
Gene Expression Profiling of Skeletal Muscles

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice
2.2. RNA-Sequencing (RNA-Seq) Data Analysis
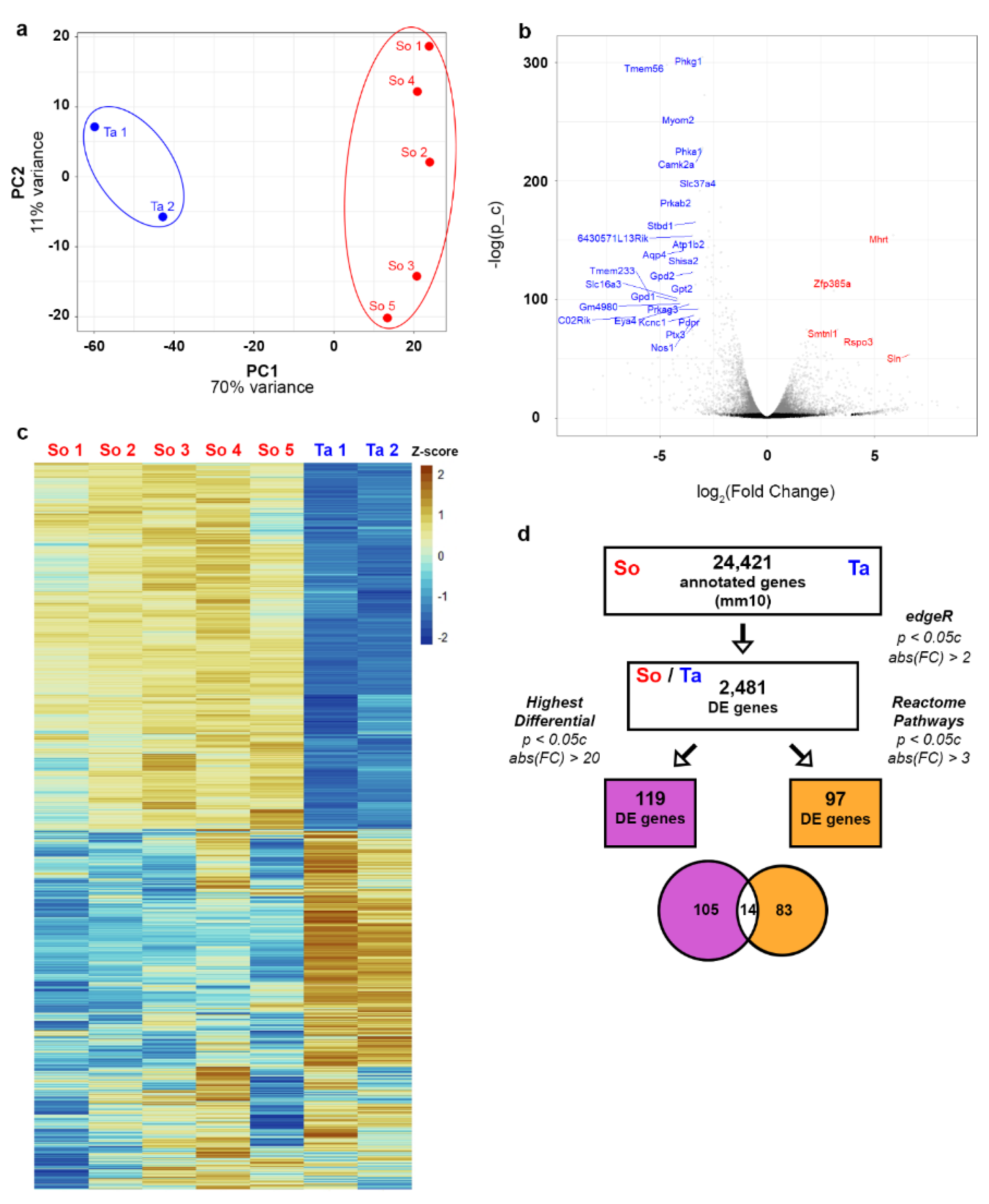
3. Results
3.1. RNA-Seq Data Quality Check
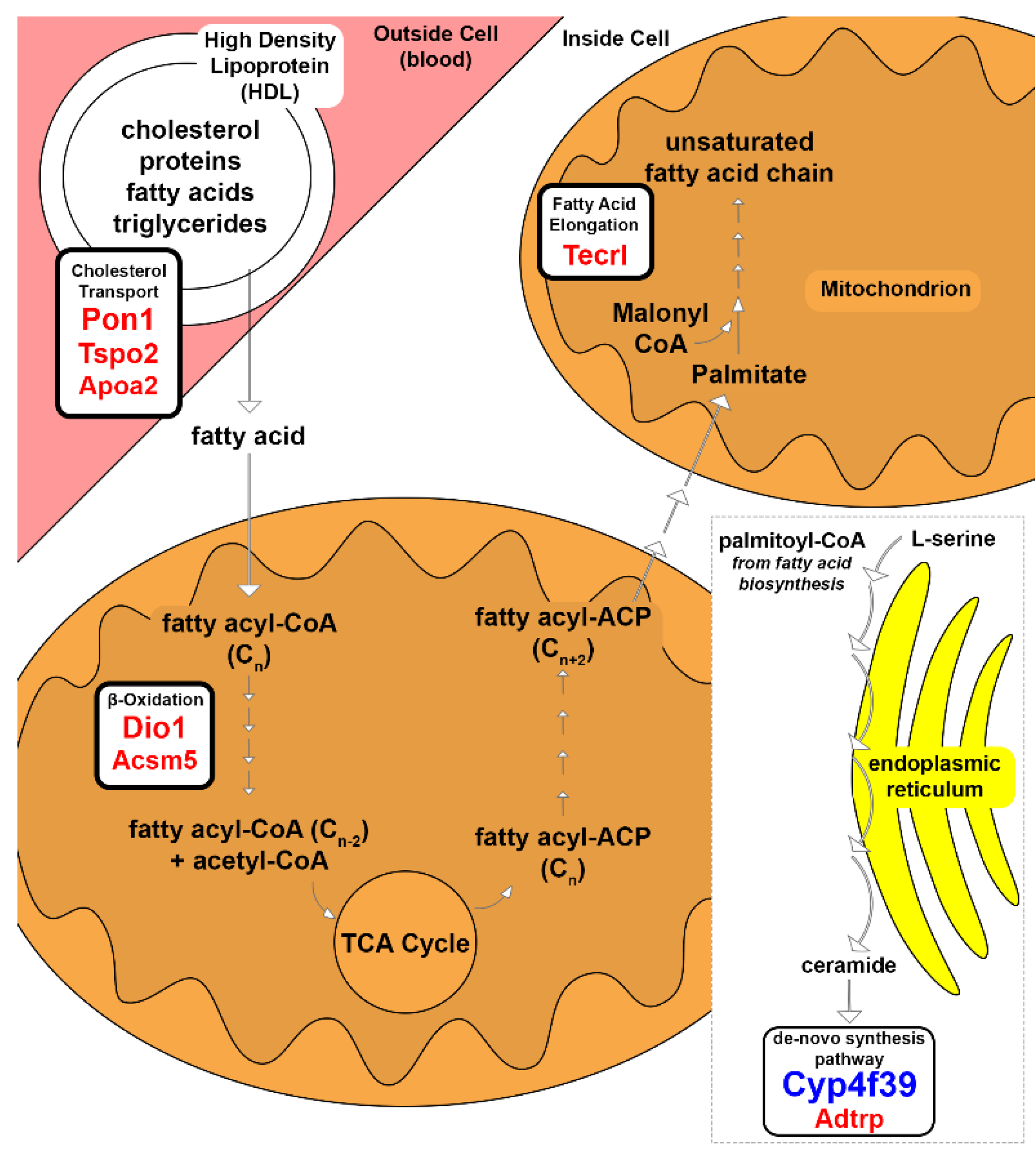
3.2. Lipid Metabolism
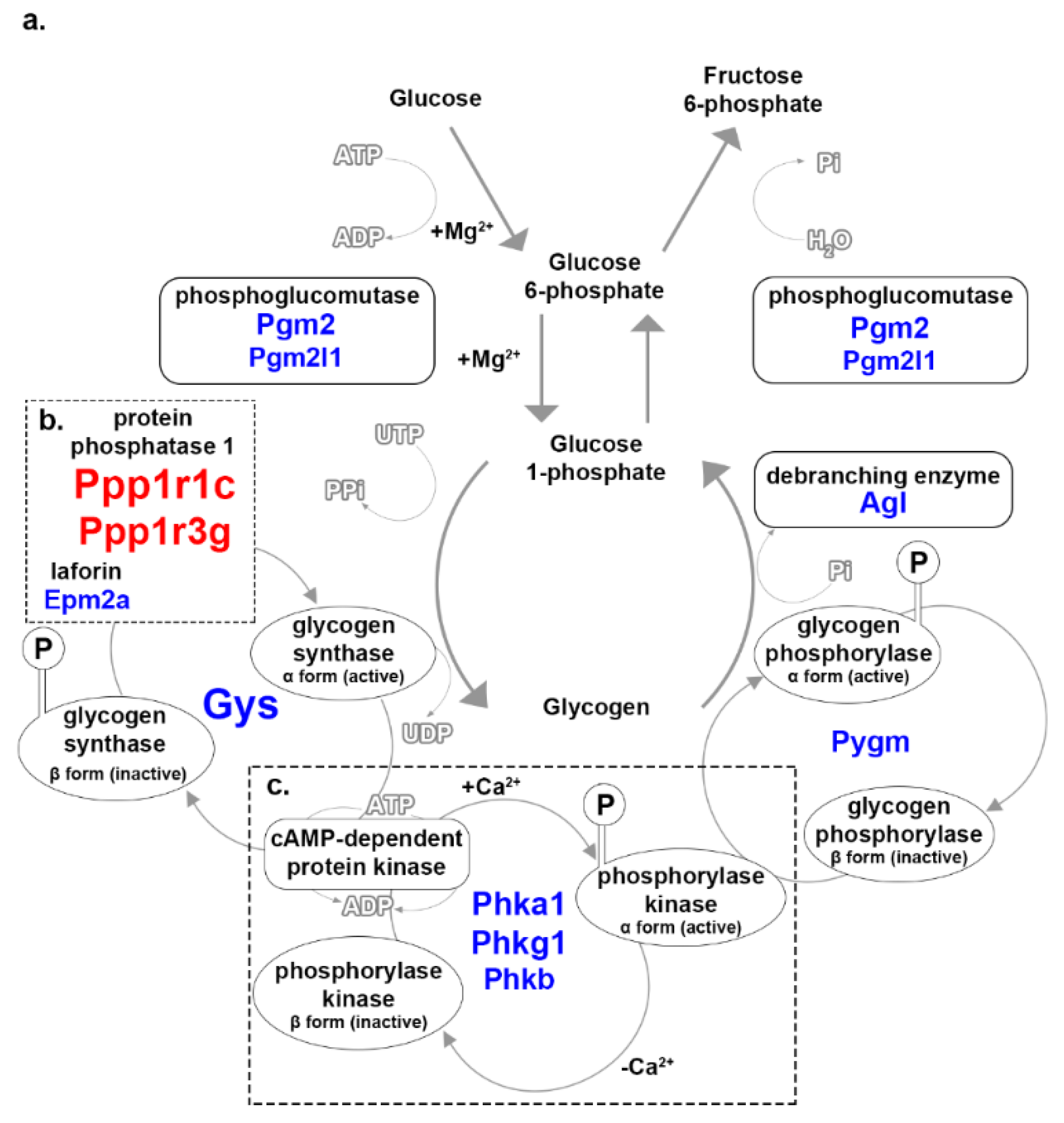
3.3. Glycogen Metabolism
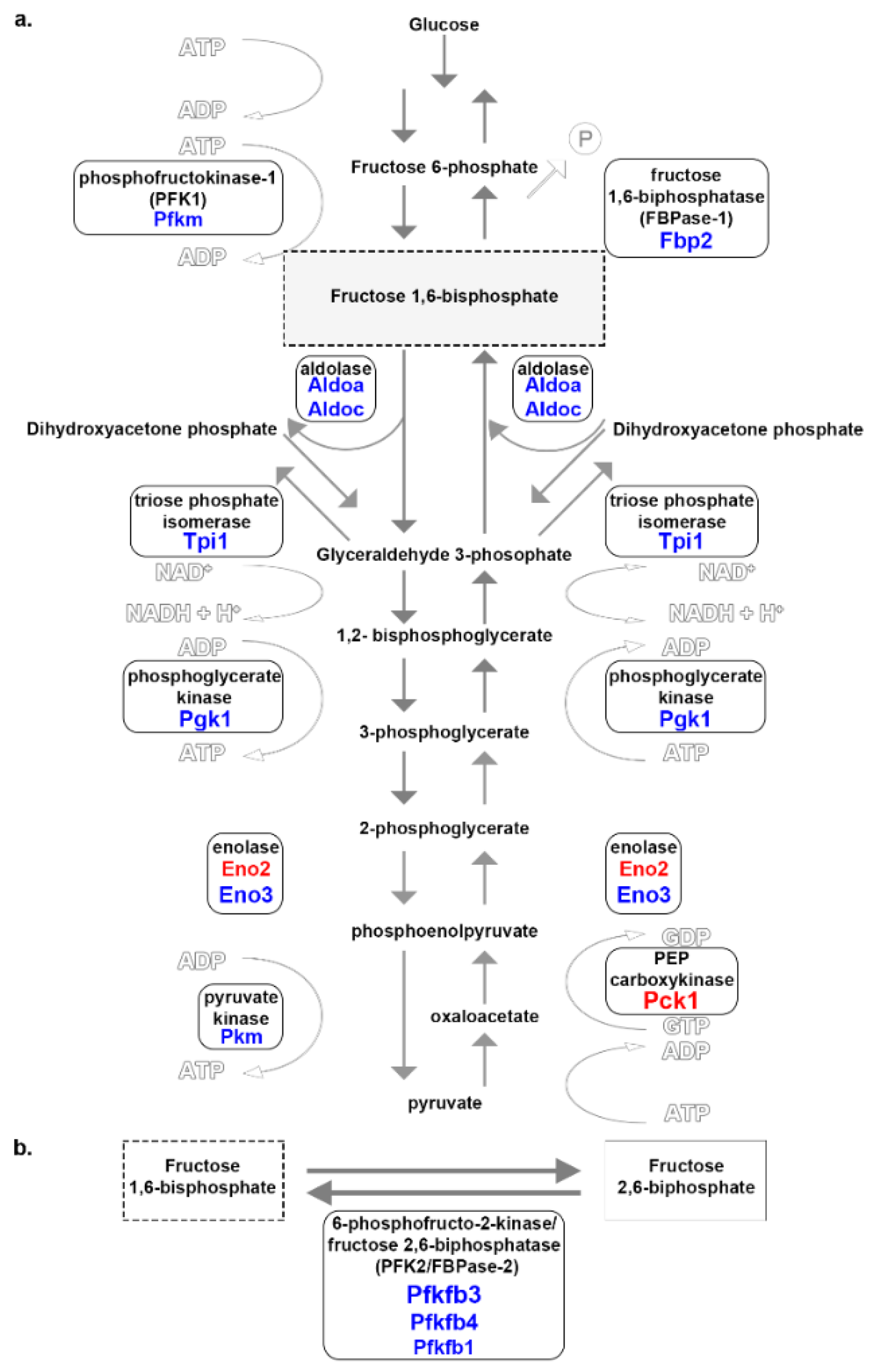
3.4. Glucose Metabolism
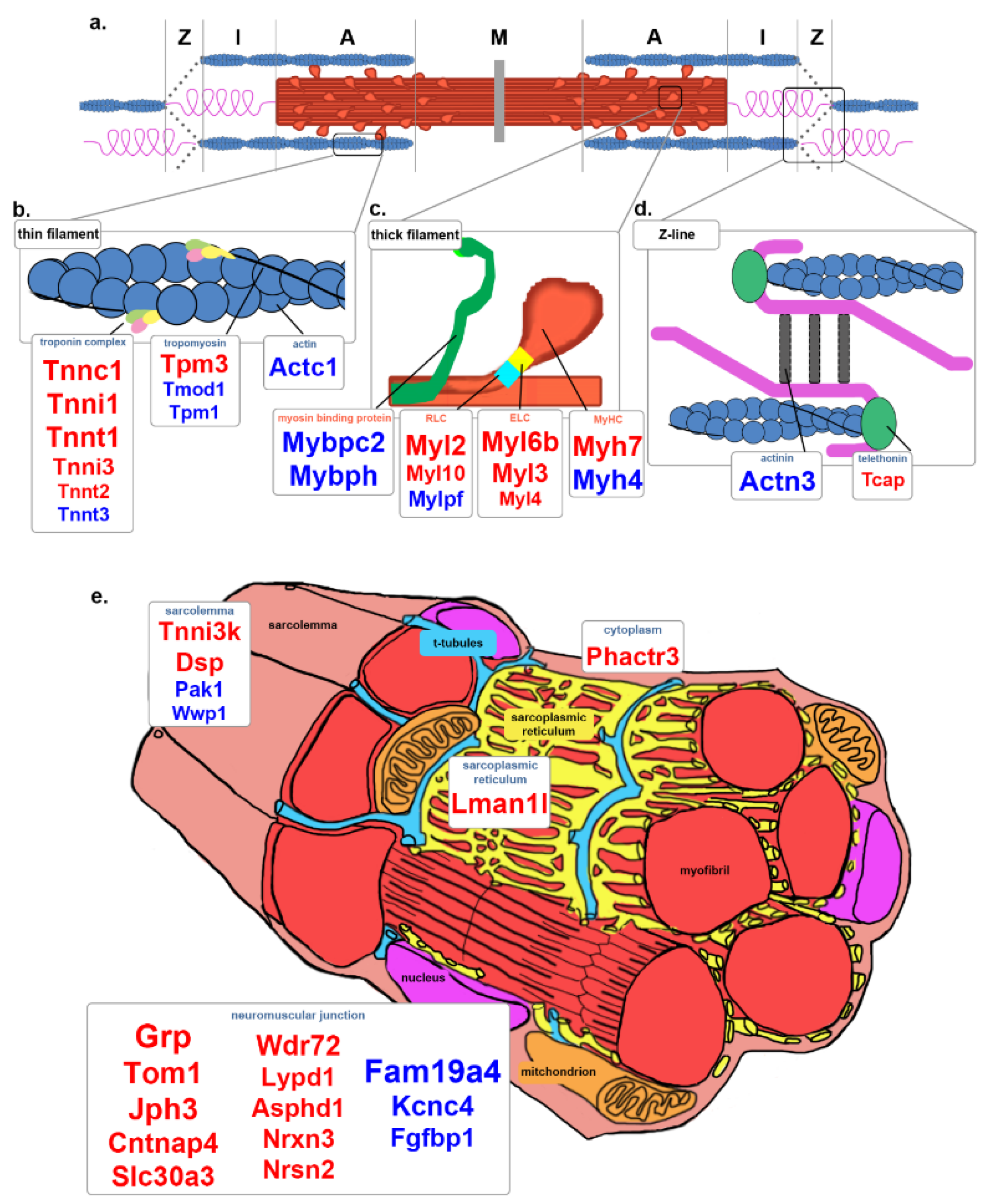
3.5. Contraction
4. Discussion
4.1. Lipid Metabolism
4.2. Glycogen and Glycosaminoglycan Metabolism
4.3. Glucose Metabolism
4.4. Contraction
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schiaffino, S.; Rossi, A.; Smerdu, V.; Leinwand, L.; Reggiani, C. Developmental Myosins: Expression Patterns and Functional Significance. Skelet. Muscle 2015, 5, 22. [Google Scholar] [CrossRef] [PubMed]
- Schiaffino, S.; Reggiani, C. Fiber Types in Mammalian Skeletal Muscles. Physiol. Rev. 2011, 91, 1447–1531. [Google Scholar] [CrossRef]
- Fujita, N.; Fujimoto, T.; Tasaki, H.; Arakawa, T.; Matsubara, T.; Miki, A. Influence of Muscle Length on Muscle Atrophy in the Mouse Tibialis Anterior and Soleus Muscles. Biomed. Res. 2009, 30, 39–45. [Google Scholar] [CrossRef]
- Kammoun, M.; Cassar-Malek, I.; Meunier, B.; Picard, B. A Simplified Immunohistochemical Classification of Skeletal Muscle Fibres in Mouse. Eur. J. Histochem. 2014, 58, 2254. [Google Scholar] [CrossRef]
- Singh, A.J.; Chang, C.-N.; Ma, H.-Y.; Ramsey, S.A.; Filtz, T.M.; Kioussi, C. FACS-Seq Analysis of Pax3-Derived Cells Identifies Non-Myogenic Lineages in the Embryonic Forelimb. Sci. Rep. 2018, 8, 7670. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Fang, Z.; Zhang, K.; Zhi, D.; Cui, X. Length Bias Correction for RNA-Seq Data in Gene Set Analyses. Bioinformatics 2011, 27, 662–669. [Google Scholar] [CrossRef]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. EdgeR: A Bioconductor Package for Differential Expression Analysis of Digital Gene Expression Data. Bioinformatics 2009, 26, 139–140. [Google Scholar] [CrossRef] [PubMed]
- Croft, D.; O’Kelly, G.; Wu, G.; Haw, R.; Gillespie, M.; Matthews, L.; Caudy, M.; Garapati, P.; Gopinath, G.; Jassal, B.; et al. Reactome: A Database of Reactions, Pathways and Biological Processes. Nucleic Acids Res. 2011, 39, D691–D697. [Google Scholar] [CrossRef]
- Yu, G.; He, Q.-Y. ReactomePA: An R/Bioconductor Package for Reactome Pathway Analysis and Visualization. Mol. BioSyst. 2016, 12, 477–479. [Google Scholar] [CrossRef]
- Shih, D.M.; Gu, L.; Xia, Y.R.; Navab, M.; Li, W.F.; Hama, S.; Castellani, L.W.; Furlong, C.E.; Costa, L.G.; Fogelman, A.M.; et al. Mice Lacking Serum Paraoxonase Are Susceptible to Organophosphate Toxicity and Atherosclerosis. Nature 1998, 394, 284–287. [Google Scholar] [CrossRef]
- Fan, J.; Rone, M.; Papadopoulos, V. Translocator Protein 2 Is Involved in Cholesterol Redistribution during Erythropoiesis. J. Biol. Chem. 2009, 284, 30484–30497. [Google Scholar] [CrossRef]
- Yang, M.; Liu, Y.; Dai, J.; Li, L.; Ding, X.; Xu, Z.; Mori, M.; Miyahara, H.; Sawashita, J.; Higuchi, K. Apolipoprotein A-II Induces Acute-Phase Response Associated AA Amyloidosis in Mice through Conformational Changes of Plasma Lipoprotein Structure. Sci. Rep. 2018, 8, 5620. [Google Scholar] [CrossRef]
