
Transcriptional Changes in Pearl Millet Leaves under Heat Stress

 , ,
, , 
Abstract
:1. Introduction
2. Methods
2.1. Plant Material and Treatment
2.2. RNA Extraction and cDNA Library Construction
2.3. Reads Mapping and Annotations
2.4. Gene Ontology, KEGG Ontology, Enrichment Analysis and Transcription Factor Identification
2.5. K-Mean Analysis
2.6. Determination of APX and SOD Activity
3. Results
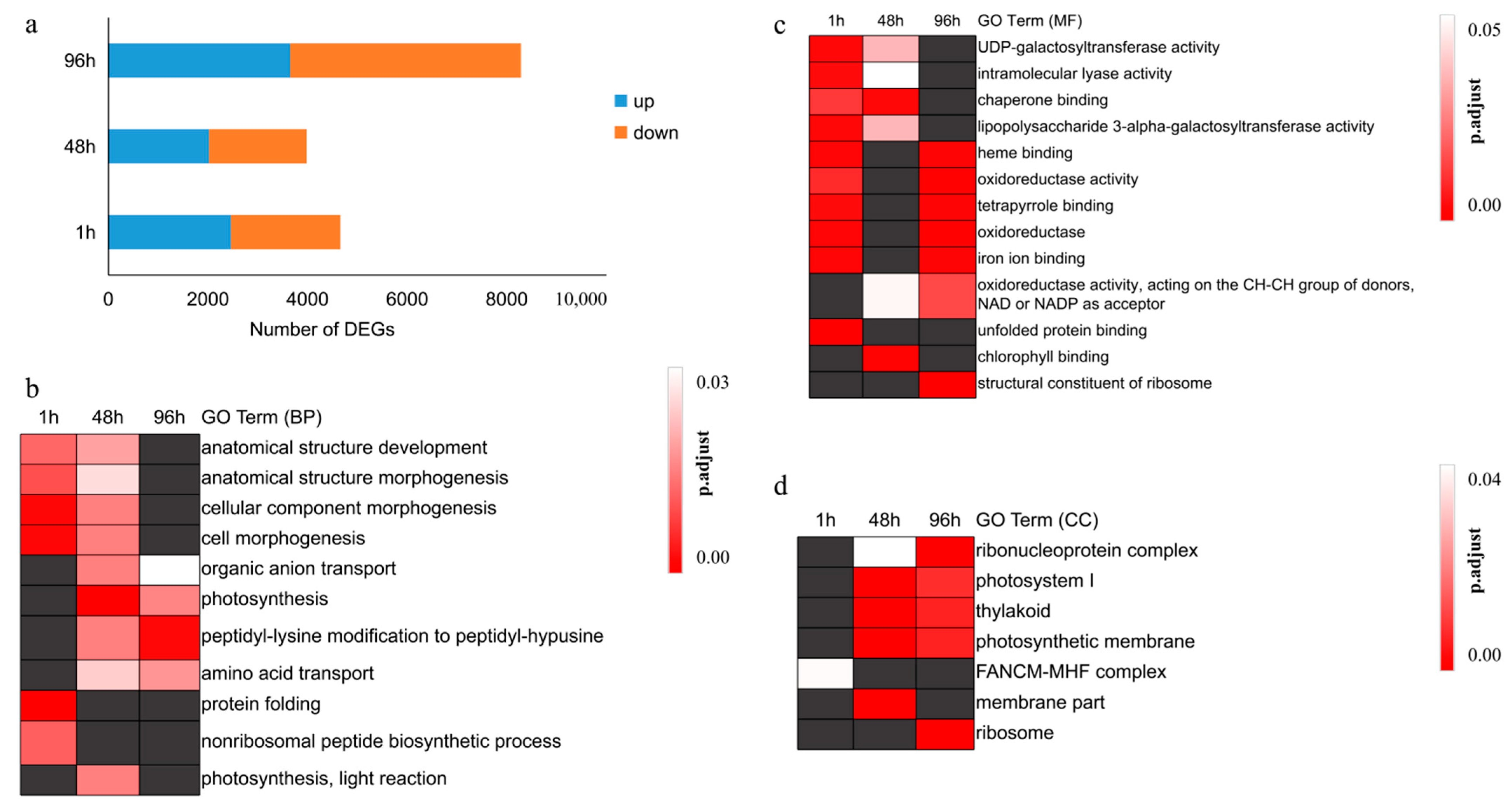
3.1. High Quality of Transcriptome Data Obtained in Pearl Millet
3.2. DEGs Related to Protein Folding and Photosynthesis Respond to Heat Stress in the Short and Medium Term, Respectively
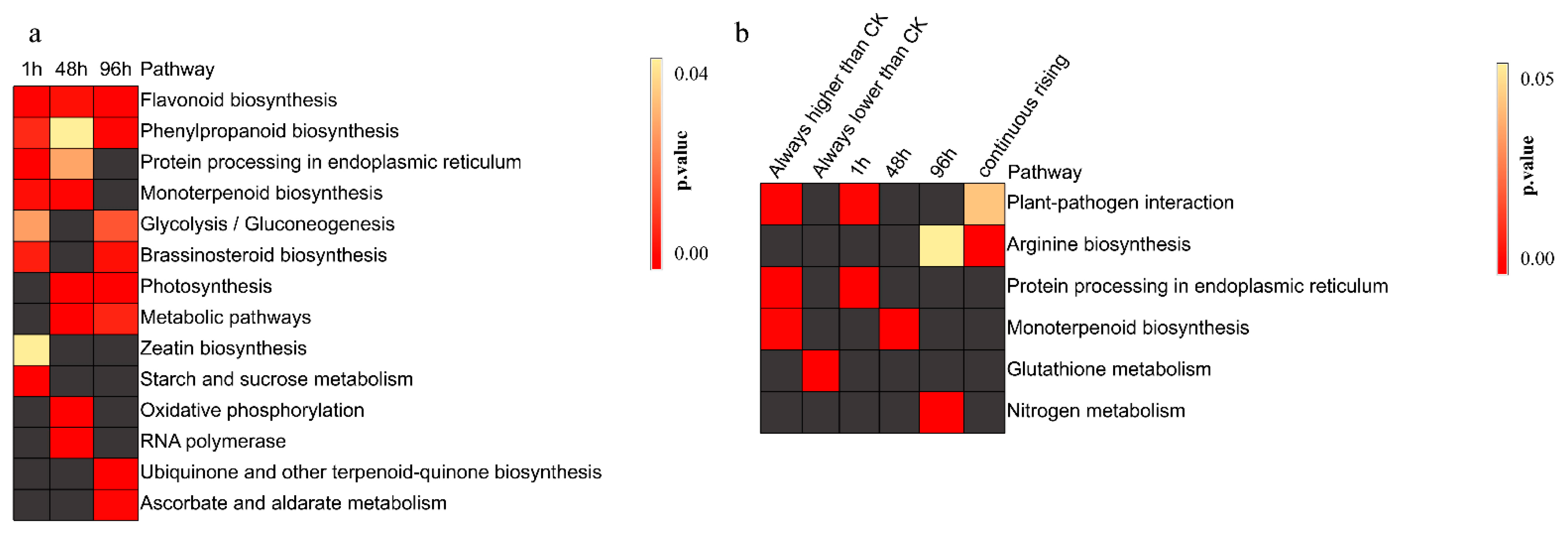
3.3. Heat Stress Affects the Synthesis of Secondary Metabolites
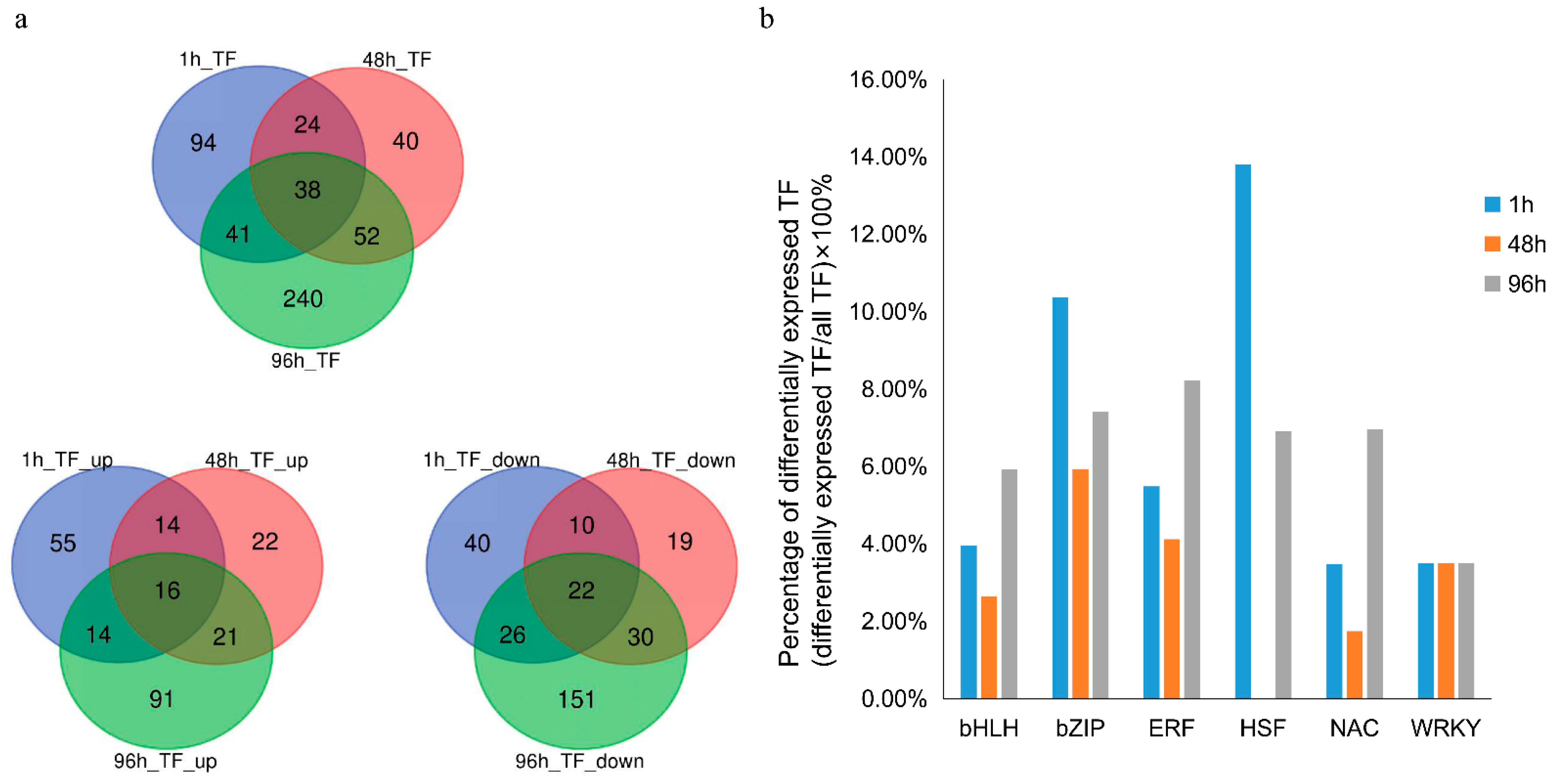
3.4. TF Families Affected by Heat Stress at Each Time Point
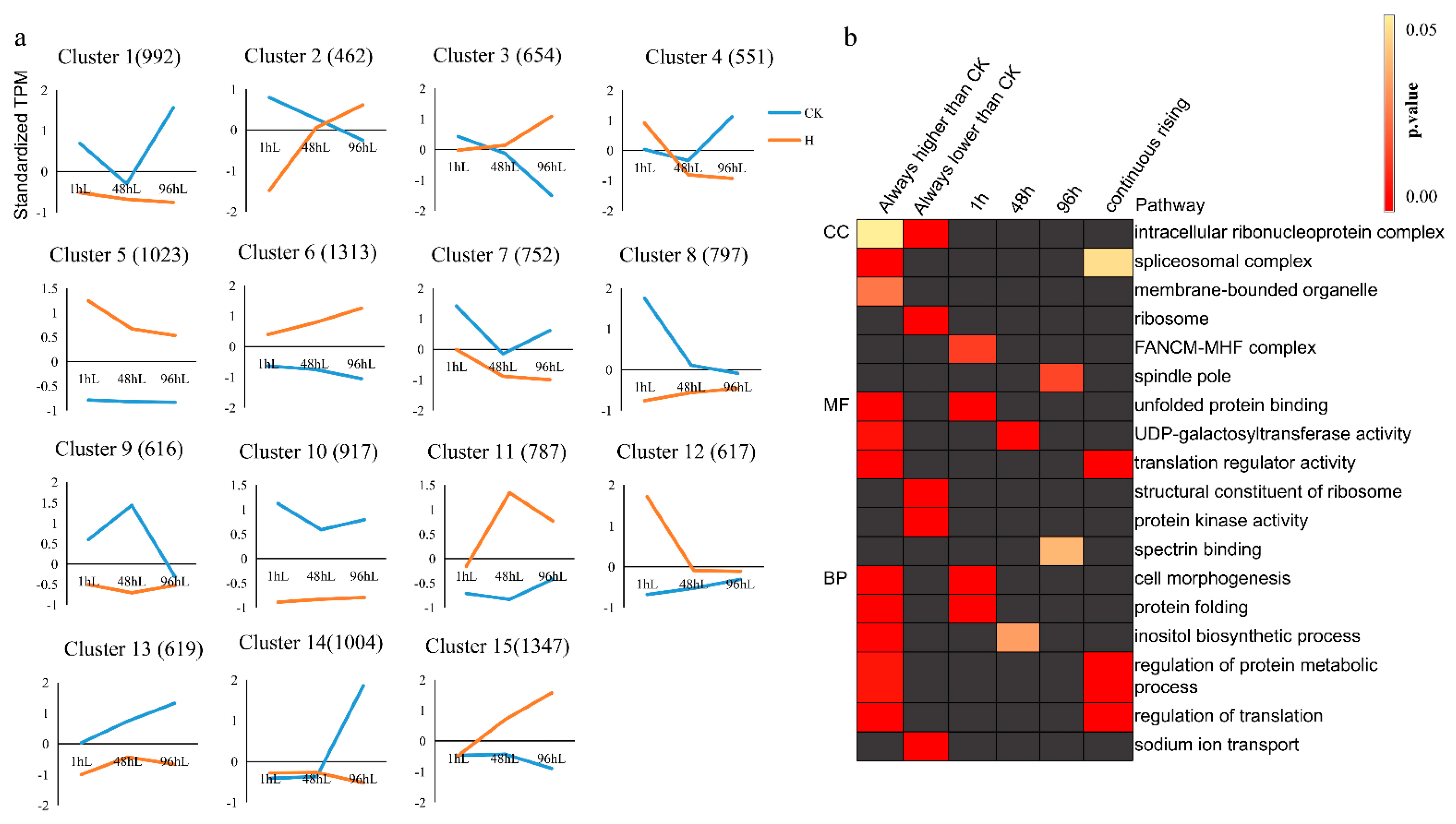
3.5. K-Means Analysis Reveals That Protein Processing in the Endoplasmic Reticulum Is an Important Pathway for Pearl Millet to Respond to Heat Stress
4. Discussion
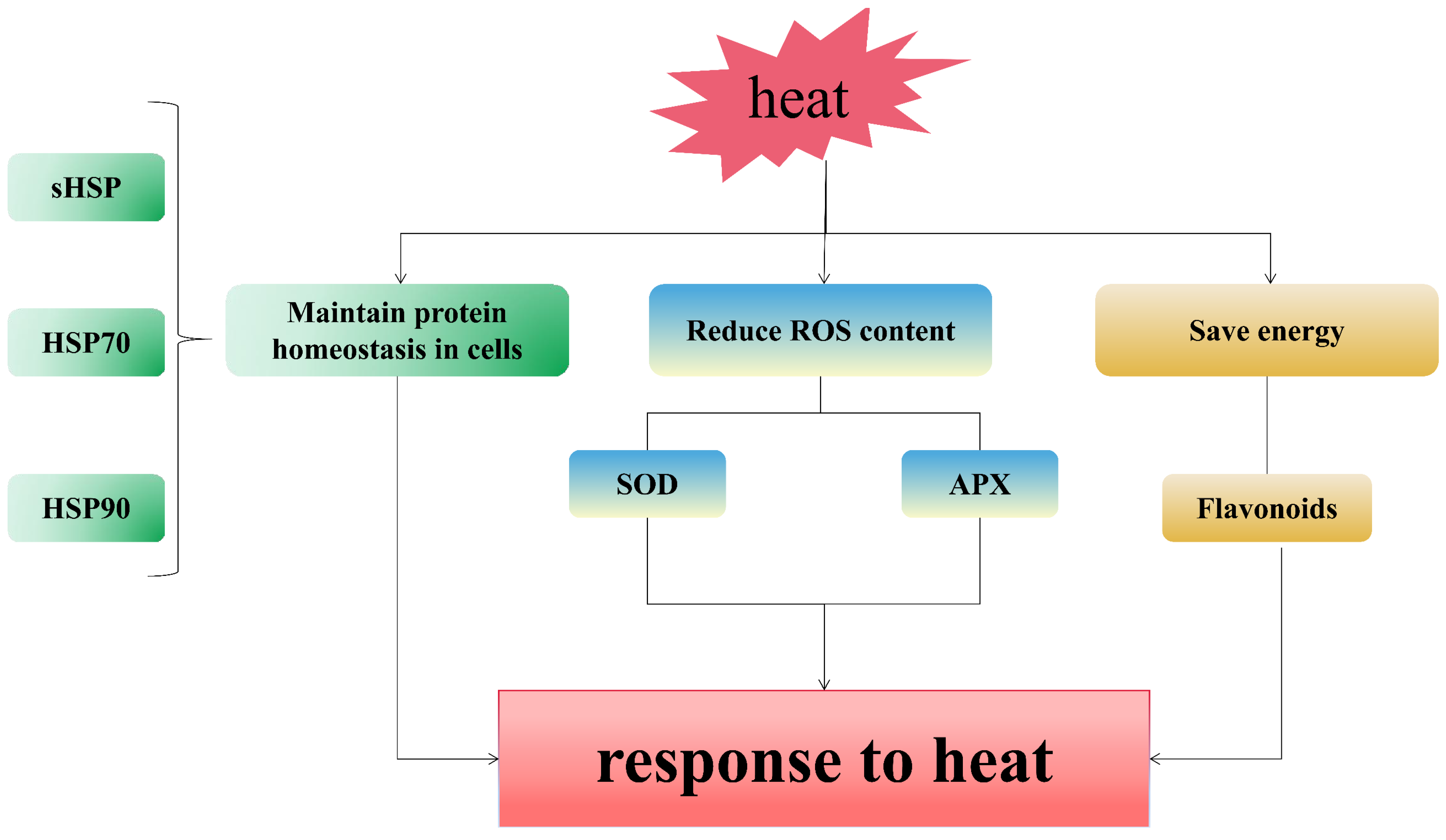
4.1. Rapid Removal of Misfolded Proteins in Cells Is an Important Pathway for Pearl Millet to Respond to Heat Stress
4.2. Maintaining Stable ROS Content Is an Important Link for Pearl Millet to Respond to Short- and Mid-Term Heat Stress
4.3. Pearl Millet Regulates the Synthesis of Secondary Metabolites in Order to Obtain More Energy in the Later Stage of Heat Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| DEG | Differentially expressed gene |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genome |
| HSP | heat shock proteins |
| ROS | reactive oxygen species |
| HSFS | heat stress transcription factors |
| SOD | Superoxide Dismutas |
| CAT | catalase |
