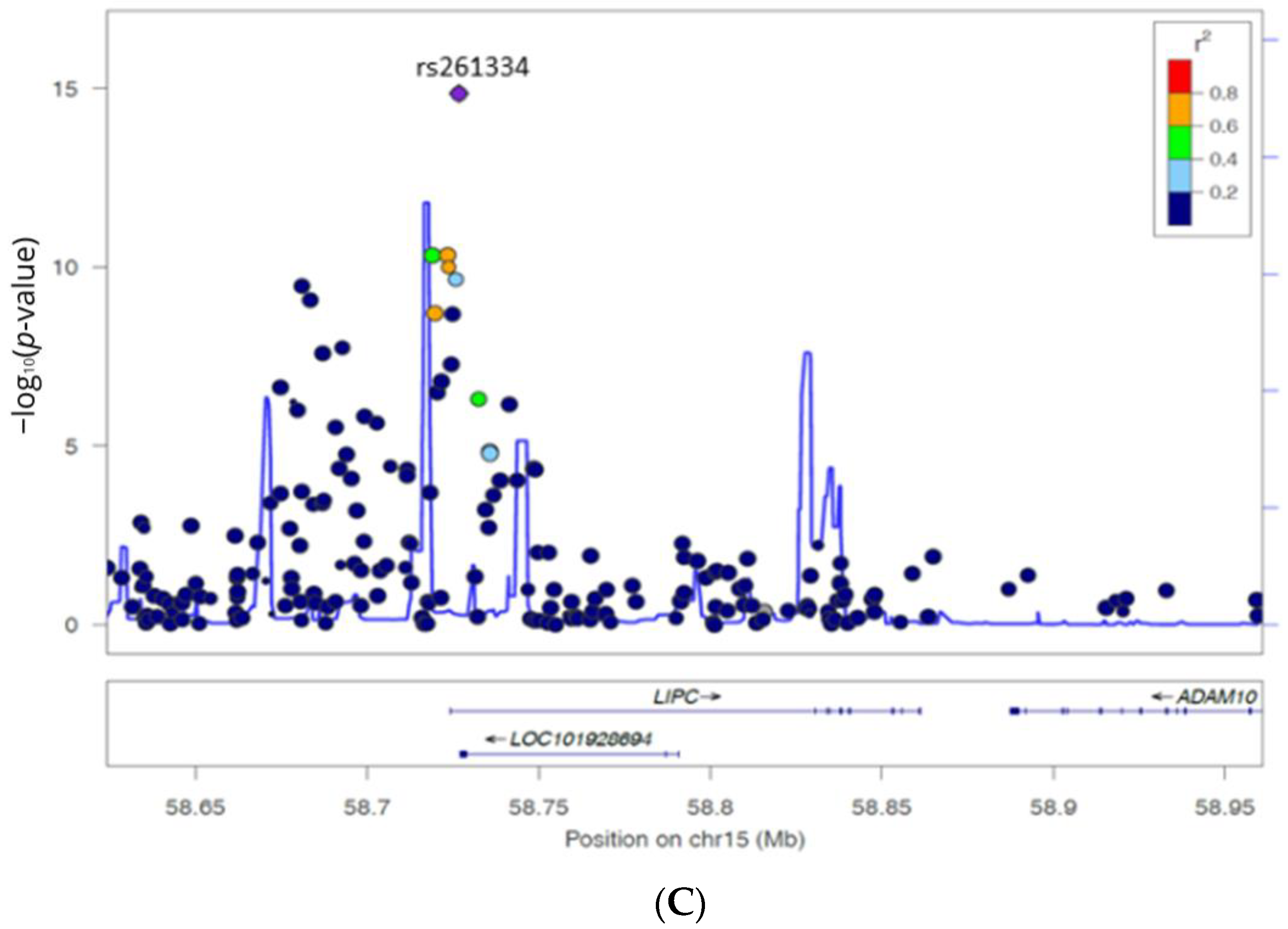
3.3. Association of LIPC Promoter Haplotypes and HDL-C Levels
3.4. Mediation Analysis of the Association between LIPC Haplotype TCG and HDL-C
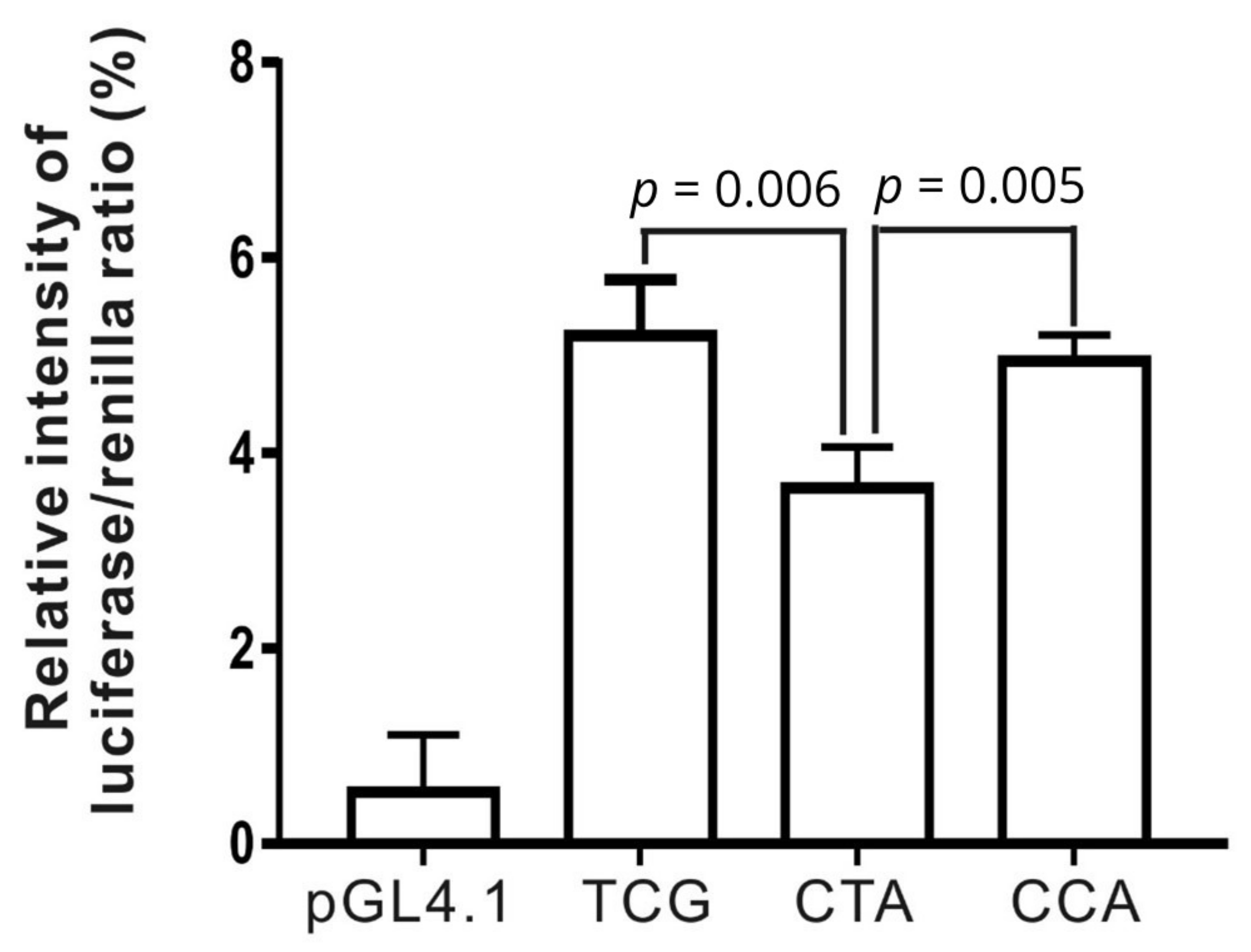
3.5. Functional Analysis of Three Haplotypes in the LIPC Promoter
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Kobayashi, J.; Miyashita, K.; Nakajima, K.; Mabuchi, H. Hepatic Lipase: A Comprehensive View of its Role on Plasma Lipid and Lipoprotein Metabolism. J. Atheroscler. Thrombosis 2015, 22, 1001. [Google Scholar] [CrossRef] [PubMed]
- Annema, W.; Tietge, U.J. Role of hepatic lipase and endothelial lipase in high-density lipoprotein-mediated reverse cholesterol transport. Curr. Atheroscler. Rep. 2011, 13, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Cohen, J.C.; Vega, G.L.; Grundy, S.M. Hepatic lipase: New insights from genetic and metabolic studies. Curr. Opin. Lipidol. 1999, 10, 259–267. [Google Scholar] [CrossRef] [PubMed]
- Thuren, T. Hepatic lipase and HDL metabolism. Curr. Opin. Lipidol. 2000, 11, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Weissglas-Volkov, D.; Pajukanta, P. Genetic causes of high and low serum HDL-cholesterol. J. Lipid Res. 2010, 51, 2032–2057. [Google Scholar] [CrossRef] [PubMed]
- Khetarpal, S.A.; Rader, D.J. Genetics of lipid traits: Genome-wide approaches yield new biology and clues to causality in coronary artery disease. Biochim. Biophys. Acta 2014, 1842, 2010–2020. [Google Scholar] [CrossRef][Green Version]
- Willer, C.J.; Schmidt, E.M.; Sengupta, S.; Peloso, G.M.; Gustafsson, S.; Kanoni, S.; Ganna, A.; Chen, J.; Buchkovich, M.L.; Mora, S.; et al. Discovery and refinement of loci associated with lipid levels. Nat. Genet. 2013, 45, 1274–1283. [Google Scholar] [CrossRef]
- Brunham, L.R.; Hayden, M.R. Human genetics of HDL: Insight into particle metabolism and function. Prog. Lipid Res. 2015, 58, 14–25. [Google Scholar] [CrossRef]
- Dron, J.S.; Hegele, R.A. Genetics of Lipid and Lipoprotein Disorders and Traits. Curr. Genet. Med. Rep. 2016, 4, 130–141. [Google Scholar] [CrossRef]
- Dron, J.S.; Hegele, R.A. Genetics of Triglycerides and the Risk of Atherosclerosis. Curr. Atheroscler. Rep. 2017, 19, 31. [Google Scholar] [CrossRef]