- Puig-Oliveras, A.; Revilla, M.; Castelló, A.; Fernández, A.I.; Folch, J.M.; Ballester, M. Expression-Based GWAS Identifies Variants, Gene Interactions and Key Regulators Affecting Intramuscular Fatty Acid Content and Composition in Porcine Meat. Sci. Rep. 2016, 6, 31803. [Google Scholar] [CrossRef]
- Bianco, A.C.; Salvatore, D.; Gereben, B.; Berry, M.J.; Larsen, P.R. Biochemistry, Cellular and Molecular Biology, and Physiological Roles of the Iodothyronine Selenodeiodinases. Endocr. Rev. 2002, 23, 38–89. [Google Scholar] [CrossRef]
- Panicker, V.; Cluett, C.; Shields, B.; Murray, A.; Parnell, K.; Perry, J.; Weedon, M.; Singleton, A.; Hernandez, D.; Evans, J.; et al. A Common Variation in Deiodinase 1 Gene DIO1 Is Associated with the Relative Levels of Free Thyroxine and Triiodothyronine. J. Clin. Endocrinol. Metab. 2008, 93, 3075–3081. [Google Scholar] [CrossRef]
- Salvatore, D.; Simonides, W.S.; Dentice, M.; Zavacki, A.M.; Larsen, P.R. Thyroid Hormones and Skeletal Muscle-New Insights and Potential Implications. Nat. Rev. Endocrinol. 2014, 10, 206–214. [Google Scholar] [CrossRef]
- Devalla, H.D.; Gélinas, R.; Aburawi, E.H.; Beqqali, A.; Goyette, P.; Freund, C.; Chaix, M.; Tadros, R.; Jiang, H.; le Béchec, A.; et al. TECRL, a New Life-threatening Inherited Arrhythmia Gene Associated with Overlapping Clinical Features of Both LQTS and CPVT. EMBO Mol. Med. 2016, 8, 1390–1408. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.U.; Sass, S.; Mueller, N.S.; Krebs, S.; Bauersachs, S.; Kaiser, S.; Blum, H.; Thirion, C.; Krause, S.; Theis, F.J.; et al. Integrative Analysis of MicroRNA and MRNA Data Reveals an Orchestrated Function of MicroRNAs in Skeletal Myocyte Differentiation in Response to TNF-α or IGF1. PLoS ONE 2015, 10, e0135284. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, M.; Itoh, N.; Sawai, M.; Sassa, T.; Kihara, A. Severe Skin Permeability Barrier Dysfunction in Knockout Mice Deficient in a Fatty Acid ω-Hydroxylase Crucial to Acylceramide Production. J. Investig. Dermatol. 2020, 140, 319–326. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska, E.; Blachnio-Zabielska, A. The Role of Ceramides in Insulin Resistance. Front. Endocrinol. 2019, 10, 577. [Google Scholar] [CrossRef]
- Piekorz, R.P.; Gingras, S.; Hoffmeyer, A.; Ihle, J.N.; Weinstein, Y. Regulation of Progesterone Levels during Pregnancy and Parturition by Signal Transducer and Activator of Transcription 5 and 20α-Hydroxysteroid Dehydrogenase. Mol. Endocrinol. 2005, 19, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Parsons, W.H.; Kolar, M.J.; Kamat, S.S.; Cognetta, A.B., III; Hulce, J.J.; Saez, E.; Kahn, B.B.; Saghatelian, A.; Cravatt, B.F. AIG1 and ADTRP Are Atypical Integral Membrane Hydrolases That Degrade Bioactive FAHFAs. Nat. Chem. Biol. 2016, 12, 367–372. [Google Scholar] [CrossRef] [PubMed]
- Terao, M.; Romão, M.J.; Leimkühler, S.; Bolis, M.; Fratelli, M.; Coelho, C.; Santos-Silva, T.; Garattini, E. Structure and Function of Mammalian Aldehyde Oxidases. Arch. Toxicol. 2016, 90, 753–780. [Google Scholar] [CrossRef] [PubMed]
- Alnouti, Y.; Klaassen, C.D. Tissue Distribution, Ontogeny, and Regulation of Aldehyde Dehydrogenase (Aldh) Enzymes MRNA by Prototypical Microsomal Enzyme Inducers in Mice. Toxicol. Sci. 2008, 101, 51–64. [Google Scholar] [CrossRef]
- Zhang, Y.; Gu, J.; Wang, L.; Zhao, Z.; Pan, Y.; Chen, Y. Ablation of PPP1R3G Reduces Glycogen Deposition and Mitigates High-Fat Diet Induced Obesity. Mol. Cell. Endocrinol. 2017, 439, 133–140. [Google Scholar] [CrossRef]