| APX | ascorbate peroxidase |
References
- Wahid, A.; Gelani, S.; Ashraf, M.; Fool, M.R. Heat tolerance in plants: An overview. Environ. Exp. Bot. 2007, 61, 199–223. [Google Scholar] [CrossRef]
- Sultan, B.; Defrance, D.; Iizumi, T. Evidence of crop production losses in West Africa due to historical global warming in two crop models. Sci. Rep. 2019, 9, 12834. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Willis, C.G.; Ruhfel, B.; Primack, R.B.; Miller-Rushing, A.J.; Davis, C.C. Phylogenetic patterns of species loss in Thoreau’s woods are driven by climate change. Proc. Natl. Acad. Sci. USA 2008, 105, 17029–17033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.; Ma, K.B.; Lu, Z.G.; Ren, S.X.; Jin, B. Differential physiological, Transcriptomic and metabolomic responses of ara-bidopsis leaves under prolonged warming and heat shock. BMC Plant Biol. 2020, 20, 86. [Google Scholar]
- Wolkovich, E.M.; Cook, B.I.; Allen, J.M.; Crimmins, T.M.; Cleland, E.E. Warming experiments under-predict plant phenological responses to climate change. Nature 2011, 485, 494–497. [Google Scholar] [CrossRef] [PubMed]
- Sun, M.; Lin, C.; Zhang, A.; Wang, X.; Yan, H.; Khan, I.; Wu, B.; Feng, G.; Nie, G.; Zhang, X.; et al. Transcriptome sequencing revealed the molecular mechanism of response of pearl millet root to heat stress. J. Agron. Crop. Sci. 2021, 207, 768–773. [Google Scholar] [CrossRef]
- Yadav, A.K. Genetic dissection of temperature tolerance in pearl millet (Pennisetum glaucum). Indian J. Agric. Sci. 2011, 81, 7–17. [Google Scholar]
- Ashraf, M.; Hafeez, M. Thermotolerance of Pearl Millet and Maize at Early Growth Stages: Growth and Nutrient Relations. Biol. Plant 2004, 48, 81–86. [Google Scholar] [CrossRef]