- Aulchenko, Y.S.; Ripatti, S.; Lindqvist, I.; Boomsma, D.; Heid, I.M.; Pramstaller, P.P.; Penninx, B.W.; Janssens, A.C.; Wilson, J.F.; Spector, T.; et al. Loci influencing lipid levels and coronary heart disease risk in 16 European population cohorts. Nat. Genet. 2009, 41, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Chasman, D.I.; Paré, G.; Mora, S.; Hopewell, J.C.; Peloso, G.; Clarke, R.; Cupples, L.A.; Hamsten, A.; Kathiresan, S.; Mälarstig, A.; et al. Forty-three loci associated with plasma lipoprotein size, concentration and cholesterol content in genome-wide analysis. PLoS Genet. 2009, 5, e1000730. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Huang, J.; Mo, Z.; He, J.; Wang, L.; Yang, X.; Tan, A.; Chen, S.; Chen, J.; Gu, C.C.; et al. Genetic Susceptibility to Lipid Levels and Lipid Change Over Time and Risk of Incident Hyperlipidemia in Chinese Populations. Circ. Cardiovasc. Genet. 2016, 9, 37–44. [Google Scholar] [CrossRef]
- Ko, Y.L.; Hsu, L.A.; Hsu, K.H.; Ko, Y.H.; Lee, Y.S. The interactive effects of hepatic lipase gene promoter polymorphisms with sex and obesity on high-density-lipoprotein cholesterol levels in Taiwanese-Chinese. Atherosclerosis 2004, 172, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, K.; Bujo, H.; Taira, K.; Itou, N.; Shibasaki, M.; Takahashi, K.; Saito, Y. Increased circulating malondialdehyde-modified LDL in the patients with familial combined hyperlipidemia and its relation with the hepatic lipase activity. Atherosclerosis 2004, 172, 181–187. [Google Scholar] [CrossRef]
- Isaacs, A.; Sayed-Tabatabaei, F.A.; Njajou, O.T.; Witteman, J.C.; van Duijn, C.M. The -514 C->T hepatic lipase promoter region polymorphism and plasma lipids: A meta-analysis. J. Clin. Endocrinol. Metab. 2004, 89, 3858–3863. [Google Scholar] [CrossRef]
- Ma, L.; Brautbar, A.; Boerwinkle, E.; Sing, C.F.; Clark, A.G.; Keinan, A. Knowledge-driven analysis identifies a gene-gene interaction affecting high-density lipoprotein cholesterol levels in multi-ethnic populations. PLoS Genet. 2012, 8, e1002714. [Google Scholar] [CrossRef]
- Hall, M.A.; Verma, A.; Brown-Gentry, K.D.; Goodloe, R.; Boston, J.; Wilson, S.; McClellan, B.; Sutcliffe, C.; Dilks, H.H.; Gillani, N.B.; et al. Detection of pleiotropy through a Phenome-wide association study (PheWAS) of epidemiologic data as part of the Environmental Architecture for Genes Linked to Environment (EAGLE) study. PLoS Genet. 2014, 10, e1004678. [Google Scholar] [CrossRef]
- James, L.R.; Brett, J.M. Mediators, moderators and tests for mediation. J. Appl. Psychol. 1984, 69, 307. [Google Scholar] [CrossRef]
- Alwin, D.F.; Hauser, R.M. The decomposition of effects in path analysis. Am. Sociol. Rev. 1975, 40, 37–47. [Google Scholar] [CrossRef]