- Irimia, J.M.; Meyer, C.M.; Peper, C.L.; Zhai, L.; Bock, C.B.; Previs, S.F.; McGuinness, O.P.; DePaoli-Roach, A.; Roach, P.J. Impaired Glucose Tolerance and Predisposition to the Fasted State in Liver Glycogen Synthase Knock-out Mice. J. Biol. Chem. 2010, 285, 12851–12861. [Google Scholar] [CrossRef]
- Ganesh, S.; Lal Agarwala, K.; Ueda, K.; Akagi, T.; Shoda, K.; Usui, T.; Hashikawa, T.; Osada, H.; Delgado-Escueta, A.V.; Yamakawa, K. Defective in the Progressive Myoclonus Epilepsy of Lafora Type, Is a Dual-Specificity Phosphatase Associated with Polyribosomes. Hum. Mol. Genet. 2000, 9, 2251–2261. [Google Scholar] [CrossRef] [PubMed]
- Garyali, P.; Siwach, P.; Singh, P.K.; Puri, R.; Mittal, S.; Sengupta, S.; Parihar, R.; Ganesh, S. The Malin-Laforin Complex Suppresses the Cellular Toxicity of Misfolded Proteins by Promoting Their Degradation through the Ubiquitin-Proteasome System. Hum. Mol. Genet. 2009, 18, 688–700. [Google Scholar] [CrossRef]
- Roach, P.J. Glycogen and Its Metabolism. Curr. Mol. Med. 2002, 2, 101–120. [Google Scholar] [CrossRef]
- Chesney, J.; Telang, S.; Yalcin, A.; Clem, A.; Wallis, N.; Bucala, R. Targeted Disruption of Inducible 6-Phosphofructo-2-Kinase Results in Embryonic Lethality. Biochem. Biophys. Res. Commun. 2005, 331, 139–146. [Google Scholar] [CrossRef]
- García, M.; Pujol, A.; Ruzo, A.; Riu, E.; Ruberte, J.; Arbós, A.; Serafín, A.; Albella, B.; Felíu, J.E.; Bosch, F. Phosphofructo-1-Kinase Deficiency Leads to a Severe Cardiac and Hematological Disorder in Addition to Skeletal Muscle Glycogenosis. PLoS Genet. 2009, 5, 615. [Google Scholar] [CrossRef] [PubMed]
- Hook, J.; Lemckert, F.; Qin, H.; Schevzov, G.; Gunning, P. Gamma Tropomyosin Gene Products Are Required for Embryonic Development. Mol. Cell. Biol. 2004, 24, 2318–2323. [Google Scholar] [CrossRef]
- Rethinasamy, P.; Muthuchamy, M.; Hewett, T.; Boivin, G.; Wolska, B.M.; Evans, C.; Solaro, R.J.; Wieczorek, D.F. Molecular and Physiological Effects of Alpha-Tropomyosin Ablation in the Mouse. Circ. Res. 1998, 82, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Wei, B.; Lu, Y.; Jin, J.-P. Deficiency of Slow Skeletal Muscle Troponin T Causes Atrophy of Type I Slow Fibres and Decreases Tolerance to Fatigue. J. Physiol. 2014, 592, 1367–1380. [Google Scholar] [CrossRef]
- Wei, B.; Jin, J.-P. TNNT1, TNNT2, and TNNT3: Isoform Genes, Regulation, and Structure–Function Relationships. Gene 2016, 582, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ju, Y.; Li, J.; Xie, C.; Ritchlin, C.T.; Xing, L.; Hilton, M.J.; Schwarz, E.M. Troponin T3 Expression in Skeletal and Smooth Muscle Is Required for Growth and Postnatal Survival: Characterization of Tnnt3tm2a(KOMP)Wtsi Mice. Genesis 2013, 51, 667–675. [Google Scholar] [CrossRef]
- Acakpo-Satchivi, L.J.R.; Edelmann, W.; Sartorius, C.; Lu, B.D.; Wahr, P.A.; Watkins, S.C.; Metzger, J.M.; Leinwand, L.; Kucherlapati, R. Growth and Muscle Defects in Mice Lacking Adult Myosin Heavy Chain Genes. J. Cell Biol. 1997, 139, 1219–1229. [Google Scholar] [CrossRef] [PubMed]
- Woolner, S.; O’Brien, L.L.; Wiese, C.; Bement, W.M. Myosin-10 and Actin Filaments Are Essential for Mitotic Spindle Function. J. Cell Biol. 2008, 182, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Szczesna-Cordary, D.; Craig, R.; Diaz-Perez, Z.; Guzman, G.; Miller, T.; Potter, J.D. Fast Skeletal Muscle Regulatory Light Chain Is Required for Fast and Slow Skeletal Muscle Development. FASEB J. 2007, 21, 2205–2214. [Google Scholar] [CrossRef] [PubMed]