- Pearson, C. Thermal Adaptation of Pennisetum: Seedling Development. Funct. Plant Biol. 1975, 2, 413–424. [Google Scholar] [CrossRef]
- Khanna-Chopra, R.; Sabarinath, S. Heat-stable chloroplastic Cu/Zn superoxide dismutase in Chenopodium murale. Biochem. Biophys. Res. Commun. 2004, 320, 1187–1192. [Google Scholar] [CrossRef]
- Nitnavare, R.B.; Yeshvekar, R.K.; Sharma, K.K.; Vadez, V.; Reddy, M.K.; Reddy, P.S. Molecular cloning, Characterization and expression analysis of a heat shock protein 10 (Hsp10) from Pennisetum glaucum (L.), a C4 cereal plant from the semi-arid tropics. Mol. Biol. Rep. 2016, 43, 861–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- James, D.; Tarafdar, A.; Biswas, K.; Sathyavathi, T.C.; Padaria, J.C.; Kumar, P.A. Development and characterization of a high temperature stress responsive subtractive cDNA library in Pearl Millet Pennisetum glaucum (L.) R. Br. Indian J. Exp. Biol. 2015, 53, 543–550. [Google Scholar] [PubMed]
- Reddy, P.S.; Thirulogachandar, V.; Vaishnavi, C.S.; Aakrati, A.; Sopory, S.K.; Reddy, M.K. Molecular characterization and expression of a gene encoding cytosolic Hsp90 from Pennisetum glaucum and its role in abiotic stress adaptation. Gene 2011, 474, 29–38. [Google Scholar] [CrossRef] [PubMed]
- Reddy, C.S.; Kim, K.M.; James, D.; Varakumar, P.; Reddy, M.K. PgPAP18, A heat-inducible novel purple acid phosphatase 18-like gene (PgPAP18-like) from Pennisetum glaucum, may play a crucial role in environmental stress adaptation. Acta Physiol. Plant. 2017, 39, 54. [Google Scholar] [CrossRef]
- Bolger, A.M.; Marc, L.; Bjoern, U. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andrews, S. Babraham Bioinformatics-FastQC A Quality Control tool for High Throughput Sequence Data. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 21 July 2021).