- Baron, R.M.; Kenny, D.A. The moderator-mediator variable distinction in social psychological research: Conceptual, strategic and statistical considerations. J. Personal. Soc. Psychol. 1986, 51, 1173–1182. [Google Scholar] [CrossRef]
- Sobel, M.E. Effect analysis and causation in linear structural equation models. Psychometrika 1990, 55, 495–515. [Google Scholar] [CrossRef]
- Tzelgov, J.; Henik, A. Suppression situations in psychological research: Definitions, implications and applications. Psychol. Bull. 1991, 109, 524. [Google Scholar] [CrossRef]
- Teng, M.S.; Hsu, L.A.; Wu, S.; Chou, H.H.; Chang, C.J.; Sun, Y.Z.; Juan, S.H.; Ko, Y.L. Mediation analysis reveals a sex-dependent association between ABO gene variants and TG/HDL-C ratio that is suppressed by sE-selectin level. Atherosclerosis 2013, 228, 406–412. [Google Scholar] [CrossRef]
- Teng, M.S.; Hsu, L.A.; Wu, S.; Sun, Y.C.; Juan, S.H.; Ko, Y.L. Association of CDH13 genotypes/haplotypes with circulating adiponectin levels, metabolic syndrome and related metabolic phenotypes: The role of the suppression effect. PLoS ONE 2015, 10, e0122664. [Google Scholar] [CrossRef]
- Teng, M.S.; Wu, S.; Hsu, L.A.; Tzeng, I.S.; Chou, H.H.; Su, C.W.; Ko, Y.L. Pleiotropic association of LIPC variants with lipid and urinary 8-hydroxy deoxyguanosine levels in a Taiwanese population. Lipids Health Dis. 2019, 18, 111. [Google Scholar] [CrossRef]
- Chen, C.H.; Yang, J.H.; Chiang, C.W.K.; Hsiung, C.N.; Wu, P.E.; Chang, L.C.; Chu, H.W.; Chang, J.; Song, I.W.; Yang, S.L.; et al. Population structure of Han Chinese in the modern Taiwanese population based on 10,000 participants in the Taiwan Biobank project. Hum. Mol. Genet. 2016, 25, 5321–5331. [Google Scholar] [CrossRef]
- MacKinnon, D.P. Analysis of mediating variables in prevention and intervention research. NIDA Res. Monogr. 1994, 139, 127–153. [Google Scholar]
- Sobel, M.E. Asymptotic confidence intervals for indirect effects in structural equation models. Sociol. Methodol. 1982, 13, 290–312. [Google Scholar] [CrossRef]
- Sobel, M.E. Direct and indirect effects in linear structural equation models. Sociol. Methods Res. 1987, 16, 155–176. [Google Scholar] [CrossRef]