- McNamara, J.W.; Sadayappan, S. Skeletal Myosin Binding Protein-C: An Increasingly Important Regulator of Striated Muscle Physiology. Arch. Biochem. Biophys. 2018, 660, 121–128. [Google Scholar] [CrossRef]
- Markert, C.D.; Meaney, M.P.; Voelker, K.A.; Grange, R.W.; Dalley, H.W.; Cann, J.K.; Ahmed, M.; Bishwokarma, B.; Walker, S.J.; Yu, S.X.; et al. Functional Muscle Analysis of the Tcap Knockout Mouse. Hum. Mol. Genet. 2010, 19, 2268–2283. [Google Scholar] [CrossRef] [PubMed]
- MacArthur, D.G.; Seto, J.T.; Raftery, J.M.; Quinlan, K.G.; Huttley, G.A.; Hook, J.W.; Lemckert, F.A.; Kee, A.J.; Edwards, M.R.; Berman, Y.; et al. Loss of ACTN3 Gene Function Alters Mouse Muscle Metabolism and Shows Evidence of Positive Selection in Humans. Nat. Genet. 2007, 39, 1261–1265. [Google Scholar] [CrossRef] [PubMed]
- Missler, M.; Zhang, W.; Rohlmann, A.; Kattenstroth, G.; Hammer, R.E.; Gottmann, K.; Südhof, T.C. Alpha-Neurexins Couple Ca2+ Channels to Synaptic Vesicle Exocytosis. Nature 2003, 423, 939–948. [Google Scholar] [CrossRef]
- Delfini, M.-C.; Mantilleri, A.; Gaillard, S.; Hao, J.; Reynders, A.; Malapert, P.; Alonso, S.; François, A.; Barrere, C.; Seal, R.; et al. TAFA4, a Chemokine-like Protein, Modulates Injury-Induced Mechanical and Chemical Pain Hypersensitivity in Mice. Cell Rep. 2013, 5, 378–388. [Google Scholar] [CrossRef]
- Rowan, M.J.M.; Christie, J.M. Rapid State-Dependent Alteration in Kv3 Channel Availability Drives Flexible Synaptic Signaling Dependent on Somatic Subthreshold Depolarization. Cell Rep. 2017, 18, 2018–2029. [Google Scholar] [CrossRef]
- Taetzsch, T.; Tenga, M.J.; Valdez, G. Muscle Fibers Secrete FGFBP1 to Slow Degeneration of Neuromuscular Synapses during Aging and Progression of ALS. J. Neurosci. 2017, 37, 70–82. [Google Scholar] [CrossRef]
- Seet, L.-F.; Hong, W. Endofin Recruits Clathrin to Early Endosomes via TOM1. J. Cell. Sci. 2005, 118, 575–587. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Karayannis, T.; Au, E.; Patel, J.C.; Kruglikov, I.; Markx, S.; Delorme, R.; Heron, D.; Salomon, D.; Glessner, J.; Restituito, S.; et al. Cntnap4 Differentially Contributes to GABAergic and Dopaminergic Synaptic Transmission. Nature 2014, 511, 236–240. [Google Scholar] [CrossRef]
- Wang, S.; Hu, Y.; Yang, J.; Smith, C.E.; Nunez, S.M.; Richardson, A.S.; Pal, S.; Samann, A.C.; Hu, J.; Simmer, J.P. Critical Roles for WDR72 in Calcium Transport and Matrix Protein Removal during Enamel Maturation. Mol. Genet. Genom. Med. 2015, 3, 302–319. [Google Scholar] [CrossRef]
- Nishi, M.; Takeshima, H.; Hashimoto, K.; Kano, M.; Hashimoto, K.; Kano, M.; Kuriyama, K.; Komazaki, S.; Shibata, S. Motor Discoordination in Mutant Mice Lacking Junctophilin Type 3. Biochem. Biophys. Res. Commun. 2002, 292, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Kaufman, R.J.; Ginsburg, D. LMAN1 and MCFD2 Form a Cargo Receptor Complex and Interact with Coagulation Factor VIII in the Early Secretory Pathway. J. Biol. Chem. 2005, 280, 25881–25886. [Google Scholar] [CrossRef] [PubMed]
- Vagnozzi, R.; Gatto, G.; Kallander, L.; Hoffman, N.; Mallilankaraman, K.; Ballard, V.; Lawhorn, B.; Stoy, P.; Philp, J.; Graves, A.; et al. Inhibition of the Cardiomyocyte-Specific Kinase TNNI3K Limits Oxidative Stress, Injury, and Adverse Remodeling in the Ischemic Heart. Sci. Transl. Med. 2013, 5, 207ra141. [Google Scholar] [CrossRef] [PubMed]