- Sun, M.; Huang, D.; Zhang, A.; Khan, I.; Yan, H.; Wang, X.; Zhang, X.; Zhang, J.; Huang, L. Transcriptome analysis of heat stress and drought stress in pearl millet based on Pacbio full-length transcriptome sequencing. BMC Plant Biol. 2020, 20, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bray, N.L.; Pimentel, H.; Melsted, P.; Pachter, L. Erratum: Near-optimal probabilistic RNA-seq quantification. Nat. Biotechnol. 2016, 34, 888. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soneson, C.; Love, M.I.; Robinson, M.D. Differential analyses for rna-seq: Transcript-level estimates improve gene-level inferences. F1000Research 2015, 4, 1521. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K.; Oshlack, A. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef] [Green Version]
- R Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2009; Available online: http://www.R-project.org (accessed on 21 July 2021).
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39, W316–W322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Jiao, C.; Sun, H.; Rosli, H.; Pombo, M.A.; Zhang, P.; Banf, M.; Dai, X.; Martin, G.; Giovannoni, J.J.; et al. iTAK: A Program for Genome-wide Prediction and Classification of Plant Transcription Factors, Transcriptional Regulators, and Protein Kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar] [CrossRef] [Green Version]
- Durak, I.; Yurtarslanl, Z.; Canbolat, O.; Akyol, O. A methodological approach to superoxide dismutase (SOD) activity assay based on inhibition of nitroblue tetrazolium (NBT) reduction. Clin. Chim. Acta 1993, 214, 103–104. [Google Scholar] [CrossRef]
- Cakmak, I.; Marschner, H. Magnesium Deficiency and High Light Intensity Enhance Activities of Superoxide Dismutase, Ascorbate Peroxidase, and Glutathione Reductase in Bean Leaves. Plant Physiol. 1992, 98, 1222–1227. [Google Scholar] [CrossRef] [Green Version]
- Shivhare, R.; Asif, M.H.; Lata, C. Comparative transcriptome analysis reveals the genes and pathways involved in terminal drought tolerance in pearl millet. Plant Mol. Biol. 2020, 103, 639–652. [Google Scholar] [CrossRef] [PubMed]
- Gill, S.S.; Anjum, N.A.; Hasanuzzaman, M.; Gill, R.; Trivedi, D.K.; Ahmad, I.; Pereira, E.; Tuteja, N. Glutathione and glutathione reductase: A boon in disguise for plant abiotic stress defense operations. Plant Physiol. Biochem. 2013, 70, 204–212. [Google Scholar] [CrossRef]
- Kim, J.H.; Lim, S.D.; Jang, C.S. Oryza sativa heat-induced RING finger protein 1 (OsHIRP1) positively regulates plant response to heat stress. Plant Mol. Biol. 2019, 99, 545–559. [Google Scholar] [CrossRef]
- The Long Road to Safe Food. Available online: https://thefinancialexpress.com.bd/views/the-long-road-to-safe-food-1547568954 (accessed on 21 July 2021).
- Karim, I.; Bahmid, N.A. Pearl millet (Pennisetum glaucum) farming for food security: Gross output, net farm income, and B/C ratio. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: Bristol, UK, 2019; Volume 235, p. 012044. [Google Scholar] [CrossRef]
- Thivierge, M.-N.; Chantigny, M.H.; Seguin, P.; Vanasse, A. Sweet pearl millet and sweet sorghum have high nitrogen uptake efficiency under cool and wet climate. Nutr. Cycl. Agroecosystems 2015, 102, 195–208. [Google Scholar] [CrossRef]
- Savchenko, G.E.; Klyuchareva, E.A.; Abramchik, L.M.; Serdyuchenko, E.V. Effect of Periodic Heat Shock on the Inner Mem-brane System of Etioplasts. Russ. J. Plant Physiol. 2002, 49, 349–359. [Google Scholar] [CrossRef]
- Cooper, P.; Ho, T.-H.D. Heat Shock Proteins in Maize. Plant Physiol. 1983, 71, 215–222. [Google Scholar] [CrossRef] [Green Version]
- Hayer-Hartl, M.; Bracher, A.; Hartl, F.U. The GroEL-GroES Chaperonin Machine: A Nano-Cage for Protein Folding. Trends Biochem. Sci. 2016, 41, 62–76. [Google Scholar] [CrossRef] [PubMed]
- Oh, S.E.; Yeung, C.; Babaei-Rad, R.; Zhao, R. Cosuppression of the chloroplast localized molecular chaperone HSP90.5 impairs plant development and chloroplast biogenesis in Arabidopsis. BMC Res. Notes 2014, 7, 643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohama, N.; Sato, H.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Transcriptional Regulatory Network of Plant Heat Stress Re-sponse. Trends Plant Sci. 2017, 22, 53–65. [Google Scholar] [CrossRef]