- MacKinnon, D.P.; Krull, J.L.; Lockwood, C.M. Equivalence of the mediation, confounding and suppression effect. Prev. Sci. Off. J. Soc. Prev. Res. 2000, 1, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Boekholdt, S.M.; Thompson, J.F. Natural genetic variation as a tool in understanding the role of CETP in lipid levels and disease. J. Lipid Res. 2003, 44, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Tall, A.R. Plasma cholesteryl ester transfer protein. J. Lipid Res. 1993, 34, 1255–1274. [Google Scholar] [CrossRef]
- Barter, P. CETP and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 2029–2031. [Google Scholar] [CrossRef]
- Shrestha, S.; Wu, B.J.; Guiney, L.; Barter, P.J.; Rye, K.A. Cholesteryl ester transfer protein and its inhibitors. J. Lipid Res. 2018, 59, 772–783. [Google Scholar] [CrossRef]
- Feitosa, M.F.; Myers, R.H.; Pankow, J.S.; Province, M.A.; Borecki, I.B. LIPC variants in the promoter and intron 1 modify HDL-C levels in a sex-specific fashion. Atherosclerosis 2009, 204, 171–177. [Google Scholar] [CrossRef]
- Piko, P.; Fiatal, S.; Werissa, N.A.; Bekele, B.B.; Racz, G.; Kosa, Z.; Sandor, J.; Adany, R. The Effect of Haplotypes in the CETP and LIPC Genes on the Triglycerides to HDL-C Ratio and Its Components in the Roma and Hungarian General Populations. Genes 2020, 11, 56. [Google Scholar] [CrossRef]
- Shohet, R.V.; Vega, G.L.; Anwar, A.; Cigarroa, J.E.; Grundy, S.M.; Cohen, J.C. Hepatic lipase (LIPC) promoter polymorphism in men with coronary artery disease. Allele frequency and effects on hepatic lipase activity and plasma HDL-C concentrations. Arterioscler. Thromb. Vasc. Biol. 1999, 19, 1975–1978. [Google Scholar] [CrossRef]
- Lahoz, C.; Peña, R.; Mostaza, J.M.; Laguna, F.; García-Iglesias, M.F.; Taboada, M.; Pintó, X. The -514C/T polymorphism of the hepatic lipase gene significantly modulates the HDL-cholesterol response to statin treatment. Atherosclerosis 2005, 182, 129–134. [Google Scholar] [CrossRef]
- Guerra, R.; Wang, J.; Grundy, S.M.; Cohen, J.C. A hepatic lipase (LIPC) allele associated with high plasma concentrations of high density lipoprotein cholesterol. Proc. Natl. Acad. Sci. USA 1997, 94, 4532–4537. [Google Scholar] [CrossRef]
- Couture, P.; Otvos, J.D.; Cupples, L.A.; Lahoz, C.; Wilson, P.W.; Schaefer, E.J.; Ordovas, J.M. Association of the C-514T polymorphism in the hepatic lipase gene with variations in lipoprotein subclass profiles: The Framingham Offspring Study. Arterioscler. Thromb. Vasc. Biol. 2000, 20, 815–822. [Google Scholar] [CrossRef] [PubMed]
- Hadzopoulou-Cladaras, M.; Cardot, P. Identification of a cis-acting negative DNA element which modulates human hepatic triglyceride lipase gene expression. Biochemistry 1993, 32, 9657–9667. [Google Scholar] [CrossRef] [PubMed]