- Gallicano, G.I.; Kouklis, P.; Bauer, C.; Yin, M.; Vasioukhin, V.; Degenstein, L.; Fuchs, E. Desmoplakin Is Required Early in Development for Assembly of Desmosomes and Cytoskeletal Linkage. J. Cell Biol. 1998, 143, 2009–2022. [Google Scholar] [CrossRef]
- Terry, E.E.; Zhang, X.; Hoffmann, C.; Hughes, L.D.; Lewis, S.A.; Li, J.; Wallace, M.J.; Riley, L.; Douglas, C.M.; Gutierrez-Montreal, M.A.; et al. Transcriptional Profiling Reveals Extraordinary Diversity among Skeletal Muscle Tissues. eLife 2018, 7, e34613. [Google Scholar] [CrossRef]
- Błachnio-Zabielska, A.; Zabielski, P.; Baranowski, M.; Gorski, J. Effects of Streptozotocin-Induced Diabetes and Elevation of Plasma FFA on Ceramide Metabolism in Rat Skeletal Muscle. Horm. Metab. Res. 2010, 42, 1–7. [Google Scholar] [CrossRef]
- Chintala, S.; Tan, J.; Gautam, R.; Rusiniak, M.E.; Guo, X.; Li, W.; Gahl, W.A.; Huizing, M.; Spritz, R.A.; Hutton, S.; et al. The Slc35d3 Gene, Encoding an Orphan Nucleotide Sugar Transporter, Regulates Platelet-Dense Granules. Blood 2007, 109, 1533–1540. [Google Scholar] [CrossRef] [PubMed]
- Kinnear, N.P.; Boittin, F.-X.; Thomas, J.M.; Galione, A.; Evans, A.M. Lysosome-Sarcoplasmic Reticulum Junctions A Trigger Zone for Calcium Signaling by Nicotinic Acid Adenine Dinucleotide Phosphate and Endothelin-1. J. Biol. Chem. 2004, 279, 54319–54326. [Google Scholar] [CrossRef]
- Tomatsu, S.; Orii, K.O.; Vogler, C.; Nakayama, J.; Levy, B.; Grubb, J.H.; Gutierrez, M.A.; Shim, S.; Yamaguchi, S.; Nishioka, T.; et al. Mouse Model on N-Acetylgalactosamine-6-Sulfate Sulfatase Deficiency (Galns-/-) Produced by Targeted Disruption of the Gene Defective in Morquio a Disease. Hum. Mol. Genet. 2003, 12, 3349–3358. [Google Scholar] [CrossRef]
- Yamamoto, S.; Oka, S.; Inoue, M.; Shimuta, M.; Manabe, T.; Takahashi, H.; Miyamoto, M.; Asano, M.; Sakagami, J.; Sudo, K.; et al. Mice Deficient in Nervous System-Specific Carbohydrate Epitope HNK-1 Exhibit Impaired Synaptic Plasticity and Spatial Learning. J. Biol. Chem. 2002, 277, 27227–27231. [Google Scholar] [CrossRef] [PubMed]
- Gokhin, D.S.; Ward, S.R.; Bremner, S.N.; Lieber, R.L. Quantitative Analysis of Neonatal Skeletal Muscle Functional Improvement in the Mouse. J. Exp. Biol. 2008, 211, 837–843. [Google Scholar] [CrossRef]
- Esser, K.; Gunning, P.; Hardeman, E. Nerve-Dependent and -Independent Patterns of MRNA Expression in Regenerating Skeletal Muscle. Dev. Biol. 1993, 159, 173–183. [Google Scholar] [CrossRef] [PubMed]
- Pette, D.; Düsterhöft, S. Altered Gene Expression in Fast-Twitch Muscle Induced by Chronic Low-Frequency Stimulation. Am. J. Physiol. 1992, 262, R333–R338. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, M.; Backer, S.; Saintpierre, B.; Izac, B.; Andrieu, M.; Letourneur, F.; Relaix, F.; Sotiropoulos, A.; Maire, P. Single-Nucleus RNA-Seq and FISH Identify Coordinated Transcriptional Activity in Mammalian Myofibers. Nat. Commun. 2020, 11, 5102. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Symbol | Gene Name | Fold Change (So/Ta) | Localization and Biochemical Properties |
|---|---|---|---|
| Metabolism | |||
| Dio1 | deiodinase, iodothyronine, type I | 69.1 | de-iodinate thyroid hormone interaction; FA oxidation; oxidative phosphorylation uncoupling; causes mitochondrial heat production [15] |
| Pon1 | paraoxonase 1 | 68.83 | protection against oxidation for HDLs and LDLs [10] |
| Tecrl | trans-2,3-enoyl-CoA reductase-like | 55 | FA elongation in polyunsaturated FA biosynthesis [17] |