- Roldán-Arjona, T.; Ariza, R.R. Repair and tolerance of oxidative DNA damage in plants. Mutat. Res. Fundam. Mol. Mech. Mutagenesis 2008, 681, 169–179. [Google Scholar] [CrossRef] [Green Version]
- Aaron, B.; Ron, M.; Nobuhiro, S. ROS as key players in plant stress signalling. J. Exp. Bot. 2014, 65, 1229–1240. [Google Scholar]
- Gilroy, S.; Białasek, M.; Suzuki, N.; Górecka, M.; Devireddy, A.R.; Karpiński, S.; Mittler, R. ROS, Calcium, and Electric Signals: Key Mediators of Rapid Systemic Signaling in Plants. Plant Physiol. 2016, 171, 1606–1615. [Google Scholar] [CrossRef]
- Huang, L.; Jia, J.; Zhao, X.; Zhang, M.; Huang, X.; Ji, E.; Ni, J.; Jiang, M. The ascorbate peroxidase APX1 is a direct target of a zinc finger transcription factor ZFP36 and a late embryogenesis abundant protein OsLEA5 interacts with ZFP36 to co-regulate OsAPX1 in seed germination in rice. Biochem. Biophys. Res. Commun. 2018, 495, 339. [Google Scholar] [CrossRef]
- Foyer, C.H.; Noctor, G. Oxidant and antioxidant signalling in plants: A re-evaluation of the concept of oxidative stress in a physiological context. Plant Cell Environ. 2005, 28, 1056–1071. [Google Scholar] [CrossRef]
- Kasuga, M.; Liu, Q.; Miura, S.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nat. Biotechnol. 1999, 17, 287–291. [Google Scholar] [CrossRef]
- Winkel-Shirley, B. Flavonoid Biosynthesis. A Colorful Model for Genetics, Biochemistry, Cell Biology, and Biotechnology. Plant Physiol. 2001, 126, 485–493. [Google Scholar]
- Davies, K.M.; Jibran, R.; Zhou, Y.; Albert, N.W.; Schwinn, K.E. The Evolution of Flavonoid Biosynthesis: A Bryophyte Per-spective. Front. Plant Sci. 2020, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Tanaka, Y.; Tsuda, S.; Kusumi, T. Metabolic Engineering to Modify Flower Color. Plant Cell Physiol. 1998, 39, 1119–1126. [Google Scholar] [CrossRef]
- Yin, Y.C.; Zhang, X.D.; Gao, Z.Q.; Hu, T.; Liu, Y. The Research Progress of Chalcone Isomerase (CHI) in Plants. Mol. Biotechnol. 2019, 61, 32–52. [Google Scholar] [CrossRef]
- Yonekura-Sakakibara, K.; Higashi, Y.; Nakabayashi, R. The Origin and Evolution of Plant Flavonoid Metabolism. Front. Plant Sci. 2019, 10, 943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Georgii, E.; Jin, M.; Zhao, J.; Kanawati, B.; Schmitt-Kopplin, P.; Albert, A.; Winkler, J.B.; Schäffner, A.R. Relationships between drought, heat and air humidity responses revealed by transcriptome-metabolome co-analysis. BMC Plant Biol. 2017, 17, 120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Zhao, Y.; Zhu, J.-K. Thriving under Stress: How Plants Balance Growth and the Stress Response. Dev. Cell 2020, 55, 529–543. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Annotation | ID | log2FC (H-1hL/CK-1hL) | log2FC (H-48hL/CK-48hL) | log2FC (H-96hL/CK-96hL) |
|---|---|---|---|---|
| ERF | i1_HQ_LWC_c16728/f8p0/1595 | 1.03 | #N/A | #N/A |
| i1_HQ_LWC_c2213/f2p0/1493 | −1.38 | #N/A | −2.55 | |
| i1_HQ_LWC_c2324/f2p2/1682 | #N/A | #N/A | −1.14 | |
| i1_HQ_LWC_c8407/f5p2/1786 | #N/A | #N/A | 1.01 | |
| i1_LQ_LWC_c11830/f1p1/1488 | #N/A | #N/A | −2.25 | |
| i1_LQ_LWC_c14109/f1p0/1383 | −1.23 | #N/A | −1.5 | |
| i1_LQ_LWC_c20904/f1p47/1586 | #N/A | #N/A | 1.26 | |
| i1_LQ_LWC_c21967/f1p3/1411 | #N/A | #N/A | −1.83 | |
| i1_LQ_LWC_c26384/f1p0/1341 | #N/A | #N/A | −1.96 | |
| i1_LQ_LWC_c36256/f1p0/1962 | 1.08 | #N/A | #N/A | |
| i1_LQ_LWC_c37745/f1p24/1102 | 1.31 | 1.43 | #N/A | |
| i1_LQ_LWC_c8826/f1p2/1547 | #N/A | #N/A | −3.09 | |
| i2_HQ_LWC_c37391/f2p0/2132 | #N/A | #N/A | 2.08 | |
| i2_LQ_LWC_c105823/f1p0/2590 | 1.3 | 1.26 | #N/A | |
| i2_LQ_LWC_c10690/f1p3/2523 | #N/A | #N/A | −1.44 | |
| i2_LQ_LWC_c12095/f1p2/2333 | #N/A | −1.81 | −1.63 | |
| i2_LQ_LWC_c27703/f1p5/2133 | #N/A | −3.16 | −5.43 | |
| i2_LQ_LWC_c37167/f1p2/3040 | #N/A | #N/A | 1.76 | |
| i2_LQ_LWC_c48555/f1p0/2677 | #N/A | #N/A | −1.94 | |
| i2_LQ_LWC_c52587/f1p0/2111 | #N/A | 1.03 | 1.73 | |
| i3_LQ_LWC_c20156/f1p0/3152 | #N/A | #N/A | −1.5 | |
| i3_LQ_LWC_c44933/f1p0/3022 | #N/A | #N/A | 1.07 | |
| bHLH | i0_LQ_LWC_c744/f1p0/869 | #N/A | #N/A | −2.02 |
| i1_HQ_LWC_c16598/f3p0/1835 | #N/A | #N/A | −1.87 | |
| i1_HQ_LWC_c2277/f2p2/1974 | #N/A | 1.69 | 1.78 | |
| i1_HQ_LWC_c2783/f5p0/1890 | 1.48 | #N/A | #N/A | |