- Quandt, K.; Frech, K.; Karas, H.; Wingender, E.; Werner, T. MatInd and MatInspector: New fast and versatile tools for detection of consensus matches in nucleotide sequence data. Nucleic Acids Res. 1995, 23, 4878–4884. [Google Scholar] [CrossRef] [PubMed]
- Oka, K.; Ishimura-Oka, K.; Chu, M.J.; Chan, L. Transcription of the human hepatic lipase gene is modulated by multiple negative elements in HepG2 cells. Gene 1996, 180, 69–80. [Google Scholar] [CrossRef]
- Wang, D.; Sul, H.S. Insulin stimulation of the fatty acid synthase promoter is mediated by the phosphatidylinositol 3-kinase pathway. Involvement of protein kinase B/Akt. J. Biol. Chem. 1998, 273, 25420–25426. [Google Scholar] [CrossRef]
- Botma, G.J.; Verhoeven, A.J.; Jansen, H. Hepatic lipase promoter activity is reduced by the C-480T and G-216A substitutions present in the common LIPC gene variant and is increased by Upstream Stimulatory Factor. Atherosclerosis 2001, 154, 625–632. [Google Scholar] [CrossRef]
- Olivecrona, G.; Olivecrona, T. Triglyceride lipases and atherosclerosis. Curr. Opin. Lipidol. 1995, 6, 291–305. [Google Scholar] [CrossRef]
- Santamarina-Fojo, S.; Haudenschild, C.; Amar, M. The role of hepatic lipase in lipoprotein metabolism and atherosclerosis. Curr. Opin. Lipidol. 1998, 9, 211–219. [Google Scholar] [CrossRef]
- Tahvanainen, E.; Syvanne, M.; Frick, M.H.; Murtomaki-Repo, S.; Antikainen, M.; Kesaniemi, Y.A.; Kauma, H.; Pasternak, A.; Taskinen, M.R.; Ehnholm, C. Association of variation in hepatic lipase activity with promoter variation in the hepatic lipase gene. The LOCAT Study Invsestigators. J. Clin. Investig. 1998, 101, 956–960. [Google Scholar] [CrossRef]
- Silbernagel, G.; Scharnagl, H.; Kleber, M.E.; Delgado, G.; Stojakovic, T.; Laaksonen, R.; Erdmann, J.; Rankinen, T.; Bouchard, C.; Landmesser, U.; et al. LDL triglycerides, hepatic lipase activity and coronary artery disease: An epidemiologic and Mendelian randomization study. Atherosclerosis 2019, 282, 37–44. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Total | Men | Women | p-Value | |
|---|---|---|---|---|
| Number | 4000 | 1734 | 2266 | |
| Age (years) | 48 (39–57) | 47 (38–57) | 48 (40–56) | 0.558 |
| BMI (kg/m2) | 24.1 ± 3.5 | 25.1 ± 3.4 | 23.4 ± 3.4 | <0.001 |
| Waist circumference (cm) | 83 (77–89.6) | 87 (82-92.9) | 79.5 (74–86) | <0.001 |