| Tspo2 | translocator protein 2 | 51.12 | free cholesterol trafficking in erythroid cells [11] |
| Acsm5 | acyl-CoA synthetase medium-chain family member 5 | 30.67 | FA β-oxidation [13] |
| Apoa2 | apolipoprotein A-II | 27.64 | modulator of reverse cholesterol transport [12] |
| Akr1c18 | aldo-keto reductase family 1, member C18 | 24.68 | catalyzes progesterone into 20-α-dihydroprogesterone (20-α-OHP) [21] |
| Adtrp | androgen dependent TFPI regulating protein | 21.66 | hydrolyzes FA esters of hydroxy FAs [22] |
| Cyp4f39 | cytochrome P450, family 4, subfamily f, polypeptide 39 | −269.23 | 1ω and 2ω diesters types and cholesteryl (O-Acyl)-w-hydroxy FAs (OAHFAs) production; FA ω-hydroxylase; acyl-ceramide production [19] |
| Amino Acid Metabolism | |||
| Aox3 | aldehyde oxidase 3 | 30.48 | potentially linked to amino acid and retinol metabolisms; oxidizes aliphatic or aromatic aldehydes into carboxylic acid [23] |
| One-Carbon Metabolism | |||
| Gnmt | glycine N-methyltransferase | 26.87 | Methylation of DNA, RNA, proteins, and lipids via the methionine cycle |
| Miscellaneous | |||
| Aldh1a7 | aldehyde dehydrogenase family 1, subfamily A7 | −20.59 | Oxidoreductase, NAD/NADP acceptor [24] |
| Gene Symbol | Gene Name | Fold Change (So/Ta) | Localization and Biochemical Properties |
|---|---|---|---|
| Glycogenolysis and Glycogenesis | |||
| Ppp1r1c | protein phosphatase 1, regulatory inhibitor subunit 1C | 75.78 | indirect regulation of glycogenesis and glycogenolysis |
| Ppp1r3g | protein phosphatase 1, regulatory subunit 3G | 36.99 | PP1 control; indirect regulation of glycogenesis and glycogenolysis [25] |
| Gys2 | glycogen synthase 2 | −90.2 | converts glucose 1-phosphate into glycogen [26] |
| Phkg1 | phosphorylase kinase γ 1 | −8.1 | activates glycogen phosphorylase [29] |
| Phka1 | phosphorylase kinase α 1 | −8 | activates glycogen phosphorylase [29] |
| Phkb | phosphorylase kinase β | −7.5 | activates glycogen phosphorylase [29] |
| Pgm2 | phosphoglucomutase 2 | −6.7 | converts of glucose 6-phosphate and glucose 1-phosphate |
| Pygm | muscle glycogen phosphorylase | −5.8 | converts glycogen into glucose 1-phosphate |
| Agl | amylo-1,6-glucosidase, 4-α-glucanotransferase | −4.4 | glucosidase activity that converts α-1,6-linked branches into α-1,4-linked branches |
| Epm2a | epilepsy, progressive myoclonic epilepsy, type 2 gene α | −3.5 | facilitates the de-phosphorylation of glycogen to promote branching [27] |
| Pgm2l1 | phosphoglucomutase 2-like 1 | −3 | conversion of glucose 6-phosphate and glucose 1-phosphate |
| Glycolysis and Gluconeogenesis | |||
| Pck1 | phosphoenolpyruvate carboxykinase 1, cytosolic | 9.3 | converts oxaloacetate to phosphoenolpyruvate |
| Eno2 | enolase 2, γ neuronal | 3 | converts 3-PG into pyruvate |
| Pfkfb3 | 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 3 | −9.3 | converts fructose-6-phosphate to F2,6-BP; indirectly regulates glycolytic flux [30] |
| Eno3 | enolase 3, β muscle | −5.4 | converts 3-PG into pyruvate in glycolysis |
| Pgk1 | phosphoglycerate kinase 1 | −5.1 | converts 1,2-bisphosphoglycerate and 3-phosphoglycerate |
| Tpi1 | triose-phosphate isomerase 1 | −4.5 | converts dihydroxyacetone phosphate to D-glyceraldehyde 3-phosphate |
| Fbp2 | fructose bisphosphatase 2 | −4.2 | converts F1,6-BP to fructose 6-phosphate |
| Pfkfb4 | 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 4 | −4.1 | converts F2,6-BP to fructose-6-phosphate; indirectly regulates glycolytic flux |