| i1_HQ_LWC_c3080/f2p1/1528 | −1.07 | #N/A | −1.09 | |
| i1_LQ_LWC_c19586/f1p0/1685 | #N/A | #N/A | −1.27 | |
| i1_LQ_LWC_c21713/f1p0/1977 | 1.69 | #N/A | #N/A | |
| i1_LQ_LWC_c26613/f1p0/1381 | −2.61 | #N/A | #N/A | |
| i1_LQ_LWC_c30650/f1p0/1843 | 1.3 | #N/A | #N/A | |
| i1_LQ_LWC_c31312/f1p3/1421 | #N/A | #N/A | 4.17 | |
| i1_LQ_LWC_c31916/f1p5/1737 | #N/A | #N/A | −2.65 | |
| i1_LQ_LWC_c32404/f1p0/1537 | −1.07 | #N/A | −1.77 | |
| i1_LQ_LWC_c33040/f1p0/1932 | 1.8 | #N/A | #N/A | |
| i1_LQ_LWC_c37910/f1p2/1185 | −1.05 | #N/A | −1.33 | |
| i1_LQ_LWC_c3911/f1p3/1651 | #N/A | #N/A | −2.88 | |
| i1_LQ_LWC_c39446/f2p0/1411 | #N/A | −1.49 | −1.03 | |
| i2_HQ_LWC_c34348/f2p3/2704 | #N/A | 2.03 | 2.51 | |
| i2_HQ_LWC_c53137/f4p2/2329 | #N/A | −1.43 | −1.31 | |
| i2_HQ_LWC_c7964/f3p0/2230 | #N/A | #N/A | 1.57 | |
| i2_LQ_LWC_c106219/f1p1/2385 | #N/A | #N/A | −2.2 | |
| i2_LQ_LWC_c133064/f1p0/2039 | #N/A | #N/A | −1.03 | |
| i2_LQ_LWC_c36807/f1p4/2205 | #N/A | 1.53 | #N/A | |
| i2_LQ_LWC_c47945/f1p0/2109 | −3.23 | #N/A | #N/A | |
| i2_LQ_LWC_c55090/f1p0/2091 | #N/A | #N/A | 1.56 | |
| i2_LQ_LWC_c68318/f1p0/2330 | #N/A | #N/A | 1.39 | |
| i2_LQ_LWC_c70175/f1p0/2208 | −4.43 | #N/A | #N/A | |
| i2_LQ_LWC_c8845/f1p0/2370 | #N/A | #N/A | 8.43 | |
| i2_LQ_LWC_c9207/f1p0/2162 | 1.25 | #N/A | 1.07 | |
| i2_LQ_LWC_c9259/f1p0/2884 | 1.33 | 1.78 | 1.47 | |
| bZIP | i0_LQ_LWC_c2149/f1p0/988 | 1.76 | #N/A | #N/A |
| i1_HQ_LWC_c11086/f2p0/1795 | −1.26 | #N/A | −1.57 | |
| i1_HQ_LWC_c16237/f3p0/1669 | 1.25 | #N/A | −1.21 | |
| i1_HQ_LWC_c19844/f2p2/1556 | #N/A | #N/A | −1.91 | |
| i1_HQ_LWC_c2355/f7p4/1991 | #N/A | #N/A | −1.85 | |
| i1_HQ_LWC_c25566/f3p0/1372 | #N/A | 1.49 | #N/A | |
| i1_HQ_LWC_c2716/f3p0/1731 | 1.52 | #N/A | #N/A | |
| i1_HQ_LWC_c29681/f2p0/1990 | 1.6 | #N/A | 1.19 | |
| i1_LQ_LWC_c12688/f1p0/1600 | #N/A | #N/A | 1.15 | |
| i1_LQ_LWC_c16633/f2p0/1915 | #N/A | −1.23 | #N/A | |
| i1_LQ_LWC_c17140/f1p0/1733 | #N/A | #N/A | −1.47 | |
| i1_LQ_LWC_c17491/f1p0/1628 | #N/A | #N/A | 1.33 | |
| i1_LQ_LWC_c18697/f1p0/1584 | #N/A | #N/A | −3.62 | |
| i1_LQ_LWC_c20333/f1p0/1549 | #N/A | 2.46 | #N/A | |
| i1_LQ_LWC_c21844/f1p0/1626 | #N/A | 20.47 | −21.21 | |
| i1_LQ_LWC_c27131/f1p4/1187 | 1.31 | 2.52 | 1.8 | |
| i1_LQ_LWC_c31593/f1p0/1765 | −1.87 | #N/A | −1.28 | |
| i1_LQ_LWC_c3398/f1p5/1925 | #N/A | #N/A | −1.08 | |
| i1_LQ_LWC_c38399/f1p0/1812 | #N/A | #N/A | −1.24 | |
| i1_LQ_LWC_c6745/f1p1/1640 | #N/A | #N/A | −1.33 | |
| i1_LQ_LWC_c6814/f1p0/1757 | #N/A | 2.1 | #N/A | |
| i2_HQ_LWC_c17257/f2p0/2441 | #N/A | 1.43 | 1.53 | |
| i2_HQ_LWC_c4682/f2p1/2509 | 1.96 | #N/A | #N/A | |
| i2_HQ_LWC_c5745/f2p2/2371 | −5.28 | −4.07 | −4.77 | |
| i2_LQ_LWC_c10506/f1p0/2572 | 1.12 | #N/A | #N/A | |
| i2_LQ_LWC_c106423/f1p0/2422 | 1.4 | 1.15 | #N/A | |
| i2_LQ_LWC_c114370/f1p0/2103 | #N/A | #N/A | −1.26 | |
| i2_LQ_LWC_c13727/f1p3/2077 | 1.28 | #N/A | 1.66 | |
| i2_LQ_LWC_c24408/f1p0/2193 | 1.41 | #N/A | 1.24 | |
| i2_LQ_LWC_c24803/f1p2/2716 | −2.06 | −2.37 | −7.89 | |
| i2_LQ_LWC_c25536/f1p2/2931 | −2.65 | −3.23 | −2.11 | |
| i2_LQ_LWC_c41884/f1p1/2042 | #N/A | #N/A | −1.1 | |
| i2_LQ_LWC_c46944/f1p7/2377 | #N/A | #N/A | −1.7 | |
| i2_LQ_LWC_c49205/f1p1/2747 | #N/A | #N/A | 1.85 | |
| i2_LQ_LWC_c52220/f1p0/2449 | 2.74 | #N/A | #N/A | |
| i2_LQ_LWC_c70730/f1p2/2959 | −4.38 | −6.06 | #N/A | |
| i2_LQ_LWC_c77190/f1p4/2024 | #N/A | #N/A | −2.62 | |
| i2_LQ_LWC_c81747/f2p2/2111 | #N/A | #N/A | 7.92 | |
| i2_LQ_LWC_c82592/f1p1/2865 | #N/A | −1.02 | −1.5 | |
| i3_LQ_LWC_c21418/f1p0/3155 | 1.15 | 1.98 | 2.09 | |
| i3_LQ_LWC_c36882/f1p6/3993 | 1.56 | #N/A | #N/A | |
| i3_LQ_LWC_c40088/f1p9/3066 | −4.82 | −2.41 | −3.43 | |
| i4_HQ_LWC_c2296/f2p3/4925 | −4.23 | −2.52 | −2.52 | |
| i4_LQ_LWC_c22019/f1p0/4881 | −3.24 | −2.56 | −2.21 | |
| i5_LQ_LWC_c10548/f1p12/5049 | 1.57 | #N/A | #N/A | |
| i7_LQ_LWC_c161/f1p0/7959 | #N/A | #N/A | −5.05 | |
| HSF | i1_LQ_LWC_c18736/f1p1/1715 | 2.58 | #N/A | #N/A |
| i1_LQ_LWC_c22025/f1p2/1544 | #N/A | #N/A | −1.93 | |
| i2_HQ_LWC_c2225/f5p1/2358 | #N/A | #N/A | −4.23 | |
| i2_HQ_LWC_c3339/f2p3/2212 | 1.4 | #N/A | 2.09 | |
| i2_HQ_LWC_c63561/f5p1/2436 | #N/A | −1.62 | −3.11 | |
| i2_HQ_LWC_c7535/f4p0/2304 | #N/A | #N/A | −2.3 | |
| i2_LQ_LWC_c4082/f1p0/2693 | 1.01 | #N/A | 1.13 | |