| Waist-to-hip ratio | 0.87 (0.82-0.91) | 0.89 (0.86-0.93) | 0.84 (0.8-0.89) | <0.001 |
| Current smokers (%) | 693 (17.3%) | 570 (32.9%) | 123 (5.4%) | <0.001 |
| Total cholesterol (mg/dL) | 194.4 ± 35.5 | 192.8 ± 34.4 | 195.6 ± 36.3 | 0.015 |
| LDL-C (mg/dL) | 121.2 ± 31.9 | 122.7 ± 31.6 | 120.0 ± 32.1 | 0.006 |
| HDL-C (mg/dL) | 53 (45–63) | 48 (41–55) | 58 (49–67) | <0.001 |
| TG (mg/dL) | 94 (66–136) | 111 (77–164) | 82.5 (60–118) | <0.001 |
| SNP1 | SNP2 | SNP3 | Frequency | TG | HDL-C | p1 Value | |||
|---|---|---|---|---|---|---|---|---|---|
| Coefficient | p | Coefficient | p | ||||||
| H1 | T | C | G | 0.613 | −0.05 | 0.025 | −0.021 | 0.05 | 0.0003 |
| H2 | C | T | A | 0.361 | 0.01 | 0.161 | 0.023 | 0.03 | 0.0009 |
| H3 | C | C | A | 0.025 | 0.16 | 0.016 | −0.01 | 0.8 | 0.28 |
| H1-TCG | ||
|---|---|---|
| Criterion 1 | α | |
| regression coefficient | −0.053 | |
| Standard error | 0.023 | |
| p value | 0.025 | |
| Criterion 2 | β | |
| regression coefficient | −0.22 | |
| Standard error | 0.014 | |
| #p value | 9.17 × 10−48 | |
| γ′ | ||
| regression coefficient | −0.033 | |
| Standard error | 0.009 | |
| * p value | 0.0003 | |
| Criterion 3 | αβ + γ′ | |
| regression coefficient | −0.021 | |
| Standard error | 0.01 | |
| p value | 0.05 | |
| Criterion 4 | αβ | |
| regression coefficient | 0.012 | |
| Standard error | 0.005 | |
| p value (Sobel test) | 0.023 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liao, Y.-H.; Er, L.-K.; Wu, S.; Ko, Y.-L.; Teng, M.-S. Functional Haplotype of LIPC Induces Triglyceride-Mediated Suppression of HDL-C Levels According to Genome-Wide Association Studies. Genes 2021, 12, 148. https://doi.org/10.3390/genes12020148
Liao Y-H, Er L-K, Wu S, Ko Y-L, Teng M-S. Functional Haplotype of LIPC Induces Triglyceride-Mediated Suppression of HDL-C Levels According to Genome-Wide Association Studies. Genes. 2021; 12(2):148. https://doi.org/10.3390/genes12020148
Chicago/Turabian StyleLiao, Yu-Huang, Leay-Kiaw Er, Semon Wu, Yu-Lin Ko, and Ming-Sheng Teng. 2021. "Functional Haplotype of LIPC Induces Triglyceride-Mediated Suppression of HDL-C Levels According to Genome-Wide Association Studies" Genes 12, no. 2: 148. https://doi.org/10.3390/genes12020148
APA StyleLiao, Y.-H., Er, L.-K., Wu, S., Ko, Y.-L., & Teng, M.-S. (2021). Functional Haplotype of LIPC Induces Triglyceride-Mediated Suppression of HDL-C Levels According to Genome-Wide Association Studies. Genes, 12(2), 148. https://doi.org/10.3390/genes12020148