| Pkm | pyruvate kinase, muscle | −3.9 | converts PEP to pyruvate |
| Aldoc | aldolase C, fructose-bisphosphate | −3.9 | converts F1,6-BP to dihydroxyacetone phosphate |
| Aldoa | aldolase A, fructose-bisphosphate | −3.8 | converts F1,6-BP to dihydroxyacetone phosphate; present in developing embryo and adult muscle |
| Pfkfb1 | 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 1 | −3.1 | regulates glycolytic flux |
| Pfkm | phosphofructokinase, muscle | −3 | regulates glycolytic flux; converts fructose 6-phosphate to F1,6-BP [31] |
| Gene Symbol | Gene Name | Fold Change (So/Ta) | Localization and Biochemical Properties |
|---|---|---|---|
| Neuromuscular Junction | |||
| Grp | gastrin releasing peptide | 149.61 | modulates autonomic system; regulates male sexual function; conveys itch sensation |
| Tom1 | target of myb1 trafficking protein | 84.21 | recruits clathrin to endosomal structures [47] |
| Jph3 | junctophilin 3 | 66.32 | formation of junctional membrane complexes; motor coordination [50] |
| Cntnap4 | contactin associated protein-like 4 | 58.74 | cell adhesion molecule and receptor [48] |
| Slc30a3 (Znt3) | solute carrier family 30 (zinc transporter), member 3 | 34.8 | transport of zinc in synaptic vesicles |
| Wdr72 | WD repeat domain 72 | 33.63 | endocytosis, protein reabsorption, and calcium excretion [49] |
| Lypd1 | Ly6/Plaur domain containing 1 | 26.17 | regulates neuronal nicotinic receptors |
| Asphd1 | aspartate β-hydroxylase domain-containing protein 1 | 25.7 | possibly neurotransmission and synaptic vesicle location/function |
| Nrxn3 | neurexin III | 24.74 | synapse organization; regulating neurotransmitter release [43] |
| Nrsn2 | neurensin 2 | 24.72 | transporting small vesicles to perinuclear region to exit towards organelles |
| Fam19a4 (Tafa4) | TAFA chemokine like family member 4 | −199.82 | modulates neuronal excitability and controls somatic sensation threshold [44] |
| Kcnc4 (Kv3.4) | potassium voltage gated channel, Shaw-related subfamily, member 4 | −33.68 | broadens action potential to prevent somatic depolarization [45] |
| Fgfbp1 | fibroblast growth factor binding protein 1 | −28.02 | slows neuromuscular junctions (NMJs) degeneration [46] |
| Sarcolemma, Sarcoplasm, and Sarcoplasmic Reticulum | |||
| Lman1l | lectin, mannose-binding 1 like | 61.63 | binds to glycoproteins transported from endoplasmic reticulum (ER) to ER-Golgi intermediate [51] |
| Tnni3k | TNNI3 interacting kinase | 27.58 | promotes oxidative stress and myocyte death; part of costamere attached to sarcolemma [52] |
| Phactr3 | phosphatase and actin regulator 3 | 26.09 | binds to cytoplasmic actin and regulates PP1 |
| Dsp | desmoplakin | 20.92 | assembles functional desmosomes; maintains cytoskeletal architecture [53] |
| Pak1 | p21 (RAC1) activated kinase 1 | −7.3 | regulates muscle-specific kinase to maintain NMJs, actin remodeling, and glucose uptakes in skeletal muscle |
| Wwp1 | WW domain containing E3 ubiquitin protein ligase 1 | −3.5 | cell proliferation and apoptosis |
| Thin Filament | |||
| Actin | |||
| Actc1 | actin, α, cardiac muscle 1 | −20.7 | assembles into filaments that are involved in muscle contraction, cell motility, cell signaling, and vesicle movement; associated with fetal skeletal muscle |
| Tropomyosin and Tropomodulin | |||
| Tpm3 | tropomyosin 3, γ | 29.17 | controls the actin filament with tropomodulin [32] |