| i2_LQ_LWC_c49668/f1p0/2181 | #N/A | #N/A | −2.36 | |
| i2_LQ_LWC_c86456/f1p1/2142 | #N/A | #N/A | −3.2 | |
| i2_LQ_LWC_c90147/f1p0/2328 | 3.71 | #N/A | #N/A | |
| i3_LQ_LWC_c14336/f1p0/3022 | #N/A | #N/A | −2.76 | |
| NAC | i1_HQ_LWC_c1312/f6p0/1522 | #N/A | #N/A | −2.02 |
| i1_HQ_LWC_c29731/f2p0/1677 | #N/A | #N/A | −1.41 | |
| i1_HQ_LWC_c8531/f2p0/2019 | #N/A | #N/A | −1.44 | |
| i1_LQ_LWC_c11267/f1p0/1925 | −1.15 | #N/A | #N/A | |
| i1_LQ_LWC_c12732/f1p0/1978 | #N/A | #N/A | 2.52 | |
| i1_LQ_LWC_c23303/f1p0/1691 | #N/A | #N/A | −3.57 | |
| i1_LQ_LWC_c24307/f1p5/1514 | #N/A | −1.18 | −1.35 | |
| i1_LQ_LWC_c31004/f1p0/1806 | #N/A | #N/A | 1.11 | |
| i1_LQ_LWC_c32800/f1p2/1730 | −1.03 | #N/A | −1.92 | |
| i1_LQ_LWC_c33678/f1p1/1854 | #N/A | #N/A | −3.12 | |
| i1_LQ_LWC_c36521/f1p0/1791 | 1.27 | 1.39 | #N/A | |
| i1_LQ_LWC_c37681/f1p0/1237 | −1.66 | #N/A | −1.52 | |
| i1_LQ_LWC_c5458/f1p0/1856 | 1.34 | #N/A | #N/A | |
| i1_LQ_LWC_c6109/f1p2/1914 | −2.38 | #N/A | −2.31 | |
| i1_LQ_LWC_c6781/f1p0/1635 | #N/A | −1.65 | #N/A | |
| i1_LQ_LWC_c9680/f1p3/1862 | #N/A | #N/A | −3.58 | |
| i2_HQ_LWC_c100859/f2p3/2422 | 2.79 | #N/A | #N/A | |
| i2_HQ_LWC_c16790/f4p2/2632 | 1.03 | #N/A | #N/A | |
| i2_HQ_LWC_c2298/f8p3/2112 | #N/A | 1.1 | #N/A | |
| i2_HQ_LWC_c78151/f10p2/2488 | #N/A | #N/A | 2.67 | |
| i2_LQ_LWC_c104913/f1p2/2561 | #N/A | #N/A | 2.33 | |
| i2_LQ_LWC_c105082/f1p0/2164 | #N/A | #N/A | −3.66 | |
| i2_LQ_LWC_c105085/f1p1/2429 | #N/A | #N/A | −1.29 | |
| i2_LQ_LWC_c106176/f1p1/2359 | #N/A | #N/A | 1.52 | |
| i2_LQ_LWC_c106220/f1p0/2782 | #N/A | #N/A | −3.88 | |
| i2_LQ_LWC_c115202/f1p2/2027 | −1.28 | −1.06 | #N/A | |
| i2_LQ_LWC_c6544/f1p1/2222 | #N/A | #N/A | −1.42 | |
| i2_LQ_LWC_c90026/f1p1/2514 | #N/A | #N/A | 1.16 | |
| i3_LQ_LWC_c12481/f1p0/3622 | #N/A | #N/A | 2.13 | |
| i3_LQ_LWC_c13300/f1p0/3336 | #N/A | #N/A | −7.52 | |
| i4_LQ_LWC_c14651/f1p0/4901 | #N/A | #N/A | 3.69 | |
| WRKY | i1_HQ_LWC_c24104/f2p8/1933 | #N/A | #N/A | −1.57 |
| i1_LQ_LWC_c10658/f1p1/1781 | #N/A | #N/A | −4.64 | |
| i1_LQ_LWC_c21040/f1p2/1452 | #N/A | #N/A | −3.08 | |
| i1_LQ_LWC_c35088/f1p0/1805 | #N/A | #N/A | 3.06 | |
| i1_LQ_LWC_c36127/f1p0/1881 | #N/A | −1.28 | −1.11 | |
| i1_LQ_LWC_c36450/f1p2/1858 | 1.64 | 1.54 | 1.35 | |
| i2_LQ_LWC_c13758/f1p0/2102 | #N/A | #N/A | −2.59 | |
| i2_LQ_LWC_c3413/f1p0/2222 | #N/A | 21.72 | #N/A | |
| i2_LQ_LWC_c8508/f1p0/2662 | −1.08 | #N/A | −1.58 | |
| i2_LQ_LWC_c85901/f1p0/2168 | 1.62 | #N/A | #N/A | |
| i3_LQ_LWC_c36452/f1p28/3212 | 1.75 | 1.6 | 1.63 | |
| i7_LQ_LWC_c306/f1p0/7354 | #N/A | #N/A | −1.82 |
| Annotation | ID | log2FC (H-1hL/CK-1hL) | log2FC (H-48hL/CK-48hL) | log2FC (H-96hL/CK-96hL) |
|---|---|---|---|---|
| sHSP | i0_HQ_LWC_c184/f2p0/791 | 6.34 | 4.00 | 2.39 |
| i0_LQ_LWC_c967/f1p0/765 | 8.77 | 6.55 | 5.04 | |
| HSP70 | i2_LQ_LWC_c35762/f1p8/2557 | 2.38 | #N/A | #N/A |
| i2_LQ_LWC_c82615/f1p8/2280 | 3.81 | 1.81 | 1.80 | |
| i2_LQ_LWC_c105136/f1p0/2340 | 9.62 | 6.07 | 4.35 | |
| i2_LQ_LWC_c105813/f1p8/2359 | 3.76 | #N/A | 2.42 | |
| i2_LQ_LWC_c106010/f1p8/2392 | 3.82 | 1.79 | 2.17 | |
| i2_LQ_LWC_c102949/f1p3/2116 | 7.95 | 6.25 | #N/A | |
| i2_LQ_LWC_c109050/f1p8/2511 | 3.46 | 1.70 | 1.67 | |
| i2_HQ_LWC_c43630/f6p12/2432 | 7.21 | 5.62 | 5.56 | |
| i2_HQ_LWC_c50321/f2p2/2517 | 1.58 | 1.38 | 1.10 | |
| i2_HQ_LWC_c69799/f2p1/2652 | 2.17 | #N/A | 1.06 | |
| i2_HQ_LWC_c127192/f35p1/2517 | 1.94 | #N/A | #N/A | |
| i1_LQ_LWC_c32521/f1p1/1976 | 1.90 | 1.79 | #N/A | |
| i2_LQ_LWC_c4855/f1p10/3010 | 3.34 | 1.78 | 1.59 | |
| i2_LQ_LWC_c7394/f1p7/2310 | −1.45 | #N/A | −1.66 | |
| i2_LQ_LWC_c3333/f1p2/2529 | 5.10 | 3.47 | 2.62 | |
| i2_LQ_LWC_c22856/f1p14/2305 | 1.11 | 1.43 | 1.04 | |
| i2_LQ_LWC_c11597/f1p0/2377 | 2.60 | #N/A | #N/A | |
| i2_LQ_LWC_c50733/f1p0/2426 | 2.94 | 2.67 | 1.87 | |
| i2_LQ_LWC_c48249/f1p5/2266 | −1.34 | #N/A | −1.42 | |
| i2_LQ_LWC_c53787/f1p5/2383 | 3.47 | 4.62 | #N/A | |
| i2_LQ_LWC_c66032/f1p53/2457 | 1.45 | #N/A | −1.87 | |
| i2_LQ_LWC_c81928/f1p8/2468 | 2.65 | #N/A | 1.88 | |
| i2_LQ_LWC_c82227/f1p0/2839 | −4.52 | #N/A | #N/A | |
| i2_LQ_LWC_c104760/f1p1/2446 | 2.07 | 1.82 | 1.11 | |
| i3_LQ_LWC_c27490/f1p0/3150 | 2.98 | 3.35 | 3.12 | |
| HSP90 | i2_LQ_LWC_c8803/f1p15/2771 | 1.90 | #N/A | −1.85 |
| i2_LQ_LWC_c33469/f1p1/2516 | 2.61 | #N/A | #N/A | |