| Tmod1 | tropomodulin 1 | −3.6 | controls the Ca2+-regulated thin filament end with tropomyosin |
| Tpm1 | tropomyosin 1, α | −3.4 | controls the Ca2+-regulated thin filament end with tropomodulin [33] |
| Troponin Complex | |||
| Tnnc1 | troponin C, cardiac/slow skeletal | 47.7 | binds to calcium and exposes the myosin head binding sites; slow skeletal muscle |
| Tnni1 | troponin I, skeletal, slow 1 | 46.2 | binds to actin and inhibits ATPase activity; slow skeletal muscle |
| Tnnt1 | troponin T1, skeletal, slow | 36.6 | anchors to tropomodulin; slow skeletal muscle [34] |
| Tnni3 | troponin I, cardiac 3 | 14.3 | binds to actin and inhibits ATPase activity; cardiac muscle |
| Tnnt2 | troponin T2, cardiac | 3.3 | anchors to tropomodulin; cardiac, embryonic, and neonatal skeletal muscles [35] |
| Tnnt3 | troponin T3, skeletal, fast | −3 | anchors to the tropomodulin; fast skeletal muscle [36] |
| Thick Filament | |||
| Myosin Heavy Chains | |||
| Myh7 | myosin, heavy polypeptide 7, cardiac muscle, β | 34.71 | isoform of myosin present in slow (type I) skeletal muscle fibers [1] |
| Myh4 | myosin, heavy polypeptide 4, skeletal muscle | −57.73 | isoform of myosin present in adult type IIB skeletal muscle fibers [37] |
| Essential Myosin Light Chains | |||
| Myl6b | myosin, light polypeptide 6B | 54.1 | controls cell adhesion, cell migration, tissue architecture, cargo transport, and endocytosis; promotes p53 protein ubiquitination and degradation |
| Myl3 | myosin, light polypeptide 3 | 27 | involved in force development and fine-scale coordinated muscle contraction |
| Myl4 | myosin, light polypeptide 4 | 3.6 | binds to Ca2+; embryonic skeletal muscle and atrial myocardium [2] |
| Regulatory Myosin Light Chains | |||
| Myl2 | myosin, light polypeptide 2, regulatory, cardiac, slow | 41.7 | stiffens myosin neck and aids in myosin head movement |
| Myl10 | myosin, light chain 10, regulatory | 4 | stiffens myosin neck and aids in myosin head movement [38] |
| Mylpf | myosin light chain, phosphorylatable, fast skeletal muscle | −4 | regulates myofilament activation by phosphorylation [39] |
| Myosin Binding Proteins | |||
| Mybpc2 | myosin binding protein C, fast type | −40.7 | regulates of myofilament contraction [40] |
| Mybph | myosin binding protein H | −42.77 | sarcomere contraction; maturation process of auto-phagosomal membranes; inhibition of non-muscle RLC MYL2A and MYH2A |
| Z-line | |||
| Tcap | titin-cap | 3.5 | binds to titin in an anti-parallel complex and stabilizes Z-line [41] |
| Actn3 | actinin α 3 | −33.3 | anchors actin filaments and supports sarcomere [42] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alto, S.I.; Chang, C.-N.; Brown, K.; Kioussi, C.; Filtz, T.M. Gene Expression Profiling of Skeletal Muscles. Genes 2021, 12, 1718. https://doi.org/10.3390/genes12111718
Alto SI, Chang C-N, Brown K, Kioussi C, Filtz TM. Gene Expression Profiling of Skeletal Muscles. Genes. 2021; 12(11):1718. https://doi.org/10.3390/genes12111718
Chicago/Turabian StyleAlto, Sarah I., Chih-Ning Chang, Kevin Brown, Chrissa Kioussi, and Theresa M. Filtz. 2021. "Gene Expression Profiling of Skeletal Muscles" Genes 12, no. 11: 1718. https://doi.org/10.3390/genes12111718
APA StyleAlto, S. I., Chang, C.-N., Brown, K., Kioussi, C., & Filtz, T. M. (2021). Gene Expression Profiling of Skeletal Muscles. Genes, 12(11), 1718. https://doi.org/10.3390/genes12111718