| i2_LQ_LWC_c90259/f1p14/2485 | 1.84 | #N/A | #N/A | |
| i2_LQ_LWC_c104439/f1p16/2591 | 2.25 | #N/A | #N/A | |
| i2_LQ_LWC_c121080/f1p8/2511 | 10.70 | 10.13 | 9.17 | |
| i2_LQ_LWC_c126787/f141p12/2519 | 1.53 | #N/A | #N/A | |
| i3_LQ_LWC_c19538/f1p2/3645 | 7.78 | 7.27 | 6.06 | |
| i4_LQ_LWC_c19908/f1p0/5045 | 2.36 | #N/A | #N/A | |
| i2_HQ_LWC_c47794/f3p1/2946 | 4.84 | 2.72 | 1.69 | |
| i2_HQ_LWC_c49563/f2p1/2825 | 4.64 | 3.59 | 3.33 | |
| i2_HQ_LWC_c126707/f9p17/2641 | 3.19 | 2.01 | 2.81 | |
| i2_HQ_LWC_c127285/f2p25/2794 | 3.36 | #N/A | #N/A | |
| i1_LQ_LWC_c22321/f1p0/1789 | 5.38 | #N/A | 4.94 | |
| i2_LQ_LWC_c3878/f1p2/2975 | 2.08 | 2.12 | 1.50 | |
| i2_LQ_LWC_c25419/f1p24/2880 | 2.65 | 2.14 | 1.94 | |
| i2_LQ_LWC_c33264/f1p2/2930 | 1.64 | 2.29 | 2.48 | |
| i2_LQ_LWC_c35607/f1p2/2963 | 2.27 | 2.30 | 1.83 | |
| i2_LQ_LWC_c42955/f2p2/2905 | 1.89 | 2.09 | 1.32 | |
| i2_LQ_LWC_c82169/f1p11/2786 | 3.20 | 1.45 | #N/A | |
| i2_LQ_LWC_c106886/f1p1/2786 | 4.84 | 3.57 | 2.97 | |
| i6_LQ_LWC_c1262/f1p0/6420 | 2.94 | #N/A | 2.71 |
| Annotation | ID | log2FC (H-1hL/CK-1hL) | log2FC (H-48hL/CK-48hL) | log2FC (H-96hL/CK-96hL) |
|---|---|---|---|---|
| SOD | i0_LQ_LWC_c2218/f1p0/833 | 1.51 | #N/A | #N/A |
| i0_LQ_LWC_c429/f1p0/846 | 2.56 | 1.81 | 1.20 | |
| APX | i1_HQ_LWC_c40231/f3p0/1461 | #N/A | 1.03 | #N/A |
| i1_LQ_LWC_c18173/f1p0/1818 | 1.47 | #N/A | −2.29 | |
| i1_LQ_LWC_c18498/f1p3/1627 | #N/A | 1.30 | #N/A | |
| i1_LQ_LWC_c38632/f1p0/1092 | 1.65 | 1.41 | #N/A | |
| i2_LQ_LWC_c66599/f1p1/2322 | 1.12 | #N/A | −1.74 | |
| i6_LQ_LWC_c640/f1p0/6667 | 2.10 | #N/A | #N/A |
| Annotation | ID | log2FC (H-1hL/CK-1hL) | log2FC (H-48hL/CK-48hL) | log2FC (H-96hL/CK-96hL) |
|---|---|---|---|---|
| C4H | i1_HQ_LWC_c27582/f8p0/1861 | #N/A | #N/A | −1.54 |
| i1_HQ_LWC_c10429/f5p1/1861 | #N/A | #N/A | −1.56 | |
| i1_LQ_LWC_c7551/f1p5/1917 | #N/A | #N/A | −4.07 | |
| CHS | i1_HQ_LWC_c1494/f3p0/1849 | −3.63 | #N/A | −2.63 |
| i1_LQ_LWC_c3272/f1p0/1605 | #N/A | −3.62 | −7.70 | |
| i1_LQ_LWC_c33680/f1p2/1647 | −4.07 | −2.25 | −3.29 | |
| i2_LQ_LWC_c18273/f1p6/2548 | −4.82 | #N/A | −2.49 | |
| i3_LQ_LWC_c22293/f1p0/3430 | −4.55 | −2.31 | −2.37 | |
| i4_LQ_LWC_c9627/f1p4/4666 | #N/A | −2.81 | −4.93 | |
| CHI | i1_LQ_LWC_c26701/f1p0/1083 | −2.10 | −2.32 | −4.31 |
| i1_LQ_LWC_c42238/f1p0/1048 | #N/A | #N/A | −1.94 | |
| i2_LQ_LWC_c66390/f1p2/2066 | #N/A | #N/A | −1.63 | |
| F3′H | i2_LQ_LWC_c5478/f1p14/2479 | 2.31 | #N/A | 2.41 |
| i3_LQ_LWC_c10106/f1p9/3646 | 1.63 | #N/A | 2.09 | |
| i1_LQ_LWC_c5071/f1p0/1485 | −6.05 | −7.62 | −6.14 | |
| CYP75B1 | i1_HQ_LWC_c27509/f19p0/1855 | −4.80 | −3.29 | −6.41 |
| i1_HQ_LWC_c39849/f7p0/1891 | #N/A | #N/A | −3.19 | |
| i3_HQ_LWC_c29407/f2p0/3044 | −2.91 | −2.82 | −3.15 | |
| i1_LQ_LWC_c32858/f1p1/1996 | −2.45 | −1.75 | −4.30 | |
| i2_LQ_LWC_c47777/f1p4/2093 | −4.79 | −4.36 | −5.65 | |
| HCT | i1_HQ_LWC_c29742/f2p0/1676 | −2.27 | −2.90 | −1.95 |
| i1_HQ_LWC_c18545/f2p0/1773 | #N/A | #N/A | −2.99 | |
| i1_LQ_LWC_c17207/f1p1/1590 | #N/A | #N/A | −5.13 | |
| i1_LQ_LWC_c19818/f1p0/1549 | #N/A | #N/A | 1.45 | |
| i1_LQ_LWC_c29534/f4p0/1653 | #N/A | −1.69 | −1.64 | |
| caffeoyl-CoA O-methyltransferase | i1_LQ_LWC_c38858/f1p0/1286 | #N/A | #N/A | −1.59 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, D.; Sun, M.; Zhang, A.; Chen, J.; Zhang, J.; Lin, C.; Zhang, H.; Lu, X.; Wang, X.; Yan, H.; et al. Transcriptional Changes in Pearl Millet Leaves under Heat Stress. Genes 2021, 12, 1716. https://doi.org/10.3390/genes12111716
Huang D, Sun M, Zhang A, Chen J, Zhang J, Lin C, Zhang H, Lu X, Wang X, Yan H, et al. Transcriptional Changes in Pearl Millet Leaves under Heat Stress. Genes. 2021; 12(11):1716. https://doi.org/10.3390/genes12111716
Chicago/Turabian StyleHuang, Dejun, Min Sun, Ailing Zhang, Jishan Chen, Jian Zhang, Chuang Lin, Huan Zhang, Xiaowen Lu, Xiaoshan Wang, Haidong Yan, and et al. 2021. "Transcriptional Changes in Pearl Millet Leaves under Heat Stress" Genes 12, no. 11: 1716. https://doi.org/10.3390/genes12111716
APA StyleHuang, D., Sun, M., Zhang, A., Chen, J., Zhang, J., Lin, C., Zhang, H., Lu, X., Wang, X., Yan, H., Tang, J., & Huang, L. (2021). Transcriptional Changes in Pearl Millet Leaves under Heat Stress. Genes, 12(11), 1716. https://doi.org/10.3390/genes12